World Journal of Neuroscience
Vol.2 No.2(2012), Article ID:19457,4 pages DOI:10.4236/wjns.2012.22017
Oxytocin in rat nucleus raphe magnus influences pain modulation
![]()
1College of Pharmacy, Xinxiang Medical University, Xixiang, China
2Jiangsu Su Bei People’s Hospital, Yangzhou University, Yangzhou, China
Email: *bcd2009@126.com
Received 17 December 2011; revised 30 January 2012; accepted 16 February 2012
Keywords: Oxytocin; Oxytocin Receptor Antagonist; Nucleus Raphe Magnus; Pain Modulation
ABSTRACT
The main function of Nucleus raphe magnus (NRM) is mostly pain mediation. Our previous study has demonstrated that oxytocin (OXT) regulates antinociception through the central nervous system rather than the peripheral organs, and pain stimulation increases OXT concentration in the NRM. The experiment was designed to investigate OXT in the rat NRM effect on pain modulation. The results showed that 1) pain stimulation increased OXT concentration in NRM perfusion liquid; 2) Intra-NRM microinjection of OXT increased the pain threshold in a dose-dependent manner, whereas intra-NRM microinjection of OXT receptor antagonist, desGly-NH2, d(CH2)5[D-Tyr2, Thr-sup-4]OVT decreased the pain threshold; 3) NRM pre-treatment with OXT receptor antagonist completely attenuated the pain threshold increase induced by intra-NRM administration of OXT. The data suggested that OXT in NRM was involved in antinociception via OXT receptors.
1. INTRODUCTION
Nucleus raphe magnus (NRM), located directly rostral to the raphe obscurus, is afferently stimulated from axons in the spinal cord and cerebellum [1]. The NRM receives descending afferents from the periaqueductal gray (PAG), hypothalamic paraventricular nucleus (PVN), central nucleus of the amygdala, lateral hypothalamic area, parvocellular reticular nucleus and the prelimbic, infralimbic, medial and lateral precentral cortices [2], which brain areas influence the main function of NRM [3]. NRM sends projections to the dorsal horn of the spinal cord to directly inhibit nociceptive neurotransmission [2].
Oxytocin (OXT), a nonapeptide posterior hormone of the pituitary, is mainly synthesized and secreted in the hypothalamic supraoptic nucleus (SON) and PVN. This hormone, combined with an apparent carrier protein (neurophysin), is transported along the hypothalamohypophyseal pathway to the neurohypophysis, where it is stored for subsequent release [4]. The remarkable functions of OXT include uterine contraction during parturition, milk-ejection reflex during lactation, cardiovascular regulation, sex activity, learning and memory [4]. Recently, it has been demonstrated that OXT is related with pain modulation. Intraventricular injection (icv) of OXT increases the pain threshold, whereas administration of anti-OXT serum (icv) decreases the pain threshold [5,6].
OXT is very similar to arginine vasopressin (AVP) in the structure, synthesis, source and distribution [7]. Intra-NRM microinjection of AVP increases the pain threshold, and local administration of vasopressin receptor antagonist decreases the pain threshold [8]. Through method of generated an OXT receptor-receptor mouse, in which part of the OXT receptor gene is replaced with Venus cDNA (a variant of yellow fluorescent protein), is to more precisely characterize OXT receptor-expressing neurons, Yoshida et al. has found that there are many OXT recaptors in the raphe nuclei including NRM [9]. Pain stimulation can change OXT concentration in the NRM tissue [5,6]. However, it is still not clear how OXT in the NRM influences pain modulation. The present study was tried to investigate OXT in the rat NRM effect on pain modulation.
2. MATERIALS AND METHODS
2.1. Animals
Adult male Sprague-Dawley rats weighing 180 - 220 g, which were obtained from Animal Center of Yangzhou University, Yangzhou, Jiangsu, China, were housed with food and water available ad libitum in a colony room under controlled temperature, humidity and a 12 hours light/dark cycle (light at 6:00 AM and dark at 6:00 PM). All the procedures were approved by Animal Care Committee of Yangzhou University and conducted according to the guidelines of the International Association for the Study of Pain [10].
2.2. Materials
OXT and desGly-NH2, d(CH2)5[D-Tyr2, Thr-sup-4]OVT (a high specific OXT receptor antagonist) were obtained from Peninsula Lab, San Carlos, CA, USA. 125Iodine was obtained from Amersham Pharmacia, Buckinghamshire, UK. The other chemicals were bought from Sigma Co., St. Louis, MO, USA.
Rabbit anti-rat OXT serum was made by Department of Neurobiology, Second Military Medical University, Shanghai, China. The reactivity of the anti-OXT serum specificity was over 99.9% with the peptide itself and less than 0.1% with other similar peptides such as AVP and vasotocin. The dilution of the antiserum was 1:20,000 for radioimmunoassay.
