World Journal of AIDS
Vol. 2 No. 2 (2012) , Article ID: 19737 , 6 pages DOI:10.4236/wja.2012.22014
Greater Expansion of IFN-(− CD4+ NKT Cells in HIV-1 Compared with HIV-2-Infected Subjects with Preserved CD4+ T Cell Counts
![]()
1MRC Laboratories, Fajara, Banjul, The Gambia; 2School of Medical Sciences, University of Cape Coast, Cape Coast, Ghana; 3London School of Hygiene & Tropical Medicine, London, UK; 4MRC Human Immunology Unit, Weatherall Institute of Molecular Medicine, John Radcliffe Hospital, Oxford, UK.
Email: *sarah.rowland-jones@ndm.ox.ac.uk
Received February 28th, 2012; revised April 5th, 2012; accepted April 15th, 2012
Keywords: NKT cells; HIV-1; HIV-2; IFN-g; CD4 T cells
ABSTRACT
Context: Human Natural Killer T cells are T lymphocytes that express an invariant ab T cells receptors and NK cells receptors. They regulate innate and adaptive immune response but are susceptible to HIV-1 infection. Objective: We compare the frequency and the activity of NKT cells in HIV-1 and HIV-2 infected individuals with CD4+ counts greater than 500/mm3 using flow cytometry after overnight stimulation with phytohemagglutinin (PHA). Results: The frequency of NKT cells was similar between both groups and also to sero-negative control subjects. There were also no significant differences in the proportions of total NKT cells and the CD4+ NKT subset that secreted interferon gamma (IFN-g) after polyclonal stimulation. However, there was a significantly higher frequency of IFN-g− CD4+ NKT cells in HIV-1-infected compared with HIV-2 infected subjects (p = 0.043). Conclusion: These data suggest there is no relationship between the functional activity of NKT cell subsets and the total NKT cell population in HIV infection. The expansion of IFN-g− CD4+ NKT cells in HIV-1 infection may serve as target for viral infection and may eventually result in their depletion during chronic infection.
1. Introduction
Natural Killer T cells represent a small but important subset of the T-lymphocyte lineage [1] with immunoregulatory functions characteristic of both Natural Killer (NK) cells as well as T lymphocytes. They comprises less than 1% of the total lymphocyte population [2] and a significant proportion express conserved invariant ab T cell receptor chains that distinguished them from T lymphocytes: this subset of NKT cells have been named type 1 NKT cells [3]. In humans, the receptors consist of Va24 chains that are preferentially paired with b11 chains [4] and are able to recognise endogenous and exogenous antigens presented by CD1d molecules [5]. About 30% of NKT cells express the CD4 marker but they rarely express CD8 [6]. They also express the effector/memory phenotypes, CD45RO, suggesting that they are chronically activated by auto-antigens [7].
NKT cells respond early in infection, as well as enhancing adaptive immunity by activating cytotoxic T lymphocytes through secretion of Th1/2 cytokines [8], such as IFN-g, TNF-a, IL-4, IL-10 and IL-13 upon activation [9]. These cytokines play a critical role in regulating the immune response: secretion of IL-4 in particular inhibits Th1 responses by inducing a Th2 response [10] and the production of IFN-g enhances Th1 responses resulting in an effective adaptive immune response [11]. Natural Killer T cells respond to infection in the presence of IL-2 and are stimulated to secrete IFN-g which enhances the effective adaptive immune response, making them key players in viral control [12].
Recently, it has been shown that NKT cells are susceptible to HIV-1 infection [7,13,14] and are significantly reduced in peripheral blood of HIV-1 infected individuals with high levels of viremia [15,16]. CD4+ NKT cells appear to be more susceptible to infection than the conventional CD4+ T cells [13] . Thus, they are rapidly depleted during disease progression when the viral load is high, whereas CD4−NKT cells are much less affected by HIV-1 infection.
NKT cells have not been studied in HIV-2 infection, where the majority of subjects progress slowly to disease and maintain low viral load for a decade or more [17]. We postulated NKT cells might be better preserved in HIV-2 infection and play a role in controlling viral replication low. Thus we compared the frequency of NKT cells and their ability to secrete IFN-g in both HIV-1 and HIV-2 infected subjects with high CD4+ T cell count.
