American Journal of Plant Sciences
Vol.5 No.5(2014), Article ID:43623,9 pages DOI:10.4236/ajps.2014.55074
Alpha-Tomatin against Witches’ Broom Disease
Luiz Humberto Gomes1*, Keila Maria Roncato Duarte2, Felipe Gabriel Andrino1, Gildemberg Amorin Leal Jr.3, Lia Matelli Garcia1, Antonio Figueira4, Simone Possedente de Lira1
1Laboratory of Natural Products, Environmental Chemistry, Department of Exact Sciences, Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, Brazil
2Institute of Animal Science and Pastures, Nova Odessa, Brazil
3Federal University of Alagoas, Maceio, Brazil
4Centro de Energia Nuclear na Agricultura, Universidade de São Paulo, Piracicaba, Brazil
Email: *luhgomes1@usp.br
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


Received 5 November 2013; revised 5 February 2014; accepted 27 February 2014
ABSTRACT
Several plants use secondary metabolites against insects and pathogens attack. Among such metabolites known glycoalkaloids are the most studied. Plants from Solanaceae Family are the most abundant on those alkaloids. Despite alfa-tomatine is a known glycoalkaloid, its specific action against Moniliophthora perniciosa, fungi responsible for witches broom disease in cocoa plantations was obtained in this work. Alfa-tomatine was infusion-extracted from Solanum lycopersicum leaves, and its action against the fungi growth was shown on the bioassays. In these tests, M. perniciosa growth was totally inhibited but other fungus tested, the inhibition was not totally, or did not present effect on growth, showing in this way, the specificity of alfa-tomatine to M. perniciosa. The advantages of work with a natural molecule for the fungi control is the stability of the molecule, its resistance to high temperature and pressure and for its use, the simple method of extraction used. The confirmation of the alfa-tomatine molecule was done by High Performance Liquid Chromatography and Mass Spectrometry in comparison to the Sigma standard.
Keywords:Cocoa; Glycoalkaloids; Solanaceae

1. Introduction
Phytotoxins can be produced by any plant or be induced by stressed conditions and act as a natural herbicide or fungicide. It varies widely in size and in chemical class to which they belong, e.g. peptides, terpenoids, diketopiperazines, macrolides, phenolics. Therefore potentially, it is a unexploited biological reservoir of important substances awaiting discovery [1] [2] . Understanding the mode of action of a host-specific toxin might provide insight into the development of plant pathogen-host plant dynamics [2] . Solanaceae Family are world known as most important crops (tomato and potato) and both are target for a whole range of pathogens. Leaves of Solanum genera produce secondary metabolites known as glycoalcaloids, which can act as protection molecules against fungi and some bacteria. Many virulence genes of plant pathogens are induced only in planta or in the presence of host components [3] . Glicoalcaloids from Solanum species were already used in the control of fungi, showing that further studies with other important fungi diseases can be promising. Among important crops and important diseases in Brazil, Witches’ broom is a most important disease of Theobroma cacao (cacao) caused by the basidiomycete Moniliophthora perniciosa (Stahel) [4] , formerly Crinipellis perniciosa. This fungus occurs in the Amazon Forest infecting cacao, some plants from Solanaceae Family, some woodyliana vine species and others [5] . Control of this fungus is hard and expensive due to the cocoa crop conditions: high humidity, tropical forest environment, high cost fungicides, short-interval applications due to perseverant raining, perennial crop, and traditional methods of farming. The introduction of witches’ broom disease into cocoa producing areas of South America and Caribbean has devastated the chocolate industry [6] . The ultimate method of witches’ broom control [7] will be through genetic resistance, therefore the resistance source “Scavina 6” is not doing very well in field conditions, leading to fungi resistant strains [8] [9] . Moniliophthora perniciosa is a hemibiotrophic pathogen, with contrasting mycelial morphology and behavior during the biotrophic and necrotrophic phases [5] Its biotrophic phase starts with the germination of a basidiospore, apoplast colonization with no specific infection structure [10] . The pathogen generally infects cacao shoots, flower cushions, single flowers, and pods inducing symptoms depending on organ infected and stage of development. Hypertrophic growth of the infected shoots, called ‘brooms’ is the most dramatic symptom, appearing 15 d after infection [7] . External symptoms evolve to tissue necrosis, from where basidiocarps emerge. Control of this fungus is hard and expensive due to the cocoa crops conditions: high humidity, tropical forest environment, high cost fungicides, short interval applications due to perseverant raining, perennial crop, traditional methods of farming [11] Leal et al. 2010. The objectives of this work were to test natural extracts from Solanum lycopersicon and purify such extracts to prove the growth inhibition of M. perniciosa and the specificity of such compound to this fungus.