2.3. Surgery
With the reference of Pellegrino L.J. rat brain atlas [11], we implanted a stainless steel guide cannula with 0.5 mm outer diameter for nucleus microinjection or push-pull perfusion into the NRM (AP 8.4 mm, LR 0.5 mm, H 8.2 mm) under the pentobarbital sodium (35 mg/kg, intraperitoneal injection) anesthesia by the stereotaxic apparatus (Jiangwan I-C, Shanghai, China). The guide cannula was fixed to the skull by dental acrylic. All the operations were carried out in the aseptic condition and the rats would be allowed to recover for at least 14 days after the surgery.
2.4. Nociceptive Tests
All animals were tested under the condition of free activity in the small cages (30 cm in diameter, 25 cm in height) from 8:00 to 10:00 am. The potassium iontophoresis inducing tail-flick served as pain stimulus. The small round wet cotton (radius 1.8 mm and area about 10 mm2) with the potassium iontophoresis was set on the tail skin. The cotton was exposed to direct electrical current, and the anode led the potassium iontophoresis to permeate the tail skin. If the current was strong enough, the permeated potassium iontophoresis resulted in the animal feeling the pain stimulation. The intensity of current at the moment of the response was recorded as the pain threshold, which was expressed as mA (WQ-9E Pain Threshold Measurer, Shanghai, China). The area size of electrode and concentration of potassium was all the same condition for evaluation of animal pain (or response to noxious stimulation) using by electric current. The duration between consecutive stimuli is 10 min, and the pain stimulus was terminated at once when the rat showed response to this stimulus.
2.5. Pain Stimulation
The direct electrical current produced by the anode of pain threshold measurer induced the acute pain to the tail skin by the potassium iontophoresis. The intensity was fixed to 1.2 ~ 1.4 times of pain threshold (0.6 - 0.7 mA) over 1 min. For the sham stimulation, the animal has been done as the animal with pain stimulation except not giving the direct electrical current.
2.6. NRM Push-Pull Perfusion
A stainless steel needle with 0.3 mm outer diameter was directly implanted into the NRM guide cannula, 1 mm beyond the tip, for pushing artificial cerebral spinal fluid (ACSF, containing 0.1 M NaCl, 1.0 mM KH2PO4, 4.0 mM KCl, 2.0 mM MgSO4, 2.0 mM CaCl2, 2.1 mM NaHCO3, and 8.0 mM Glucose) into the NRM, and pulling the ACSF out by the guide cannula. The entering ACSF was heated at 37˚C before pushing to the NRM of the consciously rat (0.05 ml/min), and the extracted fluid was collected at 0˚C every 10 min. The samples were stored at –80˚C for detection.
For NRM administration of OXT or OXT receptor antagonist, the solution, which contained the different dose of OXT or OXT receptor antagonist solved in 1.0 ml ACSF, was purified extract cross-reacted in parallel fashion with the perfusion liquid and was push into the NRM over 20 min during NRM push-pull perfusion. In fact, the perfusion style and method did not alter except for opening two cross-reacted switches in the parallel tube, which contained the different dose of OXT or OXT receptor antagonist solved in 1.0 ml perfusion liquid (ACSF).
2.7. NRM Microinjection
On the day of experiments, a stainless steel needle with 0.3 mm diameter for NRM microinjection was inserted into the guide cannula, 1 mm beyond the tip. The OXT or OXT receptor antagonist with 1-μl ACSF was gently injected into NRM over 10 min. For the control animal, only 1-μl ACSF was given.
2.8. Radioimmunoassay
OXT concentration was detected by specific antiserum. The peptide was labeled 125Iodine by the chloramines-T method, and iodinated peptide was purified by Sephadex G-50 with the assay sensitivity of 1.0 pg/tube. The intraassay coefficient of variation was less than 3.6%, and inter-assay coefficient of variation was less than 6.7%, respectively.
2.9. Histological Verification
After the rat was sacrificed under a high dose of pentobarbital sodium (80 mg/kg, intraperitoneal injection), we injected the black dye into the NRM to observe the histological location of NRM push-pull perfusion or microinjection. The data were excluded from analysis if the positions were not accurate (Figure 1), which pathological photo was same as that we reported before [8]. On the other hand, the intra-NRM injected black dye could prove that OXT or OXT antagonist limited to remain in the NRM after microinjection.
2.10. Statistical Analysis
Although referencing to the rat brain atlas, the little different induced by the rat individual difference was unavoidable which induced the gap in the OXT basal value of each rat NRM perfusion liquid in a same group. More rats in each group are necessary to meet the statistically significant if using the absolute values.
OXT concentration in NRM perfusion liquid was converted into the relative value comparing with the value itself before the treatment for the statistical analysis. Compared with its value, each value should be 0 at time before the treatment. The calculation was as follows:

All values were expressed as mean ± standard error of the mean (SEM) and were analyzed between groups by Origin v7.5 (Originlab Corporation, Northampton, MA, USA), which two-way analysis of variance (ANOVA), followed by the Bonferroni test and one-way ANOVA followed by Dunnett test and Newmann-Keuls test. p < 0.05 was considered statistically significant.