2. Methods
The Study subjects: Twenty-five HIV-1 and 25 HIV-2 subjects were recruited from a cohort of HIV infected subjects who were attending Genito-urinary medicine (GUM) clinic at the MRC Laboratories in Fajara, the Gambia [18].
Blood samples: Fifteen millilitres of blood were collected from HIV infected subjects with a CD4-T cell count of >500 cells/ul and also from 30 HIV-negative subjects who donated blood at the Royal Victoria Teaching Hospital Blood Bank, Banjul, The Gambia. Peripheral blood mononuclear cells (PBMCs) were separated from blood by high density Ficoll separation technique. The plasma viral load from the subjects were measured by reverse transcription-PCR using specific long terminal repeat primers, as described elsewhere [19], with the lower cut-off value as 100 copies/ml.
Flow cytometry: Surface and intracellular cytokine staining were carried out using 1 million PBMCs from patients after overnight stimulation with PHA (3 mg/ml). The cells were stained for Vα24, Vβ11 (Beckman Coulters, UK), CD4+ markers and IFN-g after fixation and permeabilisation. About 300,000 event were acquired with four-coloured flow cytometry and the following cell-population: total NKT cells, IFN-g+CD4−, IFNg+CD4+ and IFN-g−CD4+ NKT cells were gated and analysed.
Data analysis: Statistical Analysis was performed on differences between the groups using Mann Whitney U test according to distribution of the data and presented as medians with interquartile ranges. Viral load data were log transformed and analysed by Student T tests with data presented as mean with standard error. The relationships between % total NKT cells and the subsets, % IFN-gNKT, IFN-gCD4+ NKT and FN-g− CD4+ NKT cells, were determined by Spearman’s rank order correlation.
3. Results
The demographic characteristic and lymphocyte subsets of HIV-1, HIV-2-infected subjects and seronegative controls are shown in Table 1. All the infected subjects had CD4+ T cell counts above 500 cells/ul were not showing any clinical sign.
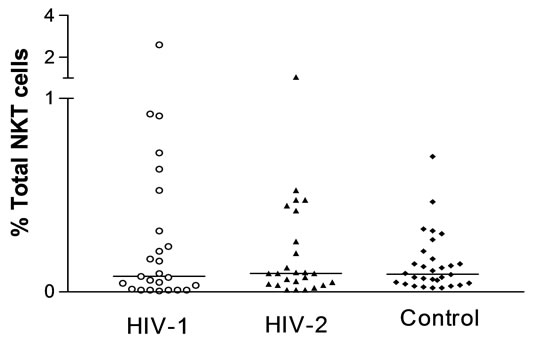
We determined the frequency of NKT cells after stimulating fresh PBMCs with 3 μg/ml of PHA and stained with anti-Vα24 and anti-Vβ11 antibodies (Figure 1). In order to determine the activity of NKT cells in HIV-1, HIV-2 infected subjects and HIV uninfected controls, we measured the frequency of IFN-g+ NKT cells by intracellular cytokine staining. The percentage NKT cells in both HIV-1 and HIV-2 were similar: 0.08 (0.03 - 0.42)% versus 0.09 (0.04 - 0.34)%, p = 0.915 (Figure 2(a)). Similarly, the percentage of NKT cells between HIV-1 subject and HIV uninfected control were 0.08 (0.03 - 0.42)% and 0.09 (0.04 - 19)%, p = 0.939.
Figure 2(b), shows the frequency of NKT cells that secrete IFN-g after stimulating with PHA. The median percentage of NKT cells secreting IFN-g from HIV-1 and HIV-2 infected subjects was similar: 1210 (0.10 - 8.63)% versus 2.63 (0.10 - 10.35)%, p = 0.591 respectively. These were similar to the HIV uninfected control: HIV-1 0.10 (0.10 - 8.63)% versus HIV uninfected controls 0.10 (0.10 - 20.70)%, p = 0.836 and HIV-2, 2.68 (0.10 - 10.35) % versus HIV uninfected control 0.10 (0.10 - 20.70)% p = 0.499.