2. Methods
Biotype assays: the Fungi strains from three different biotypes of Moniliophothora perniciosa: From Theobroma cacao, region: Pará, denominationCP44; from Solanum lycocarpum, region: Minas Gerais, denominationAFL105; and from Liana, region Pará, denomination- 322. were grown in TDA media (wheat bran 3%, dextrose 2%; agar 2%), for 30 days at 26˚C.
Leaves extracts were done using 8 g of fresh leaves from Theobroma cacao (T) and Solanum lycopersicum (S), made as an infusion with boiling water, mixed until cooling and filtered. The extract was used to make the TDA media, designed as TDAT, with Theobroma leaves and TDAS with Solanum leaves. Media were autoclaved at 1 atm for 20 min. before use. Control media was used for the biotypes with TDA formula. Petri dishes, at three replicates were kept in BOD for 10 days at 26˚C.
All vegetable material used were produced under controlled conditions, without use of agrochemicals. Seeds from Solanum lycopersicum were from Santa Clara I-5.300, lote 611625 Agroceres variety and the Theobroma cacao seeds were obtained from fruits of a clonal field at ESALQ Campus. For Solanum lycopersicum, 70 days after seedlings, leaves were cutted after flowering and before fructification period, washed in destilated water, kept in plastic bags and frozen at –80˚C. Theobroma cacao leaves were harvested when plants reach 1 m high. Leaves were also washed and kept the same way.
2.1. Biological Assays
Agar diffusion test was performed using paper discs of 6 mm diameter with vegetal extract, placed into the solid culture media and the target were inoculated by a plug of 8 mm, from older petri dishes, were fungi were previously grown [12] . Inhibition tests with other fungi: Using the same three culture media TDA, TDAT and TDAS, other fungi were tested for its development on such culture media. Fungi were: Fusarium oxysporum f.sp. vasinfectum (cotton pathogen), Colletotrichum truncatum and Cercospora kikuchi, both soy bean pathogens at three replicates each were kept in BOD for 10 days at 26˚C.
Supplementation of TDA media: TDA media were supplemented with carbon sources: 2% of each: dextrose, glycerol, pectin, cellulose, starch and maltose. Tests were also performed with TDAS, TDA + ornamental solanaceae leaves; TDAT.
Solanum leaves extraction methodologies: Extraction protocols were done with S. lycopersicum leaves using Hexan in the Soxlet system; agitation on warm hexan. Samples were artificial dried and oil extracts were used from both fractions.
2.2. Isolation and Identification of Chemical Active Compound
The isolation and identification of active chemical compound, 16 grams of S. lycopersicun leaves were grinded with milli-Q water. The extract was frozen and lyophilized. Dry extract (5.54 g) was solved in milli-Q water (300 mL) and submitted to partition liquid-liquid using dichloromethane and a separation funnel, repeated three times, obtaining two phases: organic and aqueous phase. Aqueous phase was again submitted to partition liquid-liquid with n-butanol, as previously described. Both phases obtained were tested into petri dishes bioassays in agar diffusion test, to assure which phase was responsible for the growth inhibition of M. perniciosa. Butanolic phase showed activity, so it was submitted to a chromatographic separation using a column Sephadex LH-20 with isocratic elution of methanol and the fractions obtained were then analyzed. Thin layer chromatography (TLC) and revealed with Dragendorf for UV.
Similar fractions were mixed and later submitted to new agar diffusion test. Activity fractions were reunid and passed through high performance liquid chromatography attachade to a mass detector (HPLC-MS).
2.3. Chromatography in Pre-Packed Column
Phenomenex Strataâ of different dimensions (2 and 5 g) pre-packed columns were used with silica-gel as stationary phase derivatizated with octadecilsilane groups (C18) and mobile phase using a methanol/water gradient starting with 100% water, ending with 100% methanol. Columns and eluent were chosen according to samples characteristics.
For permeation chromatography a glass column of 20 cm height and 2 cm of internal dimension. Material used as stationary phase was Sephadex LH-20 (Pharmacia Biotechâ). Eluation was done by isocratic method using methanol.
For qualitative analyzes in TLC, chromoleaves of silica-gel 60 over polyester as ultraviolet indicator F254 20 × 20 cm (Aldrich) were used. Chromoleaves were eluated into dichloromethano/methanol 8:2 solvents and revealed undr UV light with Spectroline model ENF-240 C/FE lamp, at 254 nm and 365 nm wavelenght.