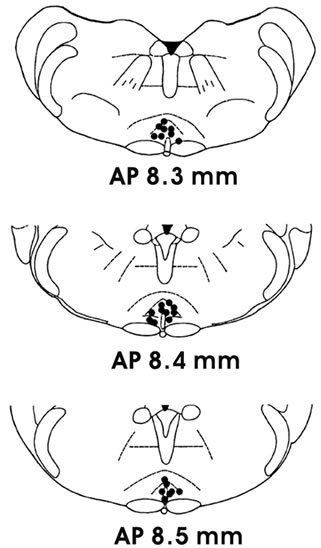
Figure 1. Histological verification of nucleus raphe magnus (NRM) push-pull perfusion or microinjection.
3. RESULTS
3.1. Pain Stimulation Increased OXT Concentration in NRM Perfusion Liquid
The basal concentration of OXT in NRM perfusion liquid was 5.2 ± 1.7 pg/ml, respectively. Giving the rat 1 min pain stimulation, OXT concentration in NRM perfusion liquid increased to 237.8% ± 42.6% at 10 min (p < 0.001), 145.6% ± 29.4% at 20 min (p < 0.001) and 63.2% ± 33.7% at 30 min; But OXT concentration in NRM perfusion liquid did not change in the control group (Figure 2).
3.2. Intra-NRM Microinjection of OXT Increased the Pain Threshold
Intra-NRM microinjection of 10 ng OXT/1 μl ACSF increased the pain threshold from 4.9 ± 0.4 mA to 9.3 ± 1.1 mA at 10 min (p < 0.001), 8.1 ± 1.0 mA at 20 min (p < 0.001) and 6.2 ± 0.6 mA at 30 min (p < 0.01); and intra-NRM microinjection of 5 ng OXT/1 μl ACSF increased the pain threshold from 5.1 ± 0.3 mA to 7.7 ± 1.1 mA at 10 min (p < 0.001) and 5.9 ± 0.3 mA at 20 min (p < 0.05); but only intra-NRM microinjection of 1 μl ACSF (Control group) did not change the pain threshold (Figure 3).
3.3. Intra-NRM Microinjection of OXT Receptor Antagonist Decreased the Pain Threshold
Intra-NRM microinjection of 1.0 μg OXT receptor antagonist (desGly-NH2, d(CH2)5[D-Tyr2, Thr-sup-4] OVT)/1 μl ACSF decreased the pain threshold from 5.0
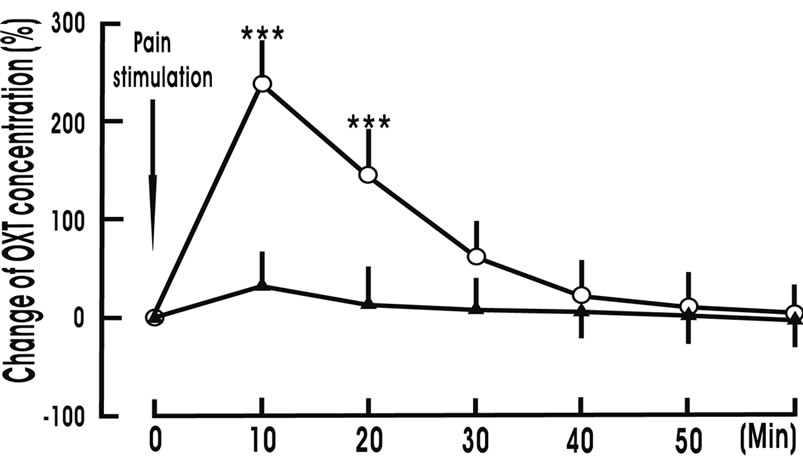
Figure 2. Pain stimulation increases oxytocin (OXT) concentration in nucleus raphe magnus perfusion liquid. Change OXT concentration (%) denotes (present value—value before treatment)/value before treatment × 100. Pain stimulation denotes the beginning of pain stimulation. Pain stimulation group (○, n = 8): The animal was given 1 min pain stimulation during experiment; Control group (▲, n = 8): The animal was give 1 min sham stimulation during experiment. N indicates the animal number in each group. The data are expressed as mean ± standard error mean (SEM). ***p < 0.001 is used for the compareson of the change of OXT concentration from pain stimulation group and control group.