We also wanted to find out whether all the IFN-g producing NKT cells are CD4+. We noted that some of the CD4+ NKT cells did not produce IFN-g. However, the frequency of CD4+ NKT cells secreting IFN-g was also
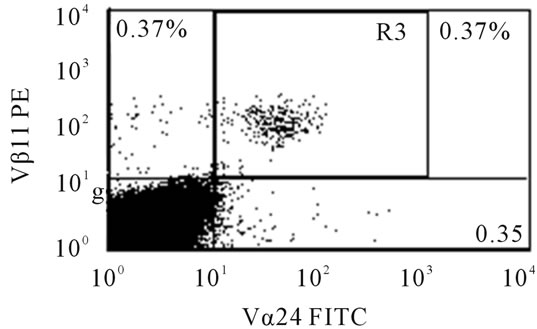
Figure 1. The dot-plot of NKT cells as identified by staining for invariant Vα24 and Vβ11 chains.
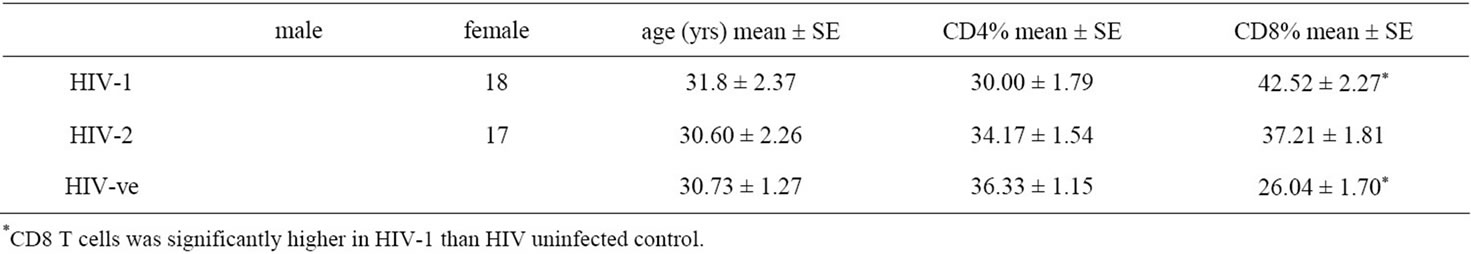
Table 1. Characteristic of HIV infected patients and uninfected controls.
 (a)
(a)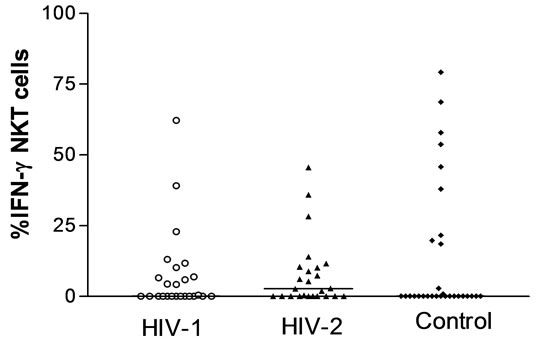 (b)
(b)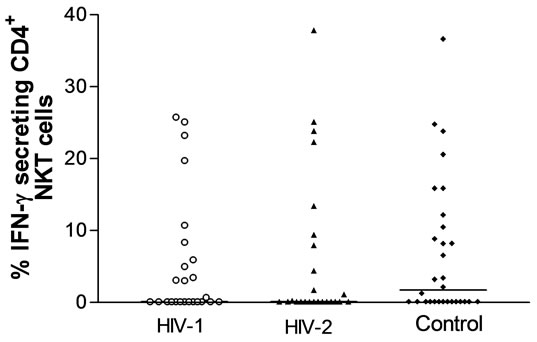 (c)
(c)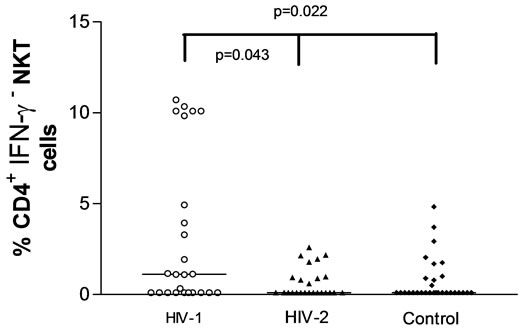 (d)
(d)
Figure 2. The frequency of circulating total NKT cells (a), IFN-g+ CD4− (b), IFN-g+CD4+ (c) and IFN-g− CD4+ NKT (d) cells defined by Vα24Vβ11 expression in HIV-1, HIV-2 infected subjects and HIV uninfected controls. Significant p values are shown.
similar though lower in HIV-1 than HIV-2 infected subjects: HIV-1 0.10 (0.10 - 7.18)% versus HIV-2 0.10 (0.10 - 8.68)% p = 0.852 (Figure 2(c)). They were also similar to HIV uninfected controls: HIV-1 0.10 (0.10 - 7.18)% versus HIV uninfected controls 1.02 (0.10 - 11.33)%, p = 0.627; HIV-2 infected subjects versus HIV uninfected controls, p = 0.533.
