Paper Menu >>
Journal Menu >>
 Applied Mathematics, 2011, 2, 1405-1416 doi:10.4236/am.2011.211199 Published Online November 2011 (http://www.SciRP.org/journal/am) Copyright © 2011 SciRes. AM Prey Predator Fishery Model with Stage Structure in Two Patchy Marine Aquatic Environment Mellachervu Naga Srinivas1, Mantripragada Ananata Satya Srinivas2, Kalyan Das1, Nurul Huda Gazi3 1School of Advanced Sciences, Department of Mathematics, VIT University, Vellore, India 2Department of Mathematics, Jawaharlal Nehru Technological University, Hyderabad, India 3Department of Mathematics, Aliah University, Kolkata, India E-mail: {mnsrinivaselr, massrinivas}@gmail.com, {kalyandas 70, nursha }@rediffmail.com Received July 4, 2011; revised August 2, 2011; accepted August 9, 2011 Abstract In this paper, we propose and analyze a mathematical model to study the dynamics of a fishery resource sys- tem with stage structure in an aquatic environment that consists of two zones namely unreserved zone (fish- ing permitted) and reserved zone (fishing is strictly prohibited). In this model we introduce a stage structure in which predators are split into two kinds as immature predators and mature predators. It is assumed that immature predators cannot catch the prey and their foods are given by their parents (mature predators). It is also assumed that the fishing of immature predators prohibited in the unreserved zone and predator species are not allowed to enter inside the reserved zone. The local and global stability analysis has been specified. Biological and Bionomical equilibriums of the system are derived. Mathematical formulation of the optimal harvesting policy is given and its solution is derived in the equilibrium case by using Pontryagin’s maximum principle. Keywords: Prey Predator, Stage Structure, Local and Global Stability, Bionomic Equilibrium, Optimal Harvesting, Pontryagin’s Maximum Principle 1. Introduction There are numerous studies on the effects of harvesting on population growth. In the context of Predator-prey interaction, some studies that treat the populations being harvested as a homogeneous resource include those of Brauer and Soudack [1,2], Dai and Tang [3], Myer- scough et al. [4], Chaudhuri [5] and Leung [6]. For a first look at the problem of harvesting from a bioeconomic or control theory point of view, see the works of Clark [7] and Levin et al. [8]. But they have not considered stage structure of species. Some of the stage structured models using time delay were considered by Aiello and Freed- man [9], Freedman and Gopalsammy [10], Rosen [11], Fisher and Goh [12], Cushing and Saleem [13], and some other authors. In general, stage structured models exhibit much more complicated dynamics than ordinary models. Bioeconomic modeling of the exploitation of biologi- cal resources such as fisheries and fore tries has gained importance in recent years. The techniques and issues associated with the bioeconomic exploitation of these resources have been discussed in detail by Clark [7,14]. Since most marine fisheries are essentially multi species in nature, exploitation of mixed species fisheries has started to draw attention from researchers. Although nu- merous models on single species fisheries have so far appeared in the fishery literature, no fully adequate stud- ies on multispecies fisheries appear to exist. It is very difficult to construct a realistic model of a multispecies community. Even if we succeed in formu- lating such a model, it is quite likely that the model may not be analytically tractable. Not every part of the catch is edible and harvesting harms some of the marine spe- cies which live on the other species in the sea. Thus the predator species are likely to become extinct with an in- discrete increase in the harvesting of prey species. There- fore, how best to harvest an ecologically or economically interdependent population in the sense of maximizing the present value of a stream of revenues from them, while maintaining ecological balance, is an important optimal control problem for fisheries. Clark [14] discussed an optimal equilibrium policy for  M. N. SRINIVAS ET AL. 1406 the combined harvesting of two ecologically