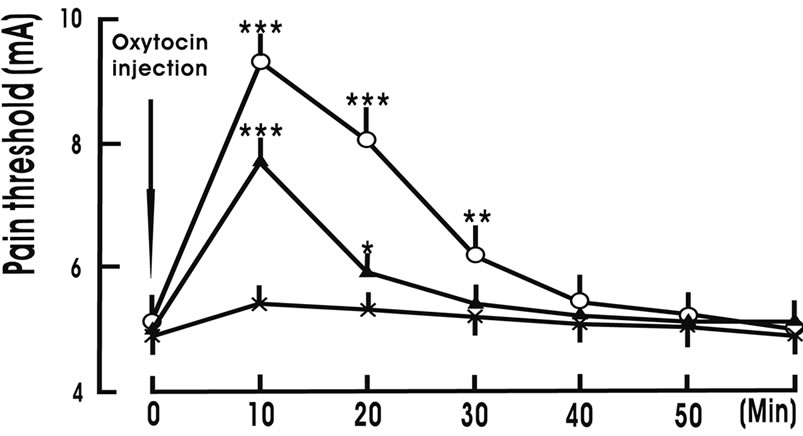
Figure 3. Intra-nucleus raphe magnus (NRM) microinjection of oxytocin increases the pain threshold. Oxytocin injection denotes the beginning of intra-NRM microinjection of oxytocin. Oxytocin 10 ng group (○, n = 8): 10 ng oxytocin/1 μl artificial cerebral spinal fluid (ACSF) was injected into the NRM over 10 min; Oxytocin 5 ng group (▲, n = 8): 5 ng oxytocin/1 μl ACSF was injected into the NRM over 10 min; Control group (×, n = 8): Only 1 μl ACSF was injected into the NRM over 10 min. N indicates the animal number in each group. The data are expressed as mean ± standard error mean (SEM). *p < 0.05, **P < 0.01 or ***p < 0.001 is used for the comparison of the pain threshold from oxytocin 10 ng group (or oxytocin 5 ng group) and control group.
± 0.4 mA to 3.2 ± 0.5 mA at 10 min (p < 0.001), 3.6 ± 0.5 mA at 20 min (p < 0.001) and 4.0 ± 0.6 mA at 30 min (p < 0.01); and intra-NRM microinjection of 0.5 μg OXT receptor antagonist/1 μl ACSF decreased the pain threshold from 4.9 ± 0.5 mA to 4.0 ± 0.3 mA at 10 min (p < 0.001) and 4.6 ± 0.4 mA at 20 min; but only intra-NRM microinjection of 1 μl ACSF (Control group) did not change the pain threshold (Figure 4).
3.4. Pretreatment with OXT Receptor Antagonist Blocked the Pain Threshold Increase Induced by Intra-NRM Microinjection of OXT
Intra-NRM microinjection of OXT increased the pain threshold, whereas intra-NRM microinjection of OXT receptor antagonist decreased the pain threshold. NRM pretreatment with 1.0 μg OXT receptor antagonist completely attenuated the pain threshold increase induced by intra-NRM microinjection of 10 ng OXT (Figure 5).
4. DISCUSSION
Having been used in many laboratories, the classical hot plate/tail-plate test to noxious heat, and hind paw withdrawal to noxious heating are very important to study the hyperalgesia/allodynia in animals with persistent/chronic pain. However, these tests have some problems in the special research for pain. For example, the latency (pain threshold) shows instability after the animal is tested many times, especially in the experiments that the duration between consecutive stimuli is short. On the other hand, it
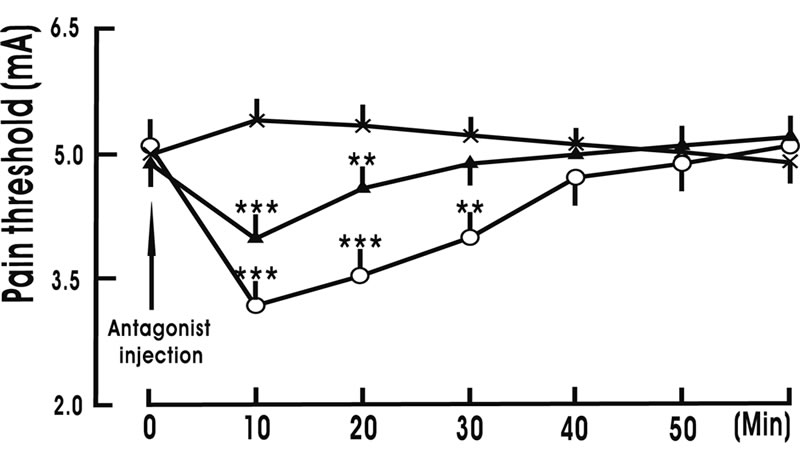
Figure 4. Intra-nucleus raphe magnus (NRM) microinjection of oxytocin receptor antagonist decreases the pain threshold. Antagonist injection denotes the beginning of intra-NRM microinjection of oxytocin receptor antagonist. Antagonist 1.0 μg group (○, n = 8): 1.0 μg oxytocin receptor antagonist (desGly-NH2, d(CH2)5[D-Tyr2, Thr-sup-4]OVT)/1 μl artificial cerebral spinal fluid (ACSF) was injected into the NRM over 10 min; Antagonist 0.5 μg group (▲, n = 8): 0.5 μg oxytocin receptor antagonist/1 μl ACSF was injected into the NRM over 10 min; Control group (×, n = 8): Only 1 μl ACSF was injected into the NRM over 10 min. N indicates the animal number in each group. The data are expressed as mean ± standard error mean (SEM). **p < 0.01 or ***p < 0.001 is used for the comparison of the pain threshold from antagonist 1.0 μg group (or antagonist 0.5 μg group) and control group.