On the other hand, the frequency of CD4+ NKT cells, which did not secrete IFN-g were significantly higher in HIV-1 infected subjects 1.12 (0.10 - 7.40)% as compare to HIV-2 infected subjects, 0.10 (0.10 - 0.95)%, p = 0.043 and HIV uninfected controls 0.10 (0.10 - 0.96)%, p = 0.022 (Figure 2(d)). There was also similar percentage IFN-g− CD4+ NKT cells in HIV-2 infected subjects and HIV uninfected control, p = 0.740.
We also evaluated the relationship between percent total NKT cells and their subsets in both HIV-1 and HIV-2 infected subjects to determine any association between them. There was no correlation between total NKT cells and IFN-g producing NKT cells in both HIV-1 infected subjects, r = 0.096, p = 0.646 and HIV-2 infected subjects, r = 0.059, p = 0.783 (Table 2). For total NKT cells and IFN-g− CD4+ NKT subsets, there was no relationship in HIV-1 infected individuals (r = −0.239, p = 0.250) or HIV-2 infected subjects (r = −0.227, p = 0.274). There was also no correlation between total NKT cells and IFN-g− CD4+ NKT subsets in HIV-1, r = 0.030, p = 0.886 and in HIV-2 infected subjects r = 0.079, p = 0.705.
The mean log plasma virus load was significantly higher in HIV-1 (4.2 ± 0.20) than HIV-2 (3.3 ± 0.20) subjects, p = 0.003 (Figure 3).
4. Discussion
The results of the study showed that the frequencies of circulating NKT cells as well as the proportion of NKT cells secreting IFN-g are similar between HIV-1 and HIV-2 infected subjects when the CD4+ count is in the normal range. However, a significantly greater proportion of the CD4+ NKT cell subset failed to secrete IFN-g in the HIV-1-infected group, compared to both HIV-2
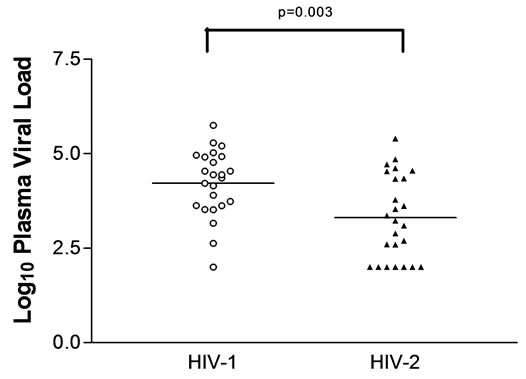
Figure 3. The quantification of plasma viral load in HIV-1 and HIV-2 infected subjects. The mean plasma viral load is represented in horizontal line and significant p value are shown.
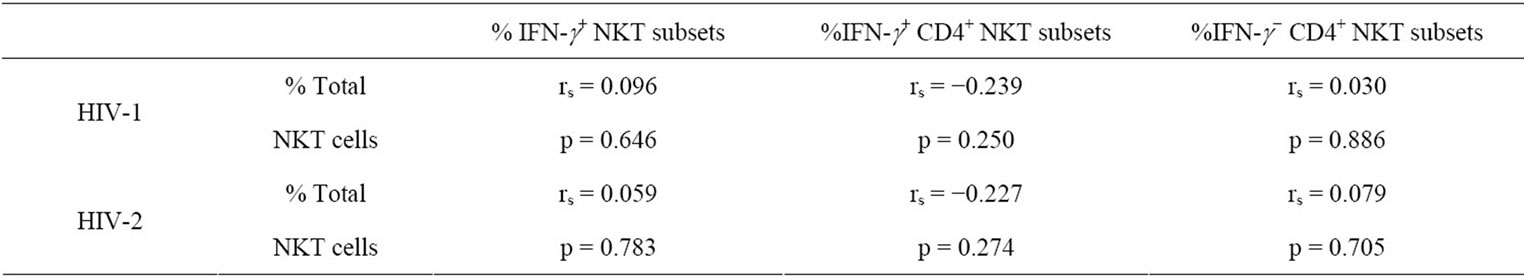
Table 2. The spearman’s rank correlation coefficient between the percent total NKT cells and % IFN-g+ NKT, %IFN-g+CD4+ and %IFN-g− CD4+ subsets.
infected subjects and the controls (Figure 2(d)), suggesting that the functional phenotype of NKT cells differs between HIV-1 and HIV-2 infection.