independent species. Chaudhuri [5,15] formulated an optimal control problem for the combined harvesting of two competing species. Models on the combined harvesting of a two species prey predator fishery have been discussed by Chaudhuri and Saha Ray [5], Mesterton-Gibbons [13], Ragozin and Brown [16] etc. Most of the mathematical models on the harvesting of a multispecies fishery have so far assumed that the species are affected by harvesting only The prey-predator system is an important population model and has been studied by many authors [17-19]. It is assumed in the classical predator-prey model that each individual predator possesses the same ability to attack prey. Classical continuous population models such as logistic model and Volterra models overlook age struc- tures and space structures. Also these models overlooked the rate at which mature predators attack the prey and the reproductive rate are also ignored. In [20] a stage struc- tured model of one species growth consisting of imma- ture and mature individuals was analyzed. In [21], it was further assumed that the time from immaturity to matu- rity is itself state dependent. In recent years, the optimal management of renewable resources, which has a direct relationship to sustainable development, has been studied extensively by Clark [14], K. S. Chaudhuri [15], T. K. Kar, M. Swarnakamal [22] and W. Wang, L. Chen [23]. At present people are facing the problems due to shortage of resources. Extensive and unregulated harvesting of marine fishes can even lead to the extinction of several fish species. This problem can be addressed by arranging marine reserved zones, where fishing and other activities are prohibited. This marine reserve not only protects species inside the reserve area but also increase fish abundance in adjacent areas. The model of ecological system reflecting these problems has been given by T. K. Kar et al. [22] and Rui Zhang et al. [24]. Wendy-Wang et al. [25], considered prey-predator model with a stage structure in which predators are split in to immature predators and matured predators. They also assumed that the matured predators catch the prey and provide food for the immature predators. Rui Zhang et al. [24] considered a prey predator fishery model with prey dispersed in two patch environment, one is free zone for fishing and other is reserved zone where fishing is prohibited. 2. The Mathematical Model Here we consider a habitat where prey and predator spe- cies are living together. It is assumed that the habitat is divided into two zones, namely, reserved and unreserved zones. It is also assumed that predator species are not allowed to enter inside the reserved zone whereas the free mixing of prey species from reserved to unreserved zone and vice-versa is permissible. In the present paper we proposed a prey-predator model by combining the two features by [24,25], in which prey dispersed in a two patch environment and predator is not allowed to enter inside the reserved zone. Also a stage-structure is incorporated in which predators are split in to immature and mature predators. Here it is assumed that the prey migrates from unreserved zone to reserved zone and vice-versa. It also assumed that the fishing of immature predators is prohibited in the unre- served zone. This paper deals with the following prey- predator system 2 111 111 2112 2111 1 d d xrx rx xyxxqE tk x (2.1) 2 222 2211 22 2 d d xrx rxx x tk (2.2) 2 11122112 12 12 d d yxy xyy twyy wyy w (2.3) 22112 222 22 12 d d yxyyw dy qEy twyy . (2.4) Here 1()x t, 2()xt represents biomass densities of prey species in the unreserved and reserved areas respec- tively at a time “t”. 1, 2 represents biomass densities of immature and mature predators in unreserved area. 1, 2 represents intrinsic growth rates of prey species in unreserved and reserved areas respectively. 1, 2 represents carrying capacities of prey species in unreserved and reserved zones respectively. ()yt ()yt r k r k repre- sents capturing rate or capturing efficiency of the preda- tors. 1 , 2 represents migration rates from unre- served to reserved zones and vice-versa. 