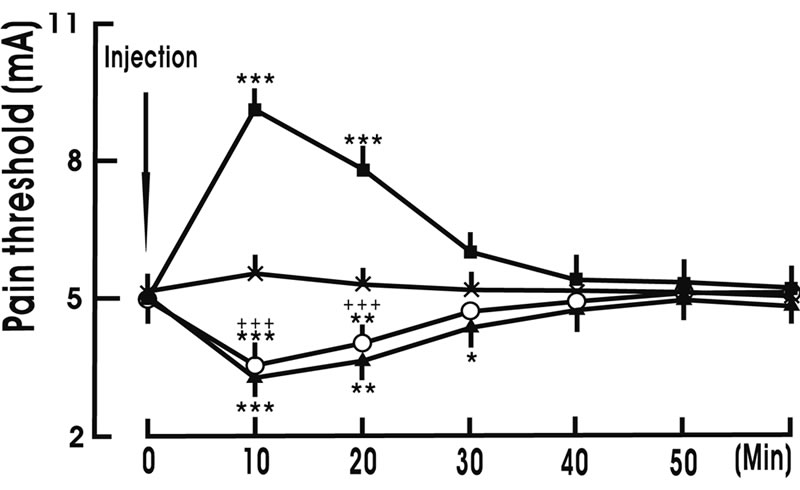
Figure 5. Pretreatment with oxytocin receptor antagonist blocked the pain threshold increase induced by intra-nucleus raphe magnus (NRM) microinjection of oxytocin. ACSF denotes artificial cerebral spinal fluid. ACSF + ACSF group (×, n = 8): NRM pretreatment with 1 μl ACSF, and then NRM was given 1 μl ACSF; ACSF + OXT group (■, n = 8): NRM pretreatment with 1 μl ACSF, and then NRM was given 10 ng OXT/1 μl ACSF; Antagonist + ACSF group (▲, n = 8): NRM pretreatment with 1.0 μg oxytocin receptor antagonist (desGly-NH2, d(CH2)5[D-Tyr2, Thr-sup-4]OVT)/1 μl ACSF, and then NRM was given 1 μl ACSF; Antagonist + OXT group (○, n = 8): NRM pretreatment with 1.0 μg xytocin receptor antagonist/1 μl ACSF, and then NRM was given 10 ng OXT/1 μl ACSF. N indicates the animal number in each group. The data are expressed as mean ± standard error mean (SEM). *p < 0.05, **p < 0.01 or ***p < 0.001 is used for the comparison of the pain threshold from ACSF + ACSF group and other group. +++ p < 0.001 is used for the comparison of the pain threshold from ACSF + OXT group and antagonist + OXT group.
may cause the animal stress when the animal is removed from the hot plate after the test [12]. Many methods for pain threshold measurement including the hot plate test to noxious heat and tail-flick test to noxious potassium iontophoresis, have been compared in our laboratory since we studied pain modulation and acupuncture analgesia in 1954 [13]. We have found that the tail-flick test to noxious potassium iontophoresis is good, relatively, to keep the basic pain threshold stability in studying pain modulation and acupuncture analgesia, and is easy, specially, to control the animal extra-stimulation under the condition of free activity [14]. Like the classical methods for pain threshold measurement such as the hot plate/tailplate test to noxious heat and hind paw withdrawal to noxious heating, the tail flicking induced by potassium inotophoresis is not merely a spinal reflex [13]. Of course, it is better that the tail-flick test to noxious potassium iontophoresis can combine with classical methods for pain threshold measurement including the hot plate test to noxious heat in one experiment for pain research.
Nucleus raphe magnus (NRM) is a serotonergic nucleus located in the rostral ventromedial medulla of the brainstem. Axons of the NRM project to the spinal cord [1], terminating primarily in the dorsal horn [2]. Brainstem nuclei that project to the dorsal horn of the spinal cord can function to inhibit afferent nociceptive transmission [7,15,16]. Activation of these descending antinociceptive pathways may be triggered by physiological stimuli [17] as well as by pharmacological agents [18]. Antinociception involving the NRM has been studies after either electrical stimulation or direct administration of pharmacological agents [19,20,21]. NRM is a key neural structure for pain modulation.