Early HIV-1 infection is associated with intense immune activation [20], which results in increased T lymphocyte proliferation at the acute stage of infection [21]. Although we were unable to determine whether the infected subjects were in primary or clinically asymptomatic stage of HIV infection, NKT cells might contribute greatly to the enhanced immune action. The dynamics of NKT cell population in acute HIV infection have not yet been determined, but it might be expected that this population would expand to support the adaptive immune response [12]. The secretion of IFN-g by NKT cells play a major role in bridging the gap between innate and adaptive immune response [22,23], and therefore important in controlling viral infection [23]. However, we found no increase in the overall numbers of circulating NKT cells in either HIV-1 or HIV-2-infected subjects in the asymptomatic phase of infection.
We also wanted to determine whether all HIV infection had an impact on the proportion of IFN-g producing NKT cells that also express CD4. The CD4 molecule is the main receptor for both HIV-1 and HIV-2 to enter the cells [24], and it has been proposed that expression of CD4 on NKT cells may lead to their preferential susceptibility to infection [13,24]. It was shown by Motsinger et al., that CD4+ NKT cells are active in the immune response to HIV-1 infection but also serve as target cells for viral infection [7]. These cells differentiate as infection progresses which increases the pool of HIV-susceptible cells [11]. About 40% - 50% of subjects in both HIV-1 and HIV-2 infected groups showed NKT cells that secrete IFN-g and express CD4+ marker: there were no significant differences in the size of this population between HIV-1 and HIV-2 infection, or between HIV infected and uninfected controls.
Our principal finding was that HIV-1-infected subjects had populations of CD4+ NKT cells that could be expanded following PHA stimulation but did not produce IFN-g, which were not seen in HIV-2 infected subjects matched for CD4+ count. It is plausible that increased T cell proliferation in HIV-1 infection due to immune activation may result in expansion of these populations of IFN-g−CD4+ NKT cells. The plasma viral load were also significantly higher in HIV-1 compared with HIV-2 infected subjects (figure 3) which could be driven by immune activation [25,26], that could in turn be associated with expansion of these cell subsets. Immune activation results in stimulation and expansion of NKT cells in vivo and subsequent rapid loss by apoptosis [27]. However, in vitro studies suggest that major NKT cells appear resistant to apoptosis and are capable of expanding significantly without exhibiting any antiviral effect [11]. In contrast, there was no expansion of CD4+ IFN-g− NKT cells in HIV-2 infected subjects compared to HIV uninfected controls suggesting that this population is not perturbed in HIV-2 infection. Further studies are needed to show whether or not HIV-2 infects CD4+ NKT cells and whether the preservation of a normal NKT cell subsets distribution is associated with preserved function in HIV-2 infection.
NKT cells express higher levels of both CCR5 and CD4 receptors than do conventional T cells [1] and this is thought to lead to preferential infection and decreasing level of NKT cells through the course of HIV-1 infection with disease progression [7,13,14]. A decrease in circulating NKT cell numbers has also been noted in several other conditions such as autoimmune diseases [28], bacterial infection [29] as well as other viral diseases [30]. However, HAART and IL-2 treatments of HIV-1 infected subjects led to an expansion in NKT cells [15,31] which could be helpful in enhancing conventional T cells [32]. This study also showed that NKT cells could be expanded in vitro in some subjects with PHA stimulation, but the resulting population was largely CD4+ and failed to secrete IFN-g, making these cells a prime target for HIV-1 infection [1]. It will be important if the expansion of NKT cells could be sustained and restore their function after treatment in HIV infection [15,33]. In HIV-2 infection, the stable population of IFN-g secreting NKT cells and the relatively low frequency (compared to HIV-1) of non IFN-g secreting CD4+ NKT cells in HIV-2 subsets may reduce the likelihood of this population being a target for preferential virus infection. However, further studies needed to assess the actual role of NKT cells in HIV-2 infection. Interestingly, the parent virus of HIV-2, SIV from sooty mangabeys (SIVsm) does not lead to the depletion of NKT cells in infected monkeys, but this may be due to the virtual absence of CD4-expressing NKT cells in sooty mangabeys [34].