1, 2 repre- sents catch-ability coefficients of prey and matured predators respectively in unreserved zone. 1, 2 re- presents the efforts applied to harvest the prey and ma- tured predator species respectively in unreserved zone. 2 represents the death rate of mature predators in un- reserved zone. 1 q q E E d represents birth rate of predators. 2 represents conversion coefficient of immature predators to matured predators. We suppose that the attacking rate of mature predators at the prey i.e. the loss rate of prey is 12 x y . Since mature predators and immature predators may have distinct consumption rates to the resource, we as- sume that they consume the resource in the ratio , wher e measures the relative consumption ratio be- tween one immature predator and one mature predator, i.e., gives the allocation ratio of food between one immature predator and one mature predator. We assume :1w w :1w Copyright © 2011 SciRes. AM  M. N. SRINIVAS ET AL.1407 that all the weighted individuals share the quantity of food availability in equal parts. As a result, a fraction of resource consumed by mature predator is 2 12 12 y xy wy y and a fraction of resource consumed by immature predators is 1 12 12 wy xy wy y . If there is no migration of fish population from the re- served area to the unreserved area (i.e. 2 = 0) and 111 rq 1 E < 0, then 1 d d x t < 0. Similarly if there is no migration of fish population from the unreserved area to reserved area (i.e. 1 = 0) and 2 r2 < 0, then 2 d d x t < 0. Therefore we assume that 11 r1 q 1 E > 0, 2 r2 > 0 throughout our analysis. 3. Existence of Equilibria The steady state equations of (2.1)-(2.4) are 2 11 111 2112 2111 1 0 rx rx xyxxqEx k (3.1) 2 22 221122 2 0 rx rxx x k (3.2) 21 112 212 12 12 0 ywy xy xy wy ywy y (3.3) 1 21222 222 12 0 wy xyd yqE y wy y (3.4) The three possible equilibrium points are 1) 11 2 (In the absence of predators in un- reserved zone); ( ,,0,0)Gx x 2) 212 1 (In the absence of mature preda- tors in unreserved zone); (,, ,0)Gxx y 3) 31212 ,,,Gxxyy (The interior equilibrium). Case 1): : 11 2 ( ,,0,0)Gx x In this case, 1 x , 2 x are the positive solutions of (3.1) & (3.2). It may be noted that for 12 , x x to be positive we must have 211 11221 2 12 22 () rrqE r k k r (3.5) 221111 1 2 rrqE (3.6) 11 1111 1 rx rqE k . (3.7) Case 2): In the absence of mature predators in the un- reserved zone, there will not be any production of im- mature predators and hence this case coincides with Case 1). Case 3): Solving (3.1), (3.2), (3.3), & (3.4), we get 1122 12 1 22 x dqE (3.8) 2 22 22222 22 4 2 kr xr r rk 11 x (3.9) 2 11 111111 21 1 rx yrqEx xw k 1 22 x (3.10) 2 11 211111 2 11 1rx yrqEx xk 2 x . (3.11) For 1 y and 2 y both to be positive, we must have 2 11 11111 22 1 rx rqExx k . (3.12) 4. Qualitative Analysis of the Model 4.1. Local Stability Analysis Let us now suppose that system (2.1)-(2.4) has a unique equilibrium 31212 ,,,Gxxyy . 11 1213 14 21 2223 24 31 3233 34 41 4243 44 J JJJ J JJJ J J JJJ J JJJ (4.1) where 111122 111211 1 11 2rxrx x Jry qE kk 1 x 21 1 J ; 2 12 212 31 12 12 yyy Jwy ywy y w ; 212 41 12 yyw Jwyy ; 12 2 J ; 2222 11 22 22 222 x 32 2rxrx x Jrkk 0J; ; 42 0J ; 13 0J ; 230J ; 22 112 212 33 22 12 12 () ()() x ywx yw Jwy ywyy ; Copyright © 2011 SciRes. AM  M. N. SRINIVAS ET AL. 1408 2 212 43 2 12 () () x yw Jwyy ; 14 1 J x ; 24 0J; 22 1112 112211 34 22 12 1212 2() ()()() 2 2 () x yywx yx yw Jwy ywy ywy y ; 22 2112112 4422 2 22 12 12 () () () x yw xwyy JdqE wy ywy y . The characteristic equation of the Jacobian matrix of (2.1)-(2.4) at 31212 ,,,Gxxyy is 43 2 1234 0aa aa (4.2) where 11222 211 1 12 12 12 122122 20 rx rxxx akkx x xywyyy p 22 1212111222 2 12 1221 22 322 21122112 12 4 22 2122112112212 2222 221 12 22 21 2( ) 0 rr x xrxrx akkkxk x xyyw xyyw p p rxxywxywrxywxyw kpxp kpp yyy 22 121211 1222 3 12 12 12 2 122112 12 22 22 21121222 11 12 2 22 0 rr x xrxrx akk kxxk xy wxyyw pp xyywxy wrxx pk p x 32 32 211222 11 412 3 22 0 xyywrx x akx p . Now, 11 222211 1 12 12 12 1221 22 2 12 2 + rx rxxx akkx x xyw yyy p xy w AB p ; 11222 211 12 12 =rxrxxx Akkx x ; 122122 Byy y 22 121211 1222 2 12 1221 rr x xrxrx a K KKxKx 22 121211 1222 2 12 1221 22 2122112112222 2222 221 12 22 21 22 322 21122112 12 4 12 12 22 21 2 22 32 2112 2( ) rr x xrxrx aKKKxK x rxxywx