2.4. High Performance Liquid Chromatography and Mass Spectrometry
The HPLC equipment was a Alliance model 2695 system, from Waters, a PDA detector (“Photodiode Array Detector”) which allowed us to observ between the wavelength 200 to a 800 nm and a mass detector model Micromass ZQ. For the HPLC-PDA-MS a silica-gel column as stationary phase derivatizated with octadecilsilano groups (C18) was used 250 × 4.6 mm, 5 µm, from Inertsil. The LC-MS/MS equipment was from Agilent 6410 MS (using the Agilent Mass Hunter program) with direct insertion on MRM mode. Cone voltage was 50 V and a positive mode detector. Ions screening were from 100 to 1200 Da. Colision energy used was 60 eV. Mass spectra obtained in spectrophotometer VG-7070 operating at 70 eV. High resolution mass spectra were obtained using nominal resolution of 5000 - 10,000. Internal standard was: perfluorquerosene, 3-nitrobenzil alcohol or polyetilenoglicol. Alfa-tomatine standard used was from Sigma (T2830).
2.5. Statistical Analysis
Statistical analisys of mycelial growth data were done by the R program version 2.3.2.2 (The R Foundation for Statistical Computing, 2009) using a randomized experimental design and a factorial analizes was done of all data from different culture media. ANOVA and Tukey test were performed at 5% of significance.
3. Results
The three biotypes of M. perniciosa were grown at 26˚C, for ten days and after this time, petri dishes were analyzed and in the TDAS medium the three biotypes did not grow, different from TDA medium where the three biotypes showed average growing of 3.6 cm radio from the initial plug at Table 1. In Figure 1 mycelial growth can be visualized during experiment with different culture media.
In a second assay, two biotypes of M. perniciosa were inoculated into TDAS culture media with 8% of Solanum leaves, in comparison to TDA medium. TDAS was autoclavated and the solanum extract was filtered in 0.22 um filter and added after TDA autoclavation. Both biotype did not grow in neither one of the TDAS media (Table 2).
As seing on Tables 1 and 2, statistical differences could be seing on othe different culture media tested, using Tukey’s analysis.
Table 3 shows there is a high specificity of growth inhibition of the Solanum compound to the M.perniciosa fungi, related to other genera, in which the leaves extract did not inhibit mycelial growth. Fusarium oxysporum among other 8 fungi, showed some inhibition, but not total as M. perniciosa.
With such results a chemical study was performed using the S. lycopersicum extract aiming to isolate and identify the compound responsible for the anti-fungi activity. Initially, dichloromethano:water and buthanol: water partitions were used. After separation of two fractions, a biological assay was performed to assure which partition the active compound was present. In the first separation, it was present in the water fraction and in the second, in the buthanol phase.

Table 1. Average mycelial growth (cm) of M. perniciosa in different culture media.
*Different letters differs statistically by Tukey test at 5% of significance.