Oxytocin (OXT) is a peptide of nine amino acids (a nonapeptide): its systematic name is cys - tyr - ile - gln - asn - cys - pro - leu - gly - NH2, which structure is very similar to that of arginine vasopressin (AVP) (cys - tyr - phe - gln - asn - cys - pro - arg - gly - NH2). OXT and AVP secreted from the pituitary gland cannot re-enter the brain because of the blood-brain barrier. Instead, the behavioral effects of OXT and AVP are thought to reflect release from centrally projecting OXT and AVP neurons, different from those that project to the pituitary gland, or which are collaterals from them. The similarity of the two peptides can cause some cross-reactions: OXT has a slight antidiuretic function, and high levels of AVP can cause uterine contractions. OXT receptors are expressed by neurons in many parts of the brain and spinal cord, including the amygdala, ventromedial hypothalamus, septum, nucleus accumbens and brainstem [22]. Intraventricular injection of OXT or AVP increases the nociceptive threshold, whereas local administration of anti-OXT serum or anti-AVP serum decreases the nociceptive threshold; pain stimulation can influence not only OXT but also AVP concentrations in some brain nuclei including the NRM [5,6,23,24]. Intra-NRM microinjection of AVP increases the pain threshold, whereas intra-NRM microinjection of vasopressin receptor antagonist decreases the pain threshold; pain stimulation induces the NRM release of AVP [8]. AVP in NRM plays a role in pain modulation [25]. Similarly as AVP, OXT in NRM may be involved in pain modulation.
Some experiments have proven that NRM neurons project to the hypothalamic paraventricular nucleus (PVN) and supraoptic nucleus (SON) [26,27]. Both PVN and SON, which are the main neural structure to syntheses and secret OXT [3], can regulate the pain process [28- 31]. The present study showed that 1) pain stimulation increased OXT concentration in NRM perfusion liquid; 2) Intra-NRM microinjection of OXT increased the pain threshold in a dose-dependent manner, whereas intra-NRM microinjection of OXT receptor antagonist decreased the pain threshold; 3) NRM pre-treatment with OXT receptor antagonist completely attenuated the pain threshold increase induced by intra-NRM microinjection of OXT. The data suggested that OXT in NRM was involved in antinociception. Although the part of our present results, which intra-NRM injection of OXT increased the pain threshold whereas local administration of OXT receptor antagonist decreased the pain threshold, was same as Wang’s report [32], we discovered the points that NRM could release OXT to regulate pain process via OXT receptors.
The NRM receives descending afferents from many brain areas such as PVN, periaqueductal gray (PAG) [2] that are related with pain modulation [3]. The NRM releases enkephalin and endorphin when stimulated [25]. Many experiments have indirectly pointed that OXT may influence the endogenous opiate peptide system [32-35]. Enkephalins co-exist with OXT in nerve terminals [36, 37]. The κ- and δ-opioid receptors are involved in the OXT antinociceptive effect in formalin-induced pain response in mice [38]. In the spinal cord, OXT can influence endogenous opiate peptides to participate in pain modulation [39,40]. The relationship between OXT and endogenous opiate peptide system may be the base to understand the mechanism of OXT antinociceptive role in NRM.
After examination of the Venus expression in the raphe nuclei including NRM, Yoshida et al. has revealed that about one-half of tryptophan hydroxylase-immunoreactive neurons are positive for Venus. The data suggest that OXT plays a potential role in the modulation of serotonin release via OXT receptors in the raphe nuclei [9]. The interaction between OXT and serotonin in the NRM may be another way of OXT antinociceptive role in NRM.
In conclusion, the present study makes it clear that 1) NRM releases OXT during pain stimulation; 2) endogenous OXT influences the pain threshold; 3) OXT recaptors are related with OXT regulating pain process. The data suggested that OXT in NRM was involved in pain modulation via OXT receptors.
5. ACKNOWLEDGEMENTS
This work was supported by Xinxiang Medical University, Jiangsu Su Bei People’s Hospital and and grants from National Natural Science Foundation of China (81100956/H0912).
REFERENCES
- Bowker, R.M., Westlund, K.N., Sullivan, M.C. and Coulter, J.D. (1982) Organization of descending serotonergic projections to the spinal cord. Progress in Brain Research, 57, 239-265. doi:10.1016/S0079-6123(08)64132-1
- Jones, S.L. and Light, A.R. (1990) Electrical stimulation in the medullary nucleus raphe magnus inhibits noxious heat-evoked protein-like immunoreactivity in the lumbar spinal cord. Brain Research, 530, 335-338. doi:10.1016/0006-8993(90)91306-2
- Hermann, D.M., Luppi, P.H., Peyron, C., Hinckel, P. and Jouvet, M. (1997) Afferent projections to the rat nuclei raphe magnus, raphe pallidus and reticularis gigantocellularis pars alpha demonstrated by iontophoretic application of choleratoxin (subunit b). Journal of Chemical Neuroanatomy, 13, 1-21. doi:10.1016/S0891-0618(97)00019-7
- McEwen, B.B. (2004) The role of vasopressin and oxytocin in memory processing. Elsevier Science, Amsterdam.