5. Conclusion
Although we were not able to determine CCR5 expression on NKT cells from our study subjects, our studies have demonstrated a higher proportion of CD4+ NKT cells with impaired IFN-g production in HIV-1 compared to HIV-2 infected subjects. These may serve as target cells for virus infection and replication leading to their subsequent depletion during chronic infection.
6. Acknowledgements
We do appreciate the contribution of Bakary Sanneh, Mamadi Njie, Abdoulai Jabang at the HIV Category 111 laboratory. Thanks to Sarah Crozier for assisting in statistical analysis of the data. We also thank Tumani Corrah, the Director of MRC Unit in the Gambia for his support. This work was supported by MRC, Head Ofiice, London, United Kingdom.
REFERENCES
- M. Kronenberg, “Toward an Understanding of NKT Cell Biology: Progress and Paradoxes,” Annual Review of Immunology, Vol. 23, 2005, pp. 877-900.
- M. J. Smyth, K. Y. Thia, S. E. Street, E. Cretney, J. A. Trapani, M. Taniguchi, et al., “Differential Tumor Surveillance by Natural Killer (NK) and NKT Cells,” The Journal of Experimental Medicine, Vol. 191, No. 4, 2000, pp. 661-668. doi:10.1084/jem.191.4.661
- S. Vasan and M. Tsuji, “A Double-Edged Sword: The Role of NKT Cells in Malaria and HIV Infection and Immunity,” Seminars in Immunology, Vol. 22, No. 2, pp. 87-96. doi:10.1016/j.smim.2009.11.001
- P. Dellabona, E. Padovan, G. Casorati, M. Brockhaus and A. Lanzavecchia, “An Invariant V Alpha 24-J Alpha Q/V Beta 11 T Cell Receptor Is Expressed in All Individuals by Clonally Expanded CD4-8- T Cells,” The Journal of Experimental Medicine, Vol. 180, No. 3, 1994, pp. 1171- 1176. doi:10.1084/jem.180.3.1171
- M. Brigl and M. B. Brenner, “CD1: Antigen Presentation and T Cell Function,” Annual Review of Immunology, Vol. 22, 2004, pp. 817-890.
- A. Bendelac, N. Killeen, D. R. Littman and R. H. Schwartz, “A Subset of CD4+ Thymocytes Selected by MHC Class I Molecules,” Science, Vol. 263, No. 5154, 1994, pp. 1774-1778. doi:10.1126/science.7907820
- A. Motsinger, D. W. Haas, A. K. Stanic, L. Van Kaer, S. Joyce and D. Unutmaz, “CD1d-Restricted Human Natural Killer T Cells Are Highly Susceptible to Human Immunodeficiency Virus 1 Infection,” The Journal of Experimental Medicine, Vol. 195, No. 7, 2002, pp. 869-879. doi:10.1084/jem.20011712
- I. Kotsianidis, J. D. Silk, E. Spanoudakis, S. Patterson, A. Almeida, R. R. Schmidt, et al., “Regulation of Hematopoiesis in Vitro and in Vivo by Invariant NKT Cells,” Blood, Vol. 107, No. 8, 2006, pp. 3138-3144. doi:10.1182/blood-2005-07-2804
- O. Akbari, P. Stock, E. Meyer, M. Kronenberg, S. Sidobre, T. Nakayama, et al., “Essential Role of NKT Cells Producing IL-4 and IL-13 in the Development of Allergen-Induced Airway Hyperreactivity,” Nature Medicine, Vol. 9, No. 5, 2003, pp. 582-588. doi:10.1038/nm851
- C. H. Kim, B. Johnston and E. C. Butcher, “Trafficking Machinery of NKT Cells: Shared and Differential Chemokine Receptor Expression among V Alpha 24(+) V Beta 11(+) NKT Cell Subsets with Distinct Cytokine-Producing Capacity,” Blood, Vol. 100, No. 1, 2002, pp. 11-16. doi:10.1182/blood-2001-12-0196
- D. Unutmaz, “NKT Cells and HIV Infection,” Microbes and Infection, Vol. 5, No. 11, 2003, pp. 1041-1047. doi:10.1016/S1286-4579(03)00185-0
- C. A. Biron and L. Brossay, “NK Cells and NKT Cells in Innate Defense against Viral Infections,” Current Opinion in Immunology, Vol. 13, No. 4, 2001, pp. 458-464. doi:10.1016/S0952-7915(00)00241-7