Table 2. Mycelial Growth average (cm) of M. perniciosa in different culture media.
*Different letters differs statistically by Tukey test at 5 % of significance.
Table 3. Mycelial average growth of fungi in different culture media, with respective host and application.
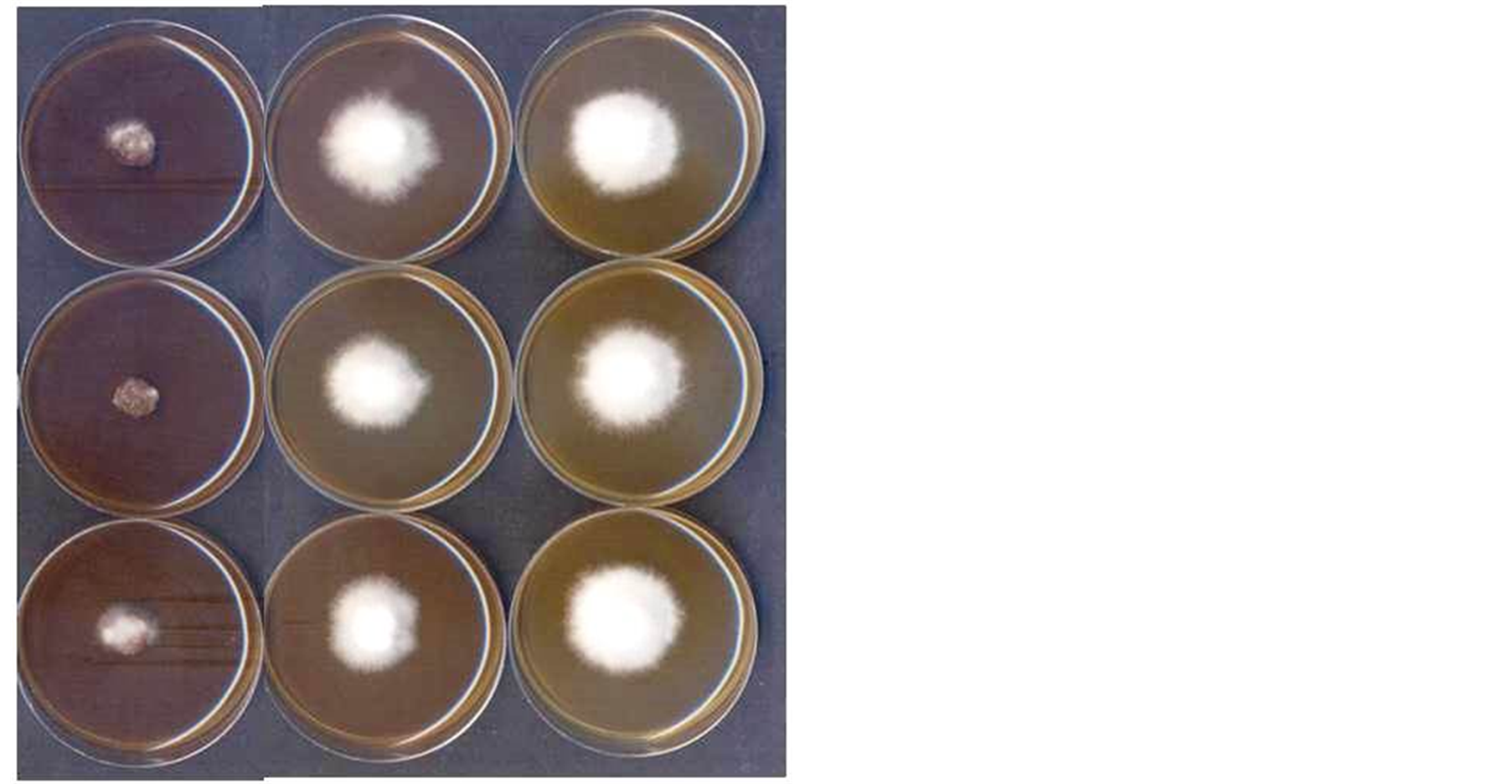
Figure 1. Mycelial growth of 3 biotytes, following the same scheme as the Table 1: from left to right, from above to down: strain CP44 media TDAS; TDAT and TDA; strain AFL105 media TDAS; TDAT and TDA; and strain L322 media TDAS; TDAT and TDA.
Buthanolic fraction was submitted to chromatographic separation in Sephadex-LH-20 gel column, where 14 fractions could be seeing, analyzed by TLC, seen on Figure 2.
Similar chemical fractions were mixed, totalizing eight fractions, again analyzed by TLC (Table 4) and anti-fungi activity was tested in agar diffusion bioassay (Table 4).
Fraction Sex - 5 had more anti-fungi activity and it was selected for the HPLC-MS analyzes.
Figure 3 represents the mass spectro of the crude extract of S. lycopersicum where can be seen the following signals, related to mass/power (m/z): 416, 528, 578, 1034 corresponding to the standard spectro of alfa-tomatina.
Searching for compounds present on S. lycopersicum with anti-fungi activity, alfa-tomatina appears as a major metabolite for this function, present in all parts of the tomato plant. According to [13] the ion 1034.7 reffers to alfa-tomatina mass and the ions 578 and 528 416 are from its fragmentation. Same results can be obtained using alfa-tomatina standard from Sigma (product code—T2830).
Confirming the similarity between ions (m/z) in the S. lycopersicum extracts with the alfa-tomatina standard an anti-fungi activity bioassay using the standard incorporated into TDA medium was performed (Figure 2). Results of this bioassay confirm the efficiency of this glycoalcaloid on the inhibition of the M. perniciosa fungi.
4. Discussion
Looking for a medium in which Moniliophthora perniciosa could present a better growth and basidiocarp development, experiments start to be done with several additions components into TDA media, a known medium for this fungi development [14] . Once strains were collected from Solanaceae family, Theobroma species and Liana species, forming the biotypes we have today of witches’ broom disease pathogens, those leaves extracts were incorporated to TDA media in order to obtain a better growth media, hoping for a media able to induce basidiocarp development [7] [15] . For the group surprise, every time Solanum lycopersicum leaves extracts were present into media, M. perniciosa strains did not grow. Based on this premisse, experiments were conducted in the way to prove the inhibition of Solanum leaves extracts against M. perniciosa fungi.
Results on Table 1 first could be a lack of some nutritional factor or a inhibition of some compound present on tomato leaves. It is known in literature that Solanaceae family has glycoalcaloids such as alfa-tomatine, especially in S. lycopersicum, toxics against a wide range of fungi, described by SANDROCK & Van ETTEN 1997 [16] .