ywrx ywxyw KpxpKpp yyy xyyw xyyw p p xyw C Ayyy p xyyw p 2 2112 12 42( )xy yw p where 22 12 12111222 1212 21 rr x xrxrx C kk kxk x 22 12 12111222 3 12 12 12 2 122 112 12 22 22 2 112122211 12 2 22 rr x xrxrx akk kxxk xy wxyyw pp x yywxy wrxx pk p x 323 2 211222 11 412 3 22 0 xyywrx x akx p . Again, 212 12 32 22 32 2112 12 4 2 2112 11 22 11 22 22 2 121 2 24 3343232 222 21122 112 12 63 () 2( ) 2( ). xyw aaaA CAB p xyyw Ap xyywrxx pkx xywx yw CB AB pp x yy wxyyw B pp Therefore, where Copyright © 2011 SciRes. AM  M. N. SRINIVAS ET AL.1409 32 22 2221121 12 123 33 22 54 54 2 2112 12 7 43 33 2 211222 11 5 22 43 43 22 2112 12 12 52 () () () . rxxx yyw aaa aAB kx p xyyw AB p xyywrxx AB kx p xyyw xyw A BAC B pp 32 32 2112 12 3 2 22112112 22 22 22 22 12 4 3343 2112 12 6 254264 2 2112 12 7 24 2 () + 2( ) 2( ) xyyw AC p rx xxyyw AC kx p xyw AC B p xyyw AC B p xyyw Ap x A 32 43 11 22211 12 5 22 32 22 211211 22 3 11 2432 43 21121122 12 4 11 2422 22 21 1211222211 2 11 22 2( ) () yywrx x kx p xyyw rxx CB kx p xyywrx x kx p xyywrxxrx x kx kx p 33 33 3 12 6 44 54 2 2112 12 8 2652 75 2 2112 12 9 254254 2112 2211 12 7 22 2432 33364 2112211 5 2( ) 2( ) 2( ) xyw AC B p xyyw CB p xyyw B p xyywrx x Bkx p xyyw xy CB p 354 2 12 5 3533 33 21122211 4 22 () . yw p xyywrxx kx p (4.3) Again, 22 22 22 2 12 12 14 42 32 32 221121 12 12 3 22 22 22 11 141 2 22 32 3254 54 2 21122112 37 22 11 12 22 2. () () . ( xywxyw aa AB AB pp rxxxyy w kx p rx x aaA kx xyyw xyyw B pp rx x kx 43 43 2112 5 22 11 12 22 )2 . () . x yy w Ap rx x Bkx (4.4) By combining first four terms of the right hand side of (4.3) with the first three terms of right hand side of (4.4), it can easily be established that 0 2 12331 4 ()aaaaaa . Since > 0, it is clear that 0 4 a 22 4123 143 ()aaaa aa a . Hence 41212 ,,,Gxxyy y Routh-Hu is locally asymptotically sta- ble. So brwitz criteria, it follows that all ei- gen values of (3.2) have negative real parts if andonly if and 0 13 4 0,0, >0,aa a 2 312 314 ()aaaaaa 22 4123 14 3 ()aaaa aa a . Hence, 31212 ,,,Gxxyy us the four populatio ns we have obtained be proved numeric stem for a set of pa es also satisfy the con ilibrium point is Locally asymptotically stable. Thns remain stable under the conditio in the above study. The fact can also ally. We have solved the above syrameter values. The parame- ter valudition for existence of the interior equ 31212 ,,,Gxxyy ain. So we use . The real data is very difficult to obta set of the hypothetical parameter values as follows: r1 = 1.6; r2 = 1.2; k1 = 270; k2 = 250; σ1 = 0.4; σ2 = 0.4; q1 = 3.0; q2 = 3.2; E1 = 0.02; E2 = 0.082; α1 = 1.9; α2 = 2.1; β = 0.06; w = 0.7; d2 = 0.22. The time series of the populations are shown in the Figure 1. The Figure 1 shows that the populations are finite for long time and the system is stable. 4.2. Global Stability Analysis Theorem I. The Equilibrium point is globally asymptotically stable. Proof: let us consider the following Lyapunov fun 11 2 ( ,,0,0)Gx x ction Copyright © 2011 SciRes. AM  M. N. SRINIVAS ET AL. Copyright © 2011 SciRes. AM 1410 050100 0 10 20 30 40 Time t P rey species i n pat ch I 050 100 0 50 100 150 Time t Prey speci es i n pat ch II 050100 10 20 30 40 50 60 Time t I m m at ure predators i n pat ch I 050 100 0 10 20 30 Time t Mat ured predators i n pat ch I Figure 1. Stable time series evolution of the prey and pre- dator populations of the model system (2.1)-(2.4). 12 121 111222 12 (, )lnln xx x xxxxlxxx xx V Differentiating V w.r.to “t” we get 11 122 2 1 12 dd d dd d x xx xxx Vl txt x . t Choosing 22 1 11 x lx , after a little algebraic manipulation, we get, 22 1222 112 2 111 2 2 2 12 12 112 d d 0. rxr V x xx tk xk xx xx xxx x Therefore is globally asymptotically 11 2 ( ,,0,0)Gx x stable. Theorem II. The equilibrium point 31 (,, ,)Gxxyy 2 1 2 is globally asymptotically stle. Proof: let us consider the following Lyapunov function ab 121 12121111222211132 22 121 1112 22111 12 1 2 2 (,, ,)lnlnlnln dd d d xxy y Vxxyyxxxlxxxlyyyly yy xxy y xxxx xxyyy Vll t 2 1 dd dtx xty 222 3 2 22 11 2 222 22 22 1 21 1122132221222 12 12 d () ()() yy y l tyt r xx x x rkx yy wy ly yxyxwlyyxdqE ywyy wyywyy 11 22 11 12 1111 11 dd d+ d rx x VxxryqE l tkx 1 112 1 d d Vx t 122 1112 2 111 11222 21 1322 1211 () rx x xx x y kxx xyy y ly ylyy wy yyy Choosing 2 11 2122 221 2 22 11 11 2 12 12 + . ()() r xx ylxx xx k xx xy xy wwy ywyy 22 1 11 x lx ; 2 1 1 l ; 12 3 21 wy y lwy 221 1 1 11 2112 11 22 22 11221 d d 1 xx r Vxx xxxx tk xx xx x xxxx xxxy 2 222 11 222 2 211 1 12 112112 11221 1112 11 11 11 22 2112 12 12 () () ()() rx xxyyx x kx xy yyyyyy ywyy x yx yx y wy ywyywy ywy y 1 2 2 wyy wy 11 12 2 22 2 1 2 2211221 12 11 1211112 () < 0. () xy wyy x x xxxxxyyyy kx xxxyywyy 2 122 2 11 12 d d rrx Vxx tk  M. N. SRINIVAS ET AL.1411 Therefore 31 2 1 2 (,,,)Gxxyy is globally asymptoti- cally stable. 