- J. K. Sandberg, N. M. Fast, E. H. Palacios, G. Fennelly, J. Dobroszycki, P. Palumbo, et al., “Selective Loss of Innate CD4(+) V Alpha 24 Natural Killer T Cells in Human Immunodeficiency Virus Infection,” Journal of Virology, Vol. 76, No. 15, 2002, pp. 7528-7534. doi:10.1128/JVI.76.15.7528-7534.2002
- H. J. Van Der Vliet, B. M. Von Blomberg, M. D. Hazenberg, N. Nishi, S. A. Otto, B. H. Van Benthem, et al., “Selective Decrease in Circulating V Alpha 24+V Beta 11+ NKT Cells during HIV Type 1 Infection,” The Journal of Immunology, Vol. 168, No. 3, 2002, pp. 1490- 1495.
- M. Moll, J. Snyder-Cappione, G. Spotts, F. M. Hecht, J. K. Sandberg and D. F. Nixon, “Expansion of CD1d-Restricted NKT Cells in Patients with Primary HIV-1 Infection Treated with Interleukin-2,” Blood, Vol. 107, No. 8, 2006, pp. 3081-3083. doi:10.1182/blood-2005-09-3636
- M. W. Mureithi, K. Cohen, R. Moodley, D. Poole, Z. Mncube, A. Kasmar, et al., “Impairment of CD1d-Restricted Natural Killer T Cells in Chronic HIV Type 1 Clade C Infection,” AIDS Research and Human Retroviruses, Vol. 27, No. 5, 2011, pp. 501-509.
- S. L. Rowland-Jones and H. C. Whittle, “Out of Africa: What Can We Learn from HIV-2 about Protective Immunity to HIV-1?” Nature Immunology, Vol. 8, No. 4, 2007, pp. 329-331. doi:10.1038/ni0407-329
- M. F. S. Van Der Loeff, S. Jaffar, A. A. Aveika, S. Sabally, T. Corrah, E. Harding, et al., “Mortality of HIV- 1, HIV-2 and HIV-1/HIV-2 Dually Infected Patients in a Clinic-Based Cohort in the Gambia,” AIDS, Vol. 16, No. 13, 2002, pp. 1775-1783. doi:10.1097/00002030-200209060-00010
- N. Berry, K. Ariyoshi, S. Jaffar, S. Sabally, T. Corrah, R. Tedder, et al., “Low Peripheral Blood Viral HIV-2 RNA in Individuals with High Cd4 Percentage Differentiates HIV-2 from HIV-1 Infection,” Journal of Human Virology, Vol. 1, No. 7, 1998, pp. 457-468.
- L. Papagno, C. A. Spina, A. Marchant, M. Salio, N. Rufer, S. Little, et al., “Immune Activation and CD8+ T-Cell Differentiation towards Senescence in HIV-1 Infection,” PLoS Biology, Vol. 2, No. 2, 2004, p. E20. doi:10.1371/journal.pbio.0020020
- V. Natarajan, R. A. Lempicki, I. Sereti, Y. Badralmaa, J. W. Adelsberger, J. A. Metcalf, et al., “Increased Peripheral Expansion of Naive CD4+ T Cells in Vivo after IL-2 Treatment of Patients with HIV Infection,” Proceedings of the National Academy of Sciences of the United States of America, Vol. 99, No. 16, 2002, pp. 10712-10717. doi:10.1073/pnas.162352399
- D. I. Godfrey, K. J. Hammond, L. D. Poulton, M. J. Smyth and A. G. Baxter, “NKT Cells: Facts, Functions and Fallacies,” Immunology Today, Vol. 21, No. 11, 2000, pp. 573-583. doi:10.1016/S0167-5699(00)01735-7
- J. A. Levy, “The Importance of the Innate Immune System in Controlling HIV Infection and Disease,” Trends in Immunology, Vol. 22, No. 6, 2001, pp. 312-316. doi:10.1016/S1471-4906(01)01925-1
- J. P. Moore, A. Trkola and T. Dragic, “Co-Receptors for HIV-1 Entry,” Current Opinion in Immunology, Vol. 9, No. 4, 1997, pp. 551-562. doi:10.1016/S0952-7915(97)80110-0
- J. V. Giorgi, R. H. Lyles, J. L. Matud, T. E. Yamashita, J. W. Mellors, L. E. Hultin, et al., “Predictive Value of Immunologic and Virologic Markers after Long or Short Duration of HIV-1 Infection,” Journal of Acquired Immune Deficiency Syndromes, Vol. 29, No. 4, 2002, pp. 346-355.