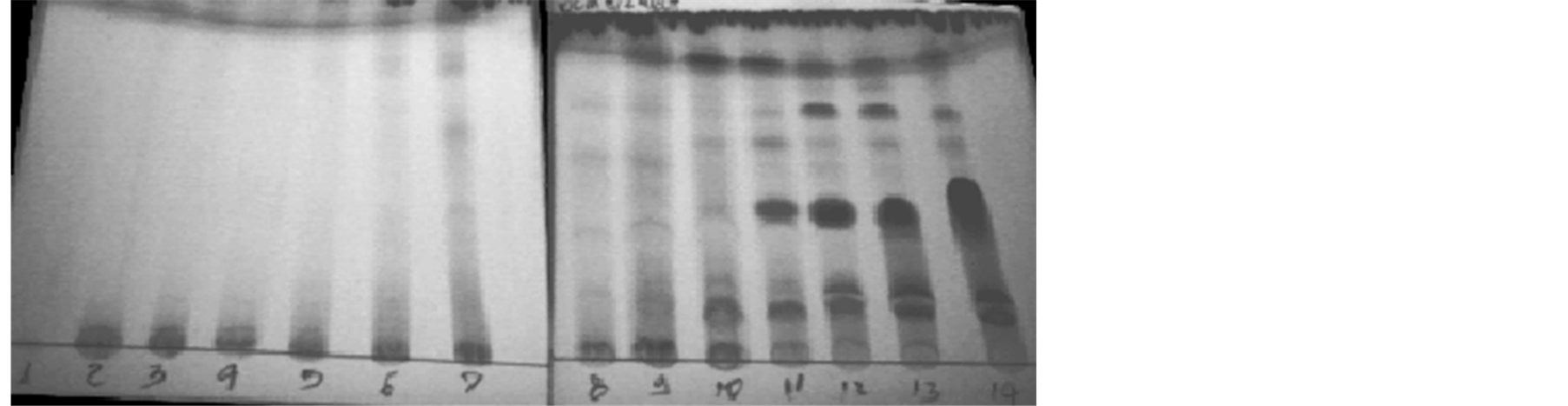
Figure 2. Sephadex gel column of TLC 14 fractions from buthanolic extraction after Dragendorf dye.
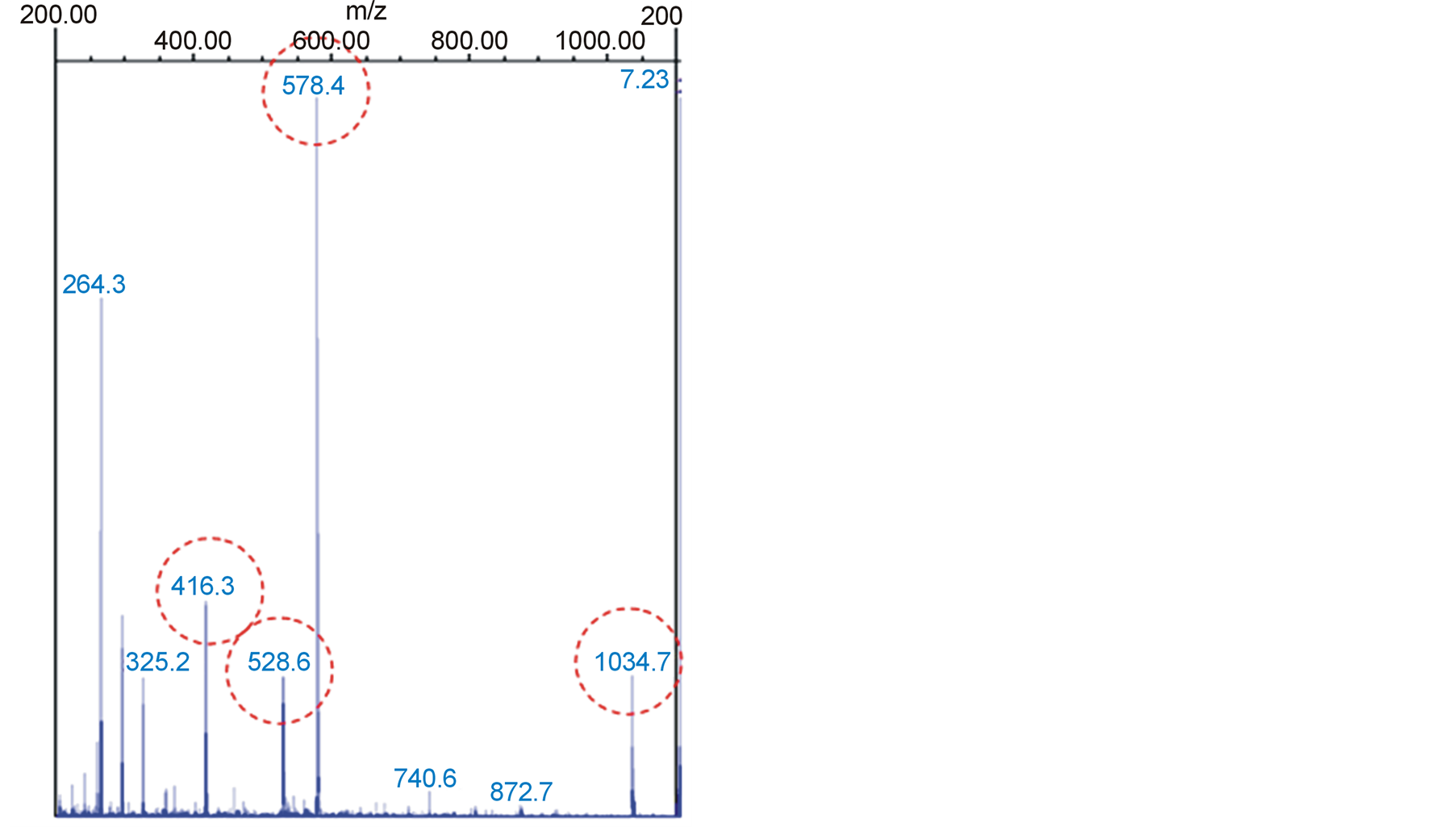
Figure 3. Mass Spectro of crude extract of Solanum lycopersicum. Íons (m/z) circled corresponding to alfatomatina.