5. Qualitative Bionomic Phenomena dy of the odels. Let r prey species, be the fishing cost per unit effort for matured predpecies, be the price per unit biomass of the prbe thce per unit bio- mass of the predator (m). Thnet revenue or conomic rent at any tiby where re- Net The bionomic equilibrium It is the stu dynamics of living resources using economic mbe the fishing cost per unit effort fo 1 c 2 c ator s ey, ature me Rp dator 22 1 p e pri erefore 2 ()c 2 R 2 p d given e12 RR R 2 E; here represents 11111 ()RpqxcE, presents Net Revenue for revenue for mature 122 qy the prey; d prespecies. 1 R 1212 12 () ,() ,() ,(),,xx yyE E is given by the following equations 2 11 111 2112 2111 1 0 rx rx xyxxqEx k (5.1) 2 22 2211 22 2 0 rx rxx x k (5.2) 2 112 2112 12 12 0 xy xyyw wy ywy y (5.3) 2112 222 22 12 0 xyywdy qEy wyy (5.4) (5.5) In order to determine the bionomic equilibrium we come across the following cases. Case I. If for the matured predator, fishing cost is greater than the revenue 2 ), then fishing of matured predator is not fea fishing of prey population remains operation1 ). Thus, when an1 2 RE y. 1111 12222 0pqxcpqcE (222 cpqy sible. Hence al (11 cp d cpq 1 qx x 20E111 we have 1 1 11 c xpq , 22 22222 2 11 4 () 2 krc xrr rp . 1 , 112 qk Since 1 c11111 pqxpqk 21 ,yE will be any point on the line 1 12111 111 2112211 22 22 21 112 1 4. 2 yqErpqk pqkr c rr rc pqk c Case II. If 1 i.e., the cost is greater than the revenue in th, then the prey fishing will be closed (i.e. ly matured predator fiing re- mains op2 111 cpqx e prey fishing 1 E= 0). On al (i.e. sh eration 222 cpqy ) 2 2 22 pq Substitu c y. te this value in (5.1), we get 21 211 1 ( )() cx xx pq 1 11 () 1x rx 222 1 k . Here 1 x is the positive solution of 32 11213140axaxaxa (5.6) where 2 12 122 122 0 rr akk ; 12 2 211 2 22 12 2 2rr c ar pq kk ; 2 221 311 2 2 221 2 22 rcr ar r pq k k 2 ; 2 42211 22 1c arrpq 1 2 . 2 11 22 c rpq < 0 (or) 2 11 22 c rpq Now if > 0, then 2 2212 11 2 2212 22 ()rcrr rpq k k 2 2 22111 22 () c rrpq 2 . and Then Equation (5.6) has a unique positive solution 11 x x 2 x to be positive, we must have For 21 11 11 2211 ck k xk pqr r . Substitute the value of 1 x in Equation (5.4), we get 21 1 2 1xyw E qwy y 2 212 d 20,E Provided 121 xd 2 2 where 1 12 21 y y w . Copyright © 2011 SciRes. AM  M. N. SRINIVAS ET AL. 1412 Case III. If 2 , then the co greater than re and the fishery will be closed. Case IV. If 2 , th nues for both th fishery will be in operation. In this case 111 cpqx, venues for 1222 cpqy both the species st is whole 111 cpqx, he species b 1222 cpqy eing positive, t en the reve- en the whole 1 1 11 c xpq (5.7) 2 2 22 c ypq . (5.8) Substitute (4.7) and (4.8) in (4.1), (4.2), (4.4) we get 2 k 211 22222 2 4rc 112 () 2 xrr r pqk (5.9) 11 21221 1 11 () 1 rc cxp Eqpqkqpq c (5.10) 1 1 2 11 q 1 2 1 q 211 22 212 () 1xyw Ed qwyy (5.11) if 10E 1122121 11111221 () 1 rc xpc pqkqcqpqq 1 1 q (5.12) if 20E 121 2 1211 cd pq . (5.13) The Non-trivial Bionomic equilibrium point 12 212 () ,() ,() ,() ,xx yEE exists if (5.12d (5.13) hold. in this section we employ the Pontryagin’s maximum principle tbtain a path of op- timal harvesting policy so that if the fish populations inre kept along this path, then the regulatory agency is assured to achieve its objective. We consider the following present value J of continuous time-stream ) an 6. Optimal Harvesting Policy In this section we study optimal harvesting policy of the system (2.1)-(2.4). Also o o side and outside the reserve zones, a a 1212 1 2 ,,,,,,e 0 d t J Px xyyE Et t (6.1) where is the net revenue given by (6.2) and