- M. D. Hazenberg, S. A. Otto, B. H. Van Benthem, M. T. Roos, R. A. Coutinho, J. M. Lange, et al., “Persistent Immune Activation in HIV-1 Infection Is Associated with Progression to AIDS,” AIDS, Vol. 17, No. 13, 2003, pp. 1881-1888. doi:10.1097/00002030-200309050-00006
- G. Eberl and H. R. MacDonald, “Rapid Death and Regeneration of NKT Cells in Anti-Cd3epsilonor IL-12- Treated Mice: A Major Role for Bone Marrow in NKT Cell Homeostasis,” Immunity, Vol. 9, No. 3, 1998, pp. 345-353. doi:10.1016/S1074-7613(00)80617-2
- S. Kojo, Y. Adachi, H. Keino, M. Taniguchi and T. Sumida, “Dysfunction of T Cell Receptor AV24AJ18+, BV11+ Double-Negative Regulatory Natural Killer T Cells in Autoimmune Diseases,” Arthritis & Rheumatism, Vol. 44, No. 5, 2001, pp. 1127-1138. doi:10.1002/1529-0131(200105)44:5<1127::AID-ANR194>3.0.CO;2-W
- M. Emoto, Y. Emoto and S. H. Kaufmann, “Interleukin- 4-Producing CD4+ NK1.1+ TCR Alpha/Beta Intermediate Liver Lymphocytes Are Down-Regulated by Listeria monocytogenes,” European Journal of Immunology, Vol. 25, No. 12, 1995, pp. 3321-3325. doi:10.1002/eji.1830251218
- J. A. Hobbs, S. Cho, T. J. Roberts, V. Sriram, J. Zhang, M. Xu, et al., “Selective Loss of Natural Killer T Cells by Apoptosis Following Infection with Lymphocytic Choriomeningitis Virus,” Journal of Virology, Vol. 75, No. 22, 2001, pp. 10746-10754. doi:10.1128/JVI.75.22.10746-10754.2001
- H. J. Van Der Vliet, M. G. Van Vonderen, J. W. Molling, H. J. Bontkes, M. Reijm, P. Reiss, et al., “Cutting Edge: Rapid Recovery of NKT Cells upon Institution of Highly Active Antiretroviral Therapy for HIV-1 Infection,” The Journal of Immunology, Vol. 177, No. 9, 2006, pp. 5775- 5778.
- H. J. Van Der Vliet, H. B. Koon, S. C. Yue, B. Uzunparmak, V. Seery, M. A. Gavin, et al., “Effects of the Administration of High-Dose Interleukin-2 on Immunoregulatory Cell Subsets in Patients with Advanced Melanoma and Renal Cell Cancer,” Clinical Cancer Research, Vol. 13, No. 7, 2007, pp. 2100-2108. doi:10.1158/1078-0432.CCR-06-1662
- D. Unutmaz, “Reviving the Killers, All You Need Is IL2,” Blood, Vol. 107, 2006, pp. 3021-3022.
- N. Rout, J. G. Else, S. Yue, M. Connole, M. A. Exley and A. Kaur, “Paucity of CD4+ Natural Killer T (NKT) Lymphocytes in Sooty Mangabeys Is Associated with Lack of NKT Cell Depletion after SIV Infection,” PLoS One, Vol. 5, No. 3, 2010, p. e9787. doi:10.1371/journal.pone.0009787
NOTES
*Corresponding author.