Table 4. Chemical similar fractions tested in bioassays for anti-fungi activity.
(+) - positive anti-fungi activity; (-) - negative anti-fungi activity.
To test both hypothesis, TDA medium was again added of S. lycoperiscum leaves (TDAS) and two biotypes inoculated. The no-growth was not for the lack of any nutrient, once in TDA M. perniciosa grew (Table 2), since this is the most suitable media for its growing [7] [11] and due to the presence of a toxic compound in the Solanum leaves.
KUBOW [17] reports the lipid oxidation by heat can lead to polymeric compound potentially toxic. To avoid heating the leave extract during culture media sterilization, leaves extract was filtered in 0.22 um membrane and added to autoclaved TDA medium. Results showed same inhibition using heat or filtration (Table 2), so in this case, no toxic compounds formed by heat are responsible for the fungi growth inhibition. Based upon those results, the alfa-tomatine is a molecule which demands no acid treatment and is stable after heat treatment, characteristics wanted in a natural fungicide, especially for perennial crops.
The anti-fungi activity of the growth inhibitor compound of M.perniciosa present in S. lycopersicum leaves was evaluated against other filamentous fungi, selected from different genera and all pathogens of important crops and of industrial importance (Table 3). In this assay, we verified a specificity of the inhibitor compound related to fungi other than M. perniciosa and F. oxysporum. According to several researchers, the growth inhibition must be due to the presence of glicoalcaloide alfa-tomatine in S. lycopersicum leaves and such inhibition occurs by the linkage of the glycoalcaloid and 3alfa-hidroxy esterol present in the sensible fungi wall, fact already observed by Lairini & Ruiz-Rubio [18] where the mechanism described leads to membrane pores and loss of it is integrity.
In order to confirm alfa tomatine is the molecule responsible for the growth inhibition of M. perniciosa, TDA media was added with alfa tomatine standard Sigma (T-2830) (Figure 4), and we can observe mycelial growth percentage using different doses of standard, in comparison to control, only TDA media. The highest dose used was 30 μg·ml−1 and the inhibition was 78%. According to literature, plants of S. lycopersicum have 10 to 30 mg of a-tomatina per kilo [19] . Considering the maximum concentration of 30 mg per kilo, in our culture media, with 8% of fresh leaves we assume 2 to 2.4 μg per mL. Such dose was enough to inhibit 100% of mycelial growth of M. perniciosa, which is 10 times superior the assay done with standard alfa-tomatina, represented on Figure 2. This result leads us to consider there are interactions between other glycoalcaloids and other chemical compound as in the crude leave extract, potentializing the anti-fungi effect. Similar results were obtained by ABDEL-MOTAAL et al. [20] , proving the interaction of two glycoalcaloides and the increasing of anti-fungi activity.
Cocoa crop is cultivated in humid tropical forests [21] , which can be characterized by a complex ecosystem with saprophytic microorganisms, very important to the litter decomposition. IN this way, a natural molecule to control M. perniciosa is aimed in a way to not interferes on this ecosystem unlike chemical control, reaching all fungi from the environment [22] . The phytopathogen F. oxysporum is emerging as a potential pathogen for mammals due to its resistance to anti-fungi drugs. In this wasy, a natural molecule would be also a troubleshooting. To prevent same resistance to anti-fungi for the control of M. perniciosa the use of alfa-tomatine could mitigate such effects, like those occurred with F. oxysporum, besides the environmental advantages.