P 1212 1 2 11111222 22 2 ,,,,,,PxxyyEE t pqxEcEp qy Ec E 1 denotes the instantaneous annual rate of dis- count, the aim of this section is to maximize J sub- jected to the state Equations (2.1). Firstly we construct the following Hamiltonian func- tion )-(2.4 111112 2222 1 E y 1 1111 2 11 211 1 2 2 1 2 21 3112212 12 12 1 421222 22 12 ee 1 1 tt HpqxcE pqyc x rxx yxxqEx k x k yw xy xy wyywy wy xydyqy wyy 2 22122 rxxx 2 E y w (6.3) here 1234 ,,, are additional unknown functions are the control vari- called the adjoint variables, 12 ,EE ables satisfying the constraints 11 max 0EE; 22 max 0EE , and 11111 ()e t tpqxc , 111 qx 222 22 ()e t tpqyc are called the switch- ing f 2 42 qy unctions. We aim to find an optimal equilibrium 1212 1 2 ,,,,,xxyy EE Hamiltonian to mize maxi H . Since Hamiltonian H al ontro is linear in the control vari- ables control can xtreme con- trols gular cls, thus we have ax 1 ,EE or the sin 2 , the optimbe e 1 (EE 1m ) () 0t, when 1 i.e., when 1 11 11 etc ; ()tp qx 10E , when 1() 0t i.e., when 1 11 11 etp ; () c tqx 22max ()EE , when 2()t0 i.e., when 2 42 22 ()etc tp qy ; 20E , when 2() 0t 2 42 22 ()etc tp qy i.e., when ; when ()0t 1 , 1 ()etc tp ; 11 11 qx or 1 0 H E 6.4) ( Copyright © 2011 SciRes. AM  M. N. SRINIVAS ET AL.1413 when 2() 0t , 2 42 22 ()etc tp qy ; or 2 0 H E . In this case, the optimal control is called the singular control, and (6.4), (6.5) are the necessary conditions for the maximization of Hamiltonian (6.5) H . By Pontrayagin’s maximal principle, the adjoi tions are nt equa- 1 1 δt11 11 1112111 σyqE 1 2 212 4 12 d d 2 r x epqE λr () H tx k ywyy wyy wyy 122 12 21 3 12 12 λσ wy ywyy (6.6) 222 122 22 22 d2 d rx Hr tx k (6.7) 3 1 2 112212 2 3 12 d d ( H ty 22 12 21 22 42 )() () 12 x yw xywy wy (6.8) ywy y xywy wy y 4 2 1 1 22 211 4222 2 12 d . () t ty 22 21 1 222 11 1222 e() 2 pqE x xyywy 322 12 12 () () dH x wy (6.9) wy ywyy xw ydq E wyy Considering the interior equilibrium 31212 ,,,Gxxyy as and Equations (6.4), (6.7), can be written 2 212 d() e d t AA t where 22 11 1 22 rx x Akx ; 1 21 11 c Ap qx 2 . We can calculate that 2 2 1 et A A . (6.10) larly, by conSimisidering the interior equilibrium 31212 ,,,Gxxyy and Equati.5), (6.8), can be wri- ons (6 tten as 3 det 3 4 d3 A A t where 212 3 12 ; wx y Awyy 2 2212 42 2 cwxy Ap qy wy y 22 12 clu that . We can conde 4 3 3 et A A . (6.11) interior equilibriumSimilarly, by considering the ,y and 312 ,,Gxxy Equations (6.10), (6.11), (6.6) can as 12 be written 1 15 6 de d t A A t where 1 5 rx Akx 122 11 x ; 24 AA 611 1 11221 13 2212 221 2 . A 2 pqEy wy AA cwyy qyy y conclude that pw We can 6 1 5 et A A . (6.12) Similarly by considering the interior equilibrium 3 G12 ,,,xxyy and Equations (6.10), (6.11), (6.6) can be written as 12 4 47 d d8 et A A t where 2 y2 21 1 7222 2 12 xw dqE wyy ; A 64212 . 822 2 1 A 5312 A xwy Apq E x AAwyy We can calculate that 8 4e At 7 A It is obvi . (6.13) ously that 1234 (), (), (), ()tttt are bound- ed as Substitute (6.12) in (6.4), we obtain a singular path t. 6 1 1 11 5 A c pqx A . (6.14) Substitute (6.13) in (6.5), we obtain a singular path Copyright © 2011 SciRes. AM  M. N. SRINIVAS ET AL. 1414 8 2 2 22 7 A c pqy A . (6.15) Using 2 221 22222 22 4 2 krx xr r rk 1 2 11 2111112 11 1rx yrqEx xk , 2 x 12345678 ,,,,,,, A AAAAAAA can be written as 1 2 2211 122 22 2 211 1/2 2211 22 22 2 2 4 11 () 22 2 4 ()() rx Arr k rx rx kr rk 1 21 11 c Ap qx 2 12 11 31 12 11 wxc Ap qx 2 2 42 2 21 11111 22 11 c Ap qrx rqEx x xk 1 1/2 2 112 2211 52222 112 2 4 ()() 2 rxkrx Arr kxr k 1 12 11 6111 22 11 pqx Ap q Erx x 22 1 1 22 12 11 1 11 22 1 ) c kx cx w y rx Ex x k 2 2 2 +p 22 2 (qr q 2 2 121 x 7222 2 12 A dqE 2 A6421 2 2 1 5312 + 82 A xw Ap xq EAA . (6.14) and (6.15) can be written as Thus 6 1 11 11 5 A c Fxp qx A and 8 2 22 22 7 A c Gyp qy A . There exists a unique po11 () x x of sitive root 10Fx in the interval 11 0 x k If the following hold F (0) <0, F) > 0, (1 k 10x for F 10x, and 22 ()yy Similarly There exists a unique positive root of 20Gy in the interval 22 0yk If the ) < 0, G (2 k) > 0, fol- lowing hold G (0 20Gy for 20y. For 11 () x x , 22 ()yy , we get 2 22 22222 2 4 () 2 kr xr r rk 11 x 21 2 1 () () wy y ; 2 11 1 11111122 1 () () () rx