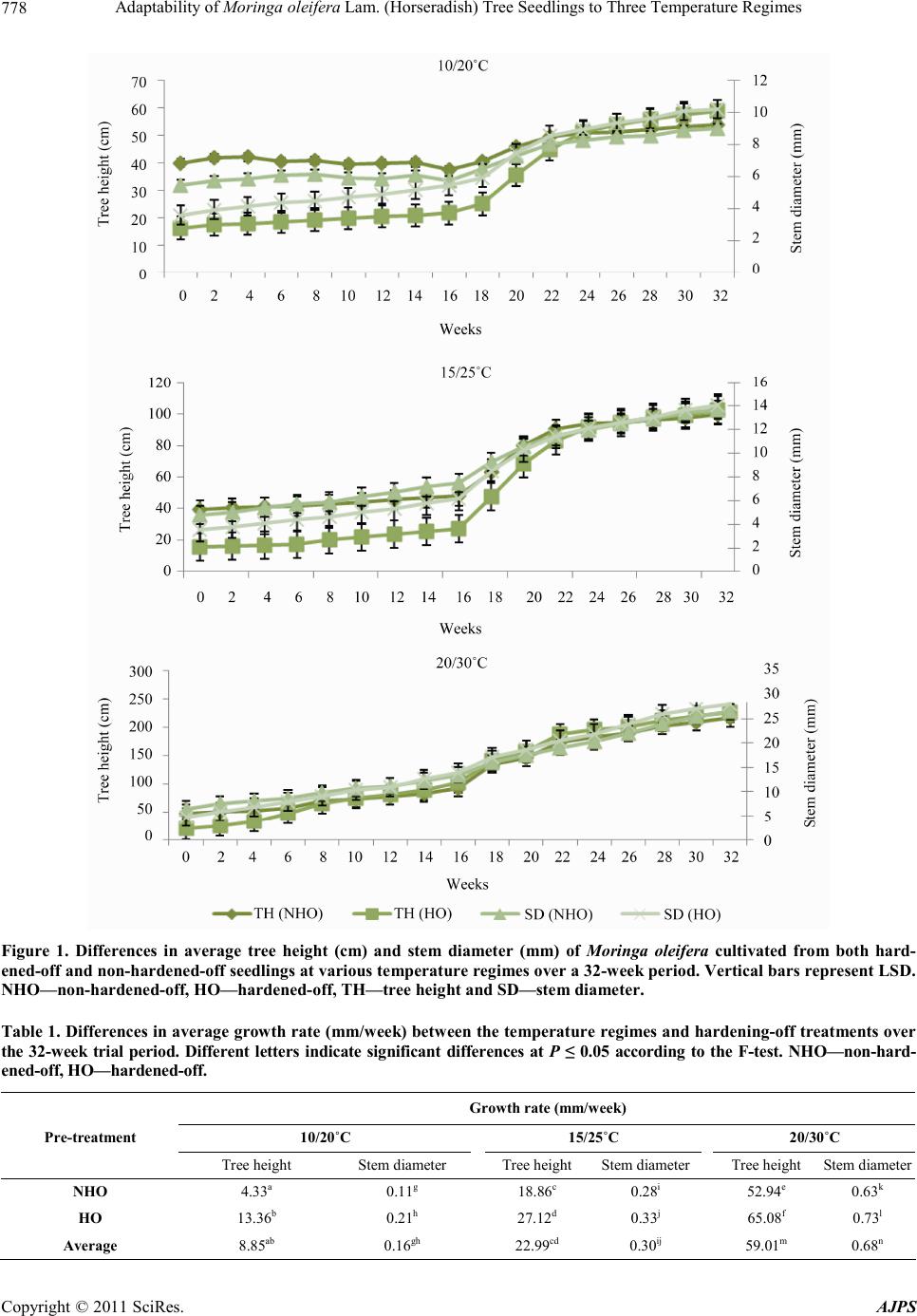
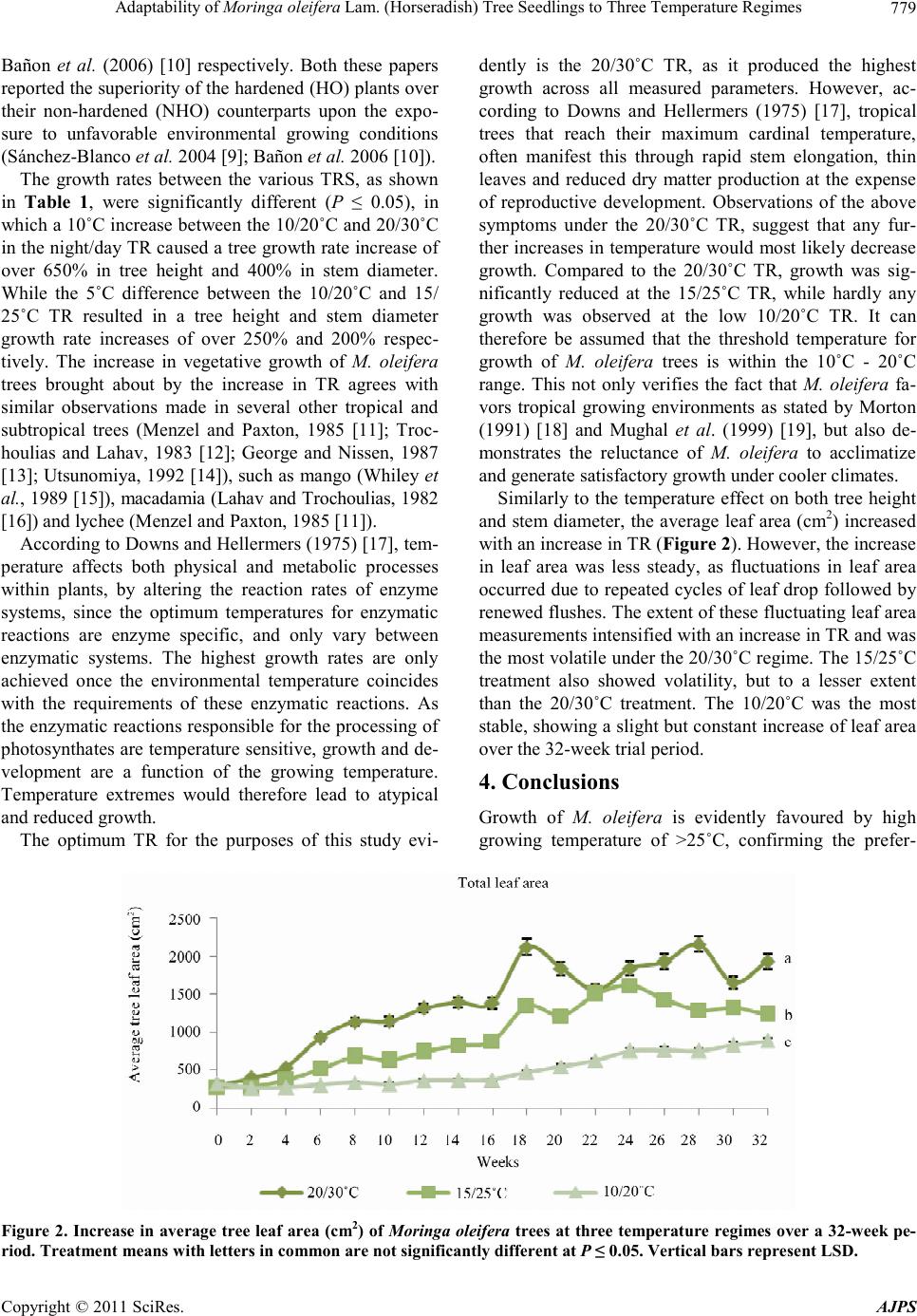
Adaptability of Moringa oleifera Lam. (Horseradish) Tree Seedlings to Three Temperature Regimes
780
ence of M. oleifera towards tropical growing environ-
ments. This was confirmed by the temperature treatment
results, where trees at the 20/30˚C TR revealed the high-
est growth rates for both tree height and stem thickening.
In addition, the 20/30˚C temperature treatment, although
variable, consistently had the highest leaf area over the
entire trial period. As the effect of additional, higher TRS
were not investigated in this study, the possibility of im-
proved growth at even higher temperatures cannot be
excluded. Tropical climates are therefore ideal for the
cultivation of M. oleifera, however satisfactory growth
during the hot summer months in sub-tropical climates is
achievable, if the winters are mild, as trees are frost ten-
der. The hardening-off of the seedlings prior to trans-
planting has proven to increase the growth rate of both
tree height and stem diameter across all three TRS. The
hardening off process is highly recommended for M.
oleifera trees, especially if intended cultivation is in be-
low optimal temperature environments.
REFERENCES
[1] S. A. A. Jahn, “Using Moringa oleifera Seeds as Coa-
gulant in Developing Countries,” Journal of the Ameri-
can Water Works Association, Vol. 80, No. 6, 1988, pp.
43-50.
[2] G. Folkard, J. Sutherland and R. Shaw, “Water Clarifica-
tion Using Moringa oleifera Coagulant. Water and Envi-
ronmental Health at London and Loughborough (WELL),”
Loughborough University, Loughborough, 1999, pp. 109-
112.
[3] U. Rashid, F. Anwar, B. R. Moser and G. Knothe, “Mor-
inga oleifera Oil: A Possible Source of Biodiesel,” Bio-
resource Technology, Vol. 99, No. 17, 2008, pp. 8175-
8179. doi:10.1016/j.biortech.2008.03.066
[4] M. D. Poeet, “Biodiesel Crop Implementation in Hawaii,”
Hawaii Agricultural Research Center, Hawaii, 2006, pp.