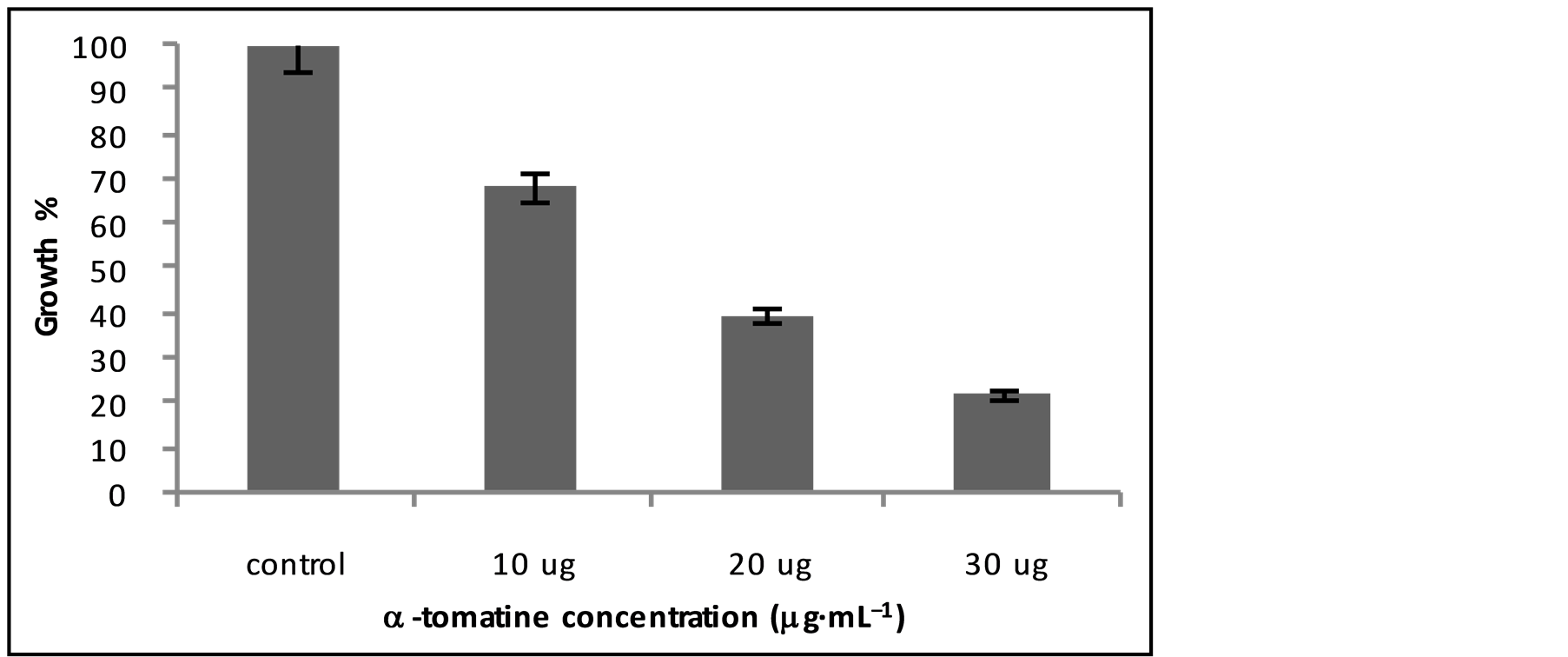
Figure 4. % growth evaluation of M. perniciosa in TDA medium (control) and with adition of alfa-tomatine standard sigma T-2830 in different concentrations (μg·mL−1), with standard deviation.
5. Conclusion
The glycoalcaloid alfa-tomatine, present in S. lycopersicum leaves is able to inhibit growth of Moniliophthora perniciosa fungi as a excellent bio compound to be used for the control of this important phytopathogen, showing important characteristics as an easy extraction method in water, high stability in aqueous media, thermic resistance and selectiveness.
References
- Graniti, A., Durbin, R.D. and Ballio, A. (1988) Phytotoxins and Plant. Pathogenesis. NATO ASI Series, Plenum, New York.
- Strobel, G., Sugawara, F. and Hershenhorn, J. (1992) Pathogens and Their Products Affecting Weedy Plants. Phytoparasitica, 20, 307-323. http://dx.doi.org/10.1007/BF02980853
- Mattinen, L., Nissinen, R., Riipi, T., Kalkkinen, N. and Pirhonen, M. (2007) Host-Extract Induced Changes in the Secretome of the Plant Pathogenic Bacterium Pectobacterium Atrosepticum. Proteomics, 27, 3527-3537. http://dx.doi.org/10.1002/pmic.200600759
- Aime, M.C. and Phillips-Mora, W. (2005) The Causal Agents of Witches’ Broom and Frosty Pod Rot of Cacao (Chocolate, Theobroma cacao) form a New Lineage of Marasmiaceae. Mycologia, 97, 1012-1022. http://dx.doi.org/10.3852/mycologia.97.5.1012
- Griffith, G.W., Bravo-Velasquez, E., Wilson, F.J., Lewis, D.M. and Hedger, J. (1997) Autoecology and Evolution of the Witches’ Broom Pathogen (Crinipellis perniciosa) of Cocoa. In: Blakeman, J.P. and Williamson, B., Eds., The Ecology of Plant Pathogens, CABI International, Wallingford, 245-267.