yrqEx x k 21 xw 1 11 1 1112112 2 () ()()()( ) xxyxx 1 1 () () () 1 Eqx rx k 21 12 2 ()Ed 22 221 2 ()()( () ()() () xyyw y qywy y ) 1 i.e., 21 1 22 212 ()( ) 1 () ()( ) xyw Ed qwyy . Hence once the optimal equilibrium, 1212 (),( ),(),( )xx yy is determined, the optimal harvesting effort 1 ()E and 2 ()E (6.11), ( can be determined. From (6.3), (610), found that .4), (6.6.12) and (6.13), we () it where = 1, 2, 3 timri. Hence they rem 7. We have considered a system of tw fe ui- lib is stable. We have shown the stability r and also numerically. We can also considered a delayed mm to take into account of the gestational de- lay of the matured predator population. It is natural that the consumption on the prey by the predator needsome time to contribute to the biomass of the predator. So we use delay differential Equation (7.1) to study sue- nomena. i um , 4, do not vary with e in optimal equilibain bounded as t . Computer Simulation Discussion o populations at dif- rent stage structure. The stability of the interior eq rium point is studied and it is shown that the system esults analytically odel syste s ch ph 2 111 111 2112 21 yxxq 11 1 d d xrx rx xEx tk Copyright © 2011 SciRes. AM 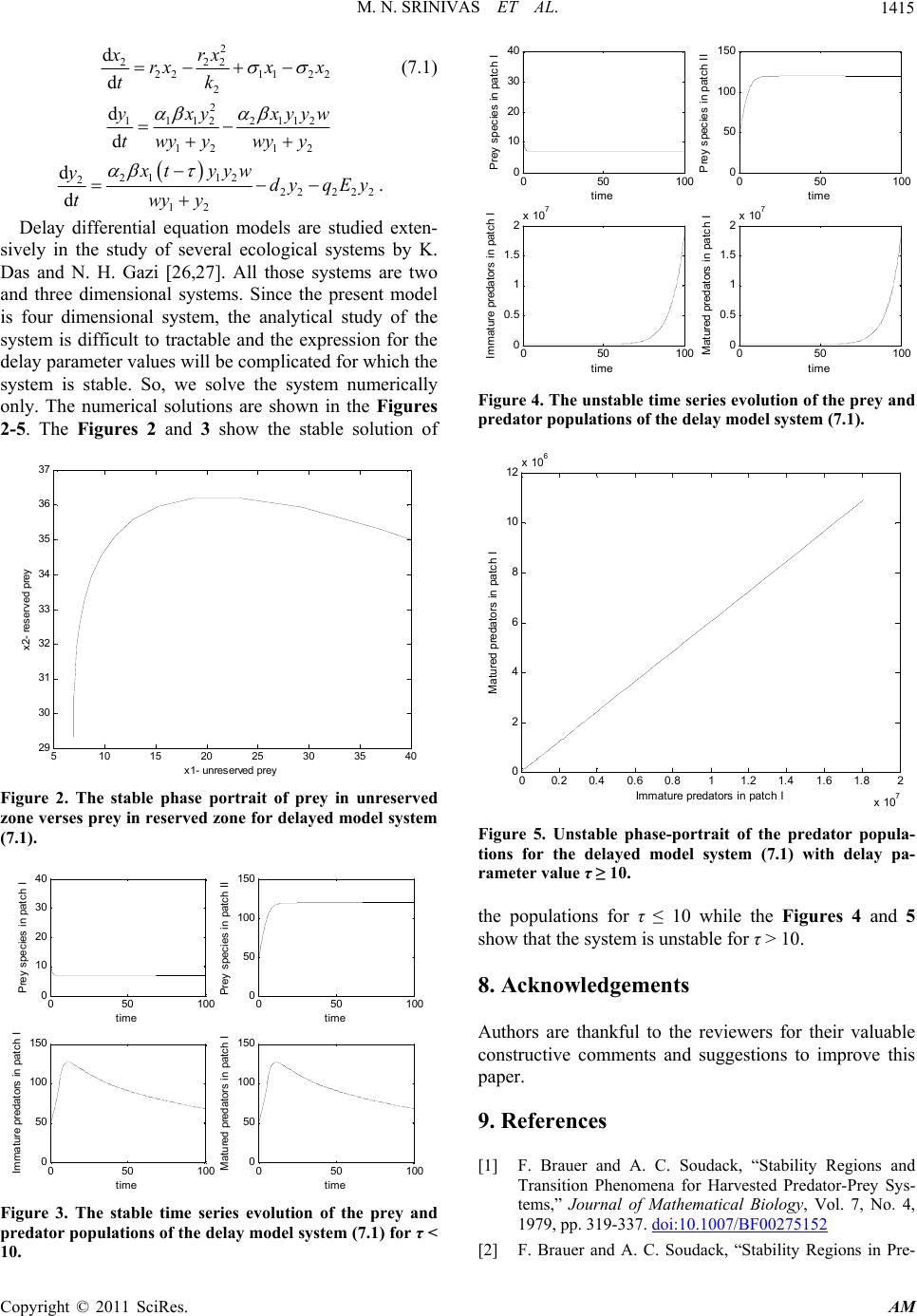 M. N. SRINIVAS ET AL.1415 2 222 221122 2 d d xrx rxx x tk (7.1) 2 11 122 112 12 12 d d yxy xyy twyy wyy w 21 12 2 222 22 12 d d xt yyw ydy qEy twyy . Delay differential equation models are studied exten- sively in the study of several ecological systems by K. Das and N. H. Gazi [26,27]. All those systems are two and three dimensional systems. Since the present model is four dimensional system, the analytical study of the system is difficult to tractable and the expression for the delay parameter values will be complicated for which the system is stable. So, we solve the system numerically only. The numerical solutions are shown in the Figures 2-5. The Figures 2 and 3 show the stable solution of 510 15 20 25 30 35 40 29 30 31 32 33 34 35 36 37 x 1- unreserved prey x 2- res erved prey Figure 2. The stable phase portrait of prey in unreserved zone verses prey in reserved zone for delayed model stem (7.1). sy 050 100 0 10 20 30 40 time s pecies in patchPrey I 050100 0 50 100 150 time Prey species in patch II 050 100 0 50 100 150 tim I m patch I e mature predat ors in 050100 0 50 100 150 time . The stable time series evolution of the prey and populations of the delay model system (7.1) for τ < 10. Matured predat ors in patch I Figure 3 predator 050 100 0 10 20 30 40 time Prey speci es in patch I 050 100 0 50 100 150 time Prey speci es in patch II 050 100 0 0.5 1 1.5 2x 10 7 time I m m ature predat ors i n patch I 050 100 0 0. 5 1 1. 5 2x 10 7 time Matured predat ors i n patch I Figure 4. The unstable time series evolution of the pr and predator populations of the delay model system (7.1). ey 00.2 0.40.6 0.8 11.2 1.41.6 1.8 2 x 10 7 0 2 4 6 8 10 12 x 10 6 Immat ure predators in pat ch I Figure 5. Unstable phase-portrait of the predator popula- tions for the delayed model system (7.1) with delay pa- rameter value τ ≥ 10. the populations for τ ≤ 10 while the Figures 4 and 5 show that the system is unstable for τ > 10. 8. Ackn Mat ured predat ors in owledgements Authors are thankful to the reviewers for their vaable constructive comments and suggestions to improve this paper. 9. References [1] F. Brauer and A. C. Soudack, “Stability Regions and Transition Phenomena for Harvested Predator-Prey Sys- tems,” Journal of Mathematical Biology, Vol. 4, patch I lu 7, No. 1979, pp. 319-337. doi:10.1007/BF00275152 F. Brauer and A. C. Soudack, “Stability Regions in Pre-[2] Copyright © 2011 SciRes. AM  M. N. SRINIVAS ET AL. Copyright © 2011 SciRes. AM 1416 dator-Prey Systems with Constant-Rate Prey Harvesting,” Journal of Mathematical Biology, Vol. 8, No. 1, 1979, pp. 55-71. doi:10.1007/BF00280586 [3] G. Dai and M. Tang, “Coexistence Region and Global Dynamics of a Harvested Predator-Prey System,” SIAM: SIAM Journal on Applied Mathematics, Vol. 58, No. 1, 1998, pp.193-210. doi:10.1137/S0036139994275799 [4] M. R. Myerscough, B. E. Gray, W. L. Hograth and J. Norbury, “An Analysis of an Ordinary Differential Equa- tion Model for a Two-Species