- Yang, J., Yang, Y., Chen, J.M., Liu, W.Y., Wang, C.H. and Lin, B.C. (2007) Effect of oxytocin on acupuncture analgesia in the rat. Neuropeptides, 41, 285-292. doi:10.1016/j.npep.2007.05.004
- Yang, J., Yang, Y., Chen, J.M., Liu, W.Y., Wang, C.H. and Lin, B.C. (2007) Central oxytocin enhances antinociception in the rat. Peptides, 28, 1113-1119. doi:10.1016/j.peptides.2007.03.003
- Willis, W.D. (1988) Anatomy and physiology of descending control of nociceptive responses of dorsal horn neurons: Comprehensive review. In: Fields, H.L. and Besson, J.M., Eds., Progress in Brain Research, 77, Elsevier, Amsterdam, 1-29. doi:10.1016/S0079-6123(08)62776-4
- Yang, J., Chen, J.M., Liu, W.Y., Song, C.Y. and Lin, B.C. (2006) Effect of arginine vasopressin in the nucleus raphe magnus on antinociception in the rat. Peptides, 27, 2224-2229. doi:10.1016/j.peptides.2006.03.017
- Yoshida, M., Takayanagi, Y., Inoue, K., Kimura, T., Young, L.J., Onaka, T. and Nishimori, K. (2009) Evidence that oxytocin exerts anxiolytic effects via oxytocin receptor expressed in serotonergic neurons in mice. Journal of Neuroscience, 29, 2259-2271. doi:10.1523/JNEUROSCI.5593-08.2009
- Zimmermann, M. (1983) Ethical guidelines for investigations of experimental pain in conscious animal. Pain, 16, 109-110. doi:10.1016/0304-3959(83)90201-4
- Pellegrino, K.J., Pellegrino, A.S. and Cushman, A.J. (1979) An atereotaxic atlas of the brain. Plenum, New York.
- Zhang, E.Q. (1990) Chinese acupuncture and moxibustion. Publishing House of Shanghai College of Traditional Chinese Medicine, Shanghai.
- Chen, Y., Wang, Y. and Yin, Q. (1991) The role of paraventricular nucleus of hypothalamus in acupuncture analgesia in rats. Acupuncture Research, 16, 32-38.
- Yang, J., Yang, Y., Chen, J.M., Xu, H.T., Liu, W.Y., Wang, C.H. and Lin, B.C. (2007) Arginine vasopressin is an important regulator in antinociceptive modulation of hypothalamic paraventricular nucleus in the rat. Neuropeptides, 41, 165-176. doi:10.1016/j.npep.2006.12.005
- Basbaum, A.I. and Fields, H.L. (1979) The origin of descending pathways in the dorsolateral funiculus of the spinal cord of the cat and rat: Further studies on the anatomy of pain modulation. The Journal of Comparative Neurology, 187, 513-532. doi:10.1002/cne.901870304
- Sandkuhler, J. (1996) The organization and function of endogenous antinociceptive system. Progress in Neurobiology, 50, 49-81.
- Millan, M.J., Przewlocki, R. and Herz, A. (1980) A non-beta-endorphinergic adenohypophyseal mechanism is essential for an analgetic response to stress. Pain, 8, 343-353.
- Gogas, K.R., Presley, R.W., Levine, J.D. and Basbaum, A.I. (1991) The antinociceptive action of supraspinal opioids results from an increase in descending inhibitory control: Correlation of nociceptive behavior and c-fos expression. Neuroscience, 42, 617-628. doi:10.1016/0306-4522(91)90031-I
- Brodie, M.S. and Produfit, H.K. (1986) Antinociception induced by local injections of carbachol into the nucleus raphe magnus in rats: Alteration by intrathecal injection of monoaminergic antagonists. Brain Research, 371, 70- 79. doi:10.1016/0006-8993(86)90811-5
- Oleson, T.D., Twombly, D.A. and Liebeskind, J.C. (1978) Effects of pain-attenuating brain stimulationand morphine on electrical activity in the raphe nuclei of the awake rat. Pain, 4, 211-230. doi:10.1016/0304-3959(77)90134-8
- Proudfit, H.K. and Anderson, E.G. (1975) Mprphine analgesia: Blockade by raphe magnus lesions. Brain Research, 98, 612-618. doi:10.1016/0006-8993(75)90380-7
- Ross, H.E., Cole, C.D., Smith, Y., Neumann, I.D., Landgraf, R., Murphy, A.Z. and Young, L.J. (2009) Characterization of the oxytocin system regulating affiliative behavior in female prairie voles. Neuroscience, 162, 892- 903. doi:10.1016/j.neuroscience.2009.05.055
- Yang, J., Song, C.Y., Liu, W.Y. and Lin, B.C. (2006) Only through the brain nuclei, arginine vasopressin regulates antinociception in the rat. Peptides, 27, 3341-3346. doi:10.1016/j.peptides.2006.08.019