- Andebrhan, T., Figueira, A., Yamada, M.M., Cascardo, J. and Furtek, D.B. (1999) Molecular Fingerprinting Suggests Two Primary Outbreaks of Witches’ Broom Disease (Crinipellis perniciosa) of Theobroma cacao in Bahia, Brazil. European Journal of Plant Pathology, 105, 167-175. http://dx.doi.org/10.1023/A:1008716000479
- Purdy, L.H. and Schmid, R.A. (1996) Status of Cacao Witches Broom: Biology, Epidemiology, and Management. Annual Review of Phytopathology, 34, 573-594. http://dx.doi.org/10.1146/annurev.phyto.34.1.573
- Paim, V.R.L., Luz, E.D.M.N., Pires, J.L., Silva, S.D.V.M., Souza, J.T., Albuquerque, P.S.B. and Santos, Filho L.P. (2006) Sources of Resistance to Crinipellis perniciosa in Progenies of Cacao Accessions Collected in the Brazilian Amazon. Scientia Agricola, 63, 572-578. http://dx.doi.org/10.1590/S0103-90162006000600011
- de Albuquerque, P.S.B., Silva, S.D.V.M., Luz, E.D.M.N., Pires, J.L., Vieira, A.M.C., Demétrio, C.G.B., Pascholatti, S.F. and Figueira, A. (2010) Novel Sources of Witches’ Broom Resistance (Causal Agent Moniliophthora perniciosa) from Natural Populations of Theobroma cacao L. from the Brazilian Amazon. Euphytica, 172, 125138. http://dx.doi.org/10.1007/s10681-009-0068-4
- Frias, G.A., Purdy, L.H. and Schmidt, R.A. (1991) Infection Biology of Crinipellis perniciosa on Vegetative Flushes of Cacao. Plant Disease, 75, 552-556. http://dx.doi.org/10.1094/PD-75-0552
- Leal, G.A., gomes, L.H., Albuquerque, P.S.B., Tavares, F.C.A. and Figueira, A. (2010) Searching for Moniliophthora perniciosa Pathogenicity Genes. Fungal Biology, 114, 842-854. http://dx.doi.org/10.1016/j.funbio.2010.07.009
- Pinto, L.M.R., Kaneko, T.M. and Ohara, M.T. (2003) Controle Biologico de Qualidade de Produtos Farmaceuticos, Correlatos e Cosméticos. 2nd Edition, Atheneu Ed., São Paulo, 325 p.
- Friedman, M. (2004) Analysis of Biological Active Compounds in Potatoes (Solanum tuberosum), Tomaotes (Lycopersicom esculentum) and Jimson Weed (Datura stramonium) Seeds. Journal of Chromatography A, 1054, 143-155.
- Furlan, S.A., Virmond, L.J., Miers, D.A., Bonatti, M., Gern, R.M.M. and Jonas, R. (1997) Mushroom Strains Able to Grow at High Temperatures and Low pH Values. World Journal of Microbiology & Biotechnology, 13, 689-692. http://dx.doi.org/10.1023/A:1018579123385
- Griffith, G.M. and Hedger, J.N. (2012) A Novel Method for Producing Basidiocarps of the Cocoa Pathogen Crinipellis perniciosa Using a Bran-Vermiculite Médium. European Journal of Plant Pathology, 1.
- Sandrock, R.W. and VanEtten, H.D. (1998) Fungal Sensitivity to and Enzymatic Degradation of the Phytoanticipin α-Tomatine. Phytopathology, 88, 137-143. http://dx.doi.org/10.1094/PHYTO.1998.88.2.137
- Kubow, S. (1993) Lipid Oxidation Products in Food and Atherogenesis. Nutritions Reviews, 51, 33-40. http://dx.doi.org/10.1111/j.1753-4887.1993.tb03064.x
- Lairini, L.K. (1998) Ruiz-Rubio Detoxification of Alfa-Tomatine by Fusarium solani. Mycological Research, 102, 1375-1380. http://dx.doi.org/10.1017/S095375629800656X
- Roddick, J.G. (1974) The Steroidal Glycoalkaloid a-Tomatine. Phytochemistry, 13, 9-25. http://dx.doi.org/10.1016/S0031-9422(00)91261-5
- Abdel-Motaal, F.A., Nassar, S.M.M., El-Zayat, S.A., El-Sayed, M.A. and Ito, S. (2010) Antifungal Activity of Endophytic Fungi Isolated from Egyptian Henbane (hyoscyamus muticus l.) Pak. Journal of Botany, 42, 2883-2894.
- Guyton, B. (2006) Cocoa Market: Supply and Outlook. Chocolate Manufactures Association. Production and Marketing Statistics, 123 p. htpp://www.worldcocoafoundation.org/infoenter/fact-sheet.asp
NOTES
*Corresponding author.