5-10.
[5] L. J. Fuglie, “The Miracle Tree, the Multiple Attributes of
Moringa,” Church World Service, Dakar, 2001, p. 85.
[6] W. Siegfried, O. Viret, B. Huber and R. Wohlhauser,
“Dosage of Plant Protection Products Adapted to Leaf
Area Index in Viticulture,” Crop Protection, Vol. 26, No.
2, 2007, pp. 73-82. doi:10.1016/j.cropro.2006.04.002
[7] R. G. D. Steele and J. H. Torrie, “Principles and Proce-
dures of Statistics,” 2nd Edition, McGraw-Hill, New York,
1980.
[8] P. Villar-Salvador, L. Ocaña, J. Peñuelas and I. Carrasco,
“Effect of Water Stress Conditioning on the Water Rela-
tions, Root Growth Capacity, and the Nitrogen and Non-
structural Carbohydrate Concentration of Pinus halepen-
sis Mill. (Aleppo Pine) Seedlings,” Annals of Forest Sci-
ence, Vol. 56, No. 6, 1999, pp. 459-465.
doi :1 0. 10 51 /fo r est: 1 99 90 60 2
[9] M. J. Sánchez-Blanco, T. Ferrández, A. Navarro, S.
Bañon and J. J. Alarcón, “Effects of Irrigation and Air
Humidity Preconditioning on Water Relations, Growth
and Survival of Rosmarinus officinalis Plants during and
after Transplanting,” Journal of Plant Physiology, Vol.
161, No. 10, 2004, pp. 1133-1142.
doi:10.1016/j.jplph.2004.01.011
[10] S. Bañon, J. Ochoa, J. A. Franco, J. J. Alarcón and M. J.
Sánchez-Blanco, “Hardening of Oleander Seedlings by
Deficit Irrigation and Low Air Humidity,” Environmental
and Experimental Botany, Vol. 56, No. 1, 2006, pp. 36-
43. doi:10. 10 16 / j. en vexp bo t . 20 04.1 2 .0 04
[11] C. M. Menzel and B. F. Paxton, “The Effect of Tem-
perature on Growth and Dry Matter Production of Lychee
Seedlings,” Scientia Horticulturae, Vol. 26, No. 1, 1985,
pp. 17-23. doi:10.1016/0304-4238(85)90097-4
[12] T. Trochoulias and E. Lahav, “The Effect of Temperature
on Growth and Dry-Matter Production of Macadamia,”
Scientia Horticulturae, Vol. 19, No. 1-2, 1983, pp. 167-
176. doi:10.1016/0304-4238(83)90058-4
[13] A. P. George and R. J. Nissen, “The Effects of Day/Night
Temperatures on Growth and Dry Matter Production of
Custard Apple (Annona cherimola × Annona squamosa,)
Cultivar African Pride,” Scientia Horticulturae, Vol. 31,
No. 3-4, 1987, pp. 269-274.
do i:1 0. 10 16 / 03 04 - 42 38 (87)90 05 2- 5
[14] N. Utsunomiya, “Effect of Temperature on Shoot Growth,
Flowering and Fruit Growth of Purple Passionfruit (Pas-
siflora edulis Sims var. edulis),” Scientia Horticulturae,
Vol. 52, No. 1-2, 1992, pp. 63-68.
do i:1 0. 10 16 / 03 04 - 42 38 (92)90 00 8- Z
[15] A. W. Whiley, T. S. Rasmussen, J. B. Saranah and B. N.
Wolstenholme, “Effect of Temperature on Growth, Dry
Matter Production and Starch Accumulation in Ten Man-
go (Mangifera indica L.) Cultivars,” Journal of Horti-
cultural Science, Vol. 64, No. 6, 1989, pp. 753-766.
[16] E. Lahav and T. Trochoulias, “The Effect of Temperature
on Growth and Dry Matter Production of Avocado Plants,”
Australian Journal of Agricultural Research, Vol. 33, No.
3, 1982, pp. 549-558. doi:10.1071/AR9820549
[17] R. J. Downs and H. Hellermers, “Environment and the
Experimental Control of Plant Growth,” Academic Press
Inc., London, 1975.
[18] J. F. Morton, “The Horseradish Tree, Moringa pterigo-
sperma (Moringaceae). A Boon to Arid Lands?” Eco-
nomic Botany, Vol. 45, No. 3, 1991, pp. 318-333.
[19] M. H. Mughal, G. Ali, P. S. Srivastava and M. Iqbal,
“Improvement of Drumstick (Moringa pterygosperms
Gaertn.)—A Unique Source of Food and Medicine through
Tissue Culture,” Hamdard Medical, Vol. 42, No. 1, 1999,
pp. 37-42.
Copyright © 2011 SciRes. AJPS