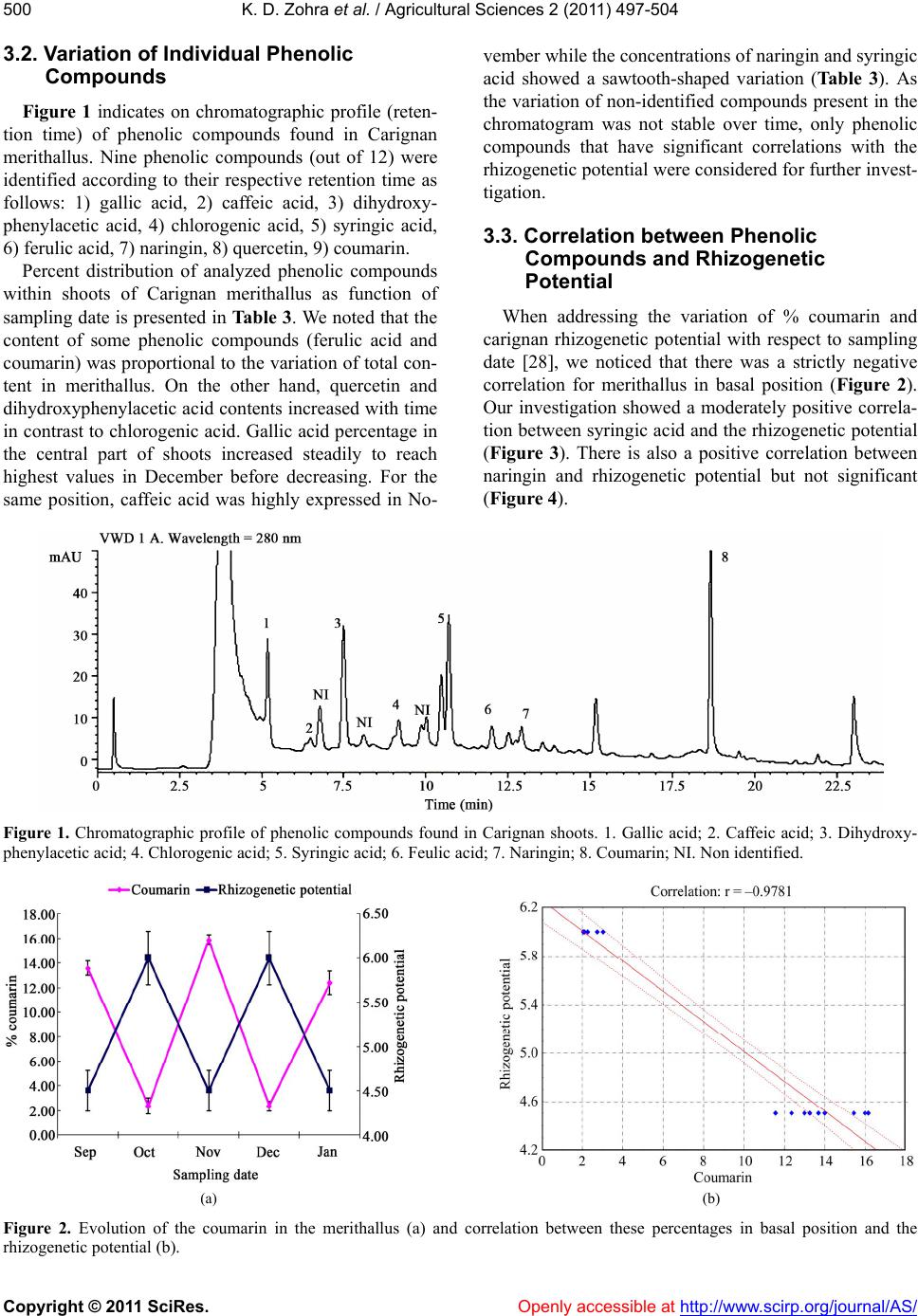
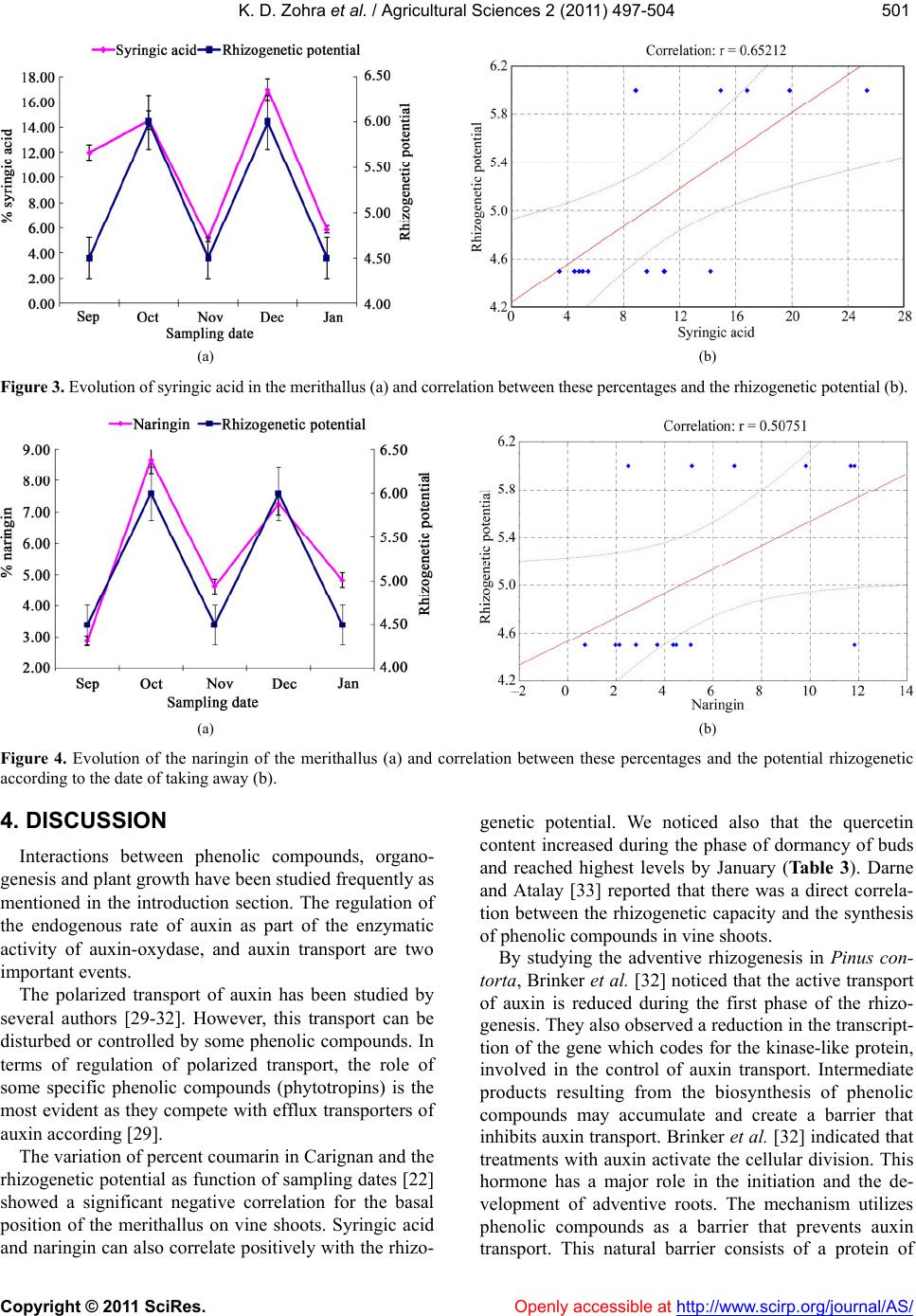
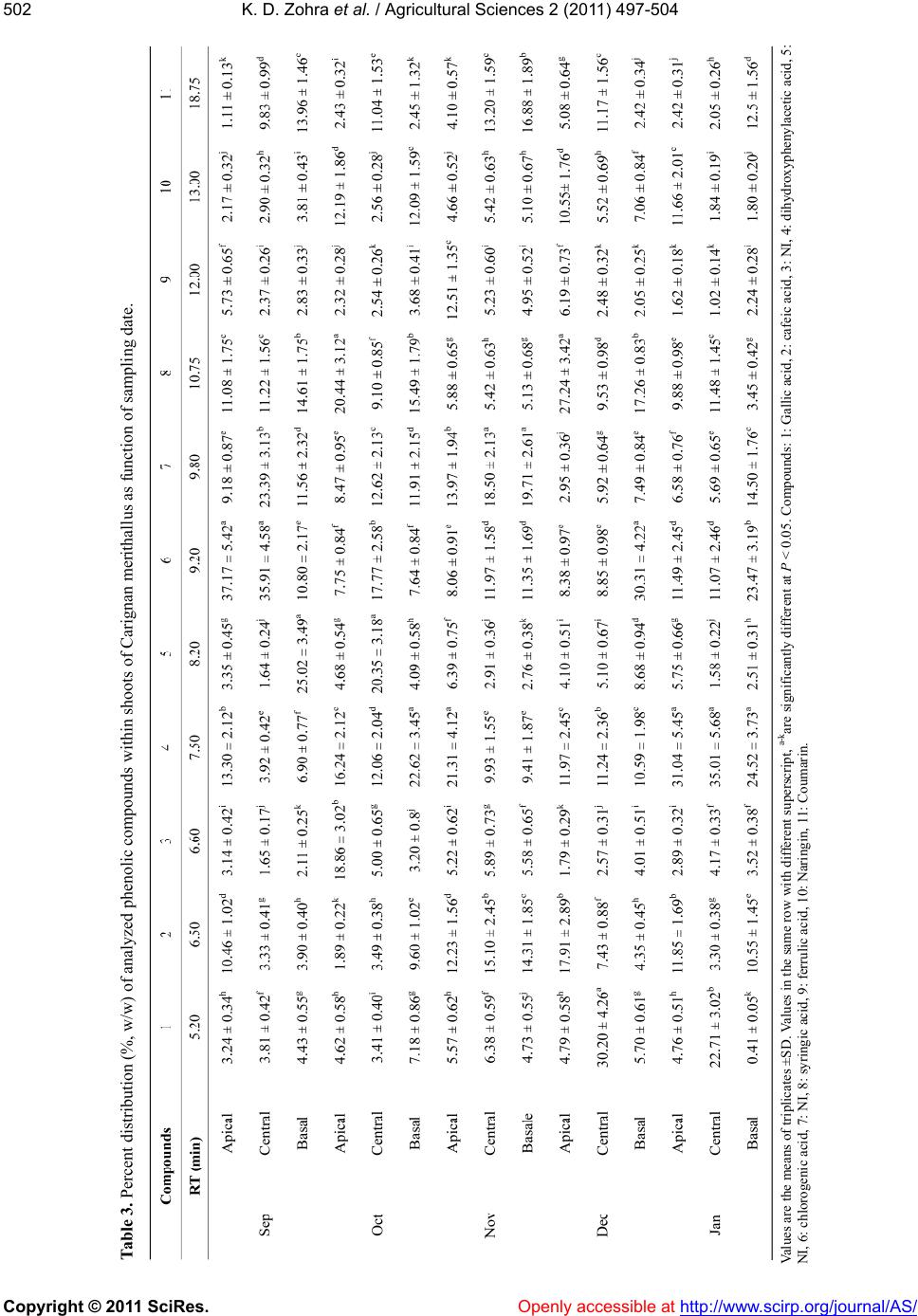
K. D. Zohra et al. / Agricultural Sciences 2 (2011) 497-504
Copyright © 2011 SciRes. Openly accessible at http://www.scirp.org/journal/AS/
503503
having a flavonolic origin [30,32] noticed that the tran-
scription of the gene coding for this enzyme is affected
during the pha se of radial primordial formation.