Predator-Prey System with Harvesting and Stocking,” Journal of Mathematical Bi- ology, Vol. 30, 1992, pp. 389-401. [5] K. S. Chaudhuri and S. S. Ray, “On the Combined Har- vesting of a Prey-Predator System,” Journal of Biological Systems, Vol. 4, No. 3, 1996, pp. 373-389. doi:10.1142/S0218339096000259 [6] A. W. Leung, “Optimal Harvesting Co-Efficient Control of Steady State Prey-Predator Diffusive Volterra-Lotka Systems,” Applied Mathematics & Optimization, Vol. 31, No. 2, 1995, pp 219-241. doi:10.1007/BF01182789 [7] C. W. Clark, “Mathemalical Bioeconomics: The Optimal Management of Renewable Resources,” John Wiley and Sons, New York, 1979. [8] S. A. Levin, T. G. Hallam and J. L. Gross, “Applied Mathematical Ecology,” Springer-Verlag, Berlin, 1989 /0025-5564(90)90019-U . [9] W. G. Aiello and H. I. Freedman, “A Time Delay Model of Single Species Growth with Stage Structure,” Mathe- matical Biosciences, Vol. 101, No. 2, 1990, pp. 139-153. doi:10.1016 H. I. Freedman and K. Gopalsammy, “Global Stability in Time-Delayed Single Species Dynamics,” Bulletin of Mathematical Biology, Vol. 48, No. 5-6, 1986, pp. 485- 492. [11] G. Rosen, “Time Delays Produced by Essential Nonlin- earity in Population Growth Models,” Bulletin of Mathe- matical Biology, Vol. 49, No. 2, 1987, pp. 253-255. [12] M. E. Fisher and B. S. Goh, “Stability Results for De- layed Recruitment Models in Population Dynamics,” Journal of Mathematical Biology, Vol. 19, No. 1, 1984, pp. 147-156. doi:10.1007/BF00275937 [10] [13] M. Mesterton-Gibbons, “On the Optimal Policy for the Combined Harvesting of Predator and Prey,” Natural Resource Modeling, Vol. 3, 1988, pp. 63-90. [14] C. W. Clark, “Mathematical Bioeconomics: The Optimal Management of Renewable Resources,” Wiley, New of a Multi Species Fishery,” Ecological Modelling, Vol. 32, No. 4, 1986, pp. 267-279. doi:10.1016/0304-3800(86)90091-8 York, 1976. [15] K. S. Chaudhuri, “A Bio Economic Model of Harvesting [16] D. L. Ragozin and G. Brown, “Harvest Policies and Non Market Valuation in a Predator Prey System,” Journal of Environmental Economics and Management, Vol. 12, No. 2, 1985, pp. 155-168. doi:10.1016/0095-0696(85)90025-7 [17] A. Hastings, “Global Stability of Two Species Systems,” Journal of Mathematical Biology, Vol. 5, 1978, pp.399- 403. [18] X.-Z. He, “Stability and Delays in a Predator-Prey Sys- tem,” Journal of Mathematical Analysis and Applications, Vol. 198, No. 2, 1996, pp. 355-370. doi:10.1006/jmaa.1996.0087 [19] B. S. Goh, “Global Stability in Two Species Interac- tions,” Journal of Mathematical Biology, Vol. 3, No. 3-4, 1976, pp. 313-318. doi:10.1007/BF00275063 [20] W. G. Aiello and H. I. Freedman, “A Time Delay Mode doi:10.1016/0025-5564(90)90019-U l of Single Species Growth with Stage Structure,” Mathe- matical Biosciences, Vol. 101, No. 2, pp. 139-153. [21] W. G. Aiello, H. I. Freedman and J. Wu, “Analysis of a Model Representing Stage Structured Population Growth with State-Dependent Time Delay,” SIAM: SIAM Journal on Applied Mathematics, Vol. 52, No. 3, 1992, pp. 855- 869. [22] T. K. Kar and M. Swarnakamal, “Influence of Prey Re- serve in a Prey-Predator Fishery,” Non-Linear Analysis, Vol. 65, No. 9, 2006, pp.1725-1735. [23] W. Wang and L. Chen, “Optimal Harvesting Policy for Single Population with Periodic Coefficients,” Mathe- matical Biosciences, Vol. 152, No. 2, 1998, pp. 165-177. doi:10.1016/S0025-5564(98)10024-X [24] R. Zhang, J. F. Sun and H. X. Yang, “Analysis of a Prey- Predator Fishery Model with Prey Reserve,” Applied Ma- thematical Sciences, Vol. 50, No. 1, 2007, pp. 2481- 2492. [25] W. D. Wang, Y. Takeeuchi,Y. Saito and S. Nakaoka, “Prey-Predator System with Parental Care for Predators,” Journal of Theoritical Biology, Vol. 241, No. 3, 2005, pp. 451-458. doi:10.1016/j.jtbi.2005.12.008 [26] K. Das, N. H. Gazi, “Structural Stability Analysis of an 010, pp. 2191- ontrol of Parameters of a De- Algal Bloom Mathematical Model in Trophic Interac- tion,” International Journal of Non-linear Ananlysis: Real World Applications, Vol. 11, No. 4, 2 2206. 27] N. H. Gazi and K. Das, “C[ layed-Diffusive Autotroph-Herbivore System,” Interna- tional Journal of Biological System, Vol. 18, No. 2, 2010, pp. 509-529. |