- Yang, J., Yang, Y., Wang, C.H., Wang, G., Xu, H.T., Liu, W.Y. and Lin, B.C. (2009) Effect of arginine vasopressin on acupuncture analgesia in the rat. Peptides, 30, 241-247. doi:10.1016/j.peptides.2008.10.013
- Yang, J., Yuan, H.F., Chu, J.G., Yang, Y., Xu, H.T., Wang, G., Liu, W.Y. and Lin, B.C. (2009) Arginine vasopressin antinociception in the rat nucleus raphe magnus is involved in the endogenous opiate peptide and serotonin system. Peptides, 30, 1366-1371. doi:10.1016/j.peptides.2009.03.014
- Kiss, J.Z., Voorhuis, T.A., van Eekelen, J.A., de Kloet, E.R. and de Wied, D. (1997) Organization of vasotocin-immunoreactive cells and fibers in the canary brain. The Journal of Comparative Neurology, 263, 347-364. doi:10.1002/cne.902630304
- Larsen, P.J., Hay-Schmidt, A., Vrang, N. and Mikkelsen, J.D. (1996) Origin of projections from the midbrain raphe nuclei to the hypothalamic paraventricular nucleus in the rat: A combined retrograde and anterograde tracing study. Neuroscience, 70, 963-988. doi:10.1016/0306-4522(95)00415-7
- Yang, J., Chen, J.M., Yang, Y., Liu, W.Y., Song, C.Y. and Lin, B.C. (2008) Investigating the role of hypothalamic paraventricular nucleus in nociception of the rat. International Journal of Neuroscience, 118, 473-485. doi:10.1080/00207450601123563
- Yang, J. and Lin, B.C. (1992) Hypothalamic paraventricular nucleus plays a role in acupuncture analgesia through the central nervous system in the rat. Acupuncture & Electro-Therapeutics Research, 17, 209-220.
- Yang, J., Yang, Y., Chen, J.M., Liu, W.Y., Wang, C.H. and Lin, B.C. (2008) Effect of hypothalamic supraoptic nucleus on acupuncture analgesia in the rat. Brain Research Bulletin, 75, 681-686. doi:10.1016/j.brainresbull.2007.11.004
- Yang, J., Yang, Y., Chen, J.M., Liu, W.Y., Wang, C.H. and Lin, B.C. (2008) Investigating the role of the hypothalamic supraoptic nucleus in nociception in the rat. Life Science, 82, 166-173. doi:10.1016/j.lfs.2007.10.023
- Wang, J.W., Lundeberg, T. and Yu, L.C. (2003) Antinociceptive role of oxytocin in the nucleus raphe magnus of rats, an involvement of mu-opioid receptor. Regulary Peptides, 115, 153-159. doi:10.1016/S0167-0115(03)00152-6
- Arletti, R., Benelli, A. and Bertolini, A. (1993) Influence of oxytocin on nociception and morphine antinociception. Neuropeptides, 24, 125-129. doi:10.1016/0143-4179(93)90075-L
- Micevych, P. and Elde, R. (1980) Relationship between enkephalinergic neurons and the vasopressin-oxytocin neuroendocrine system of the cat: An immunohistochemical study. The Journal of Comparative Neurology, 190, 135-146. doi:10.1002/cne.901900109
- You, Z.D., Li, J.H., Song, C.Y., Wang, C.H. and Lu, C.L. (2003) Chronic morphine treatment inhibits oxytocin synthesis in rats. Neuroreport, 11, 3113-3116. doi:10.1097/00001756-200009280-00015
- Martin, R. and Voigt, K.H. (1981) Enkephalins co-exist with oxytocin and vasopressin in nerve terminals of rat neurohypophysis. Nature, 289, 502-504. doi:10.1038/289502a0
- Nordmann, J.J., Cazalis, M., Dayanithi, G., Castanas, E., Giraud, P., Legros, J.J. and Louis, F. (1986) Are opioid peptides co-localized with vasopressin or oxytocin in the neural lobe of the rat? Cell and Tissue Research, 246, 177-182. doi:10.1007/BF00219015
- Reeta, K.H., Mediratta, P.K., Rathi, N., Jain, H., Chugh, C. and Sharma, K.K. (2006) Role of kappaand deltaopioid receptors in the antinociceptive effect of oxytocin in formalin-induced pain response in mice. Regulatory Peptides, 135, 85-90. doi:10.1016/j.regpep.2006.04.004
- Millan, M.J., Millan, M.H., Członkowski, A. and Herz, A. (1984) Vasopressin and oxytocin in the rat spinal cord: distribution and origins in comparison to [Met]enkephalin, dynorphin and related opioids and their irresponsiveness to stimuli modulating neurohypophyseal secretion. Neuroscience, 13, 179-187. doi:10.1016/0306-4522(84)90268-9
- Yang, J. (1994) Intrathecal administration of oxytocin induces analgesia in low back pain involving the endogenous opiate peptide system. Spine, 19, 867-871. doi:10.1097/00007632-199404150-00001
NOTES
*Corresponding author.