5. CONCLUSIONS
The evolution of phenolic compounds was studied on
Carignan merithallus during the phases of bud dormancy
and end of dormancy. Our outcomes indicate that rhizo-
genesis coincides with quercetin synthesis. This phenolic
compound belongs to the class of phytotropins according
to Brunn et al. [29]. We observed also a negative corre-
lation between coumarin and rhizogenetic potential of
Carignan vine. In contrast, positive correlations were
found with naringin and syringic acid.
REFERENCES
[1] Ho, C.T., Lee, C.Y. and Houng, M.T. (1992) Phenolic
compounds in food and their effects on health. I. Analy-
sis, occurrence, and chemistry. ACS Symposium Series,
506.
[2] Oleszek, W., Amiot, M.J. and Aubert, S.Y. (1994) Identi-
fication of some phenolics in pear fruit. Journal of Agri-
cultural and Food Chemistry, 42, 1261-1265.
doi:10.1021/jf00042a002
[3] Amiot, M.J., Aubert, S. and Nicolas, J. (1993) Phenolic
composition and browning susceptibility of various apple
and pear cultivars at maturity and postharvest. Acta Hor-
ticultureae, 343, 67-69.
[4] Blankenship, S.M. and Richardson, D.G. (1985) Ch an ges i n
phenolic acids and external ethylene during long-term
cold storage of pears. Journal of American Society of
Horticulturea Science, 110, 336-339.
[5] Challice, J.C. and A.H. Williams, (1968) Phenolic com-
pounds of genus Pyrus-II. A chemotaxonomic survey.
Phytochemistry, 7, 1781.
doi:10.1016/S0031-9422(00)86651-0
[6] Challice, J.C. and Williams, A.H. (1972) Phenolic com-
pounds of genus Pyrus. Phytochemistry, 11, 37-44.
doi:10.1016/S0031-9422(00)89964-1
[7] Risch, B. and Hermann, K. (1989) Contents of hydroxy-
cinnamic acid derivates and catechins in pome and stone
fruits. Zeitschrift Fuer Lebensmittel-Untersuchung Und
-forschung, 186, 225-230. doi:10.1007/BF01043317
[8] Ayaz, F.A., Kadioglu, A. and Reunanen, M. (1997)
Changes in phenolic acid contents of Diospyros lotus L.
during fruit development. Journal of Agricultural and
Food Chemistry, 45, 2539-2541. doi:10.1021/jf960741c
[9] Babic, I., Amiot, M.J., Nguyen-The, C. and Aubert, S.
(1993) Changes in phenolic content in fresh readyto-use
shredded carrots during storage. Journal of Food Science,
58, 351-356. doi: 10. 1111/j .1365-2621.1993.tb04273.x
[10] Nortje, B.K. and Koeppen, B.H. (1965) The flavanol
glycosides in the fruit of Prunus communis L. cultivar
Bon Chrestien. Biochemist r y J ournal, 95, 209-211.
[11] Manach, C., Scalbert, A., Morand, C., Rémésy, C. and
Jiménez, L. (2004) Polyphenols: Food sources and bioa-
vailability. American Journal of Clinical Nutrition, 79,
727-747.
[12] Balasundram, N., Sundram, K. and Samman, S. (2007).
Phenolic compounds in plants and agri-industrial by
products: Antioxidant activity, occurrence, and potential
uses. Food Chemistry, 99, 191-203.
doi:10.1016/j.foodchem.2005.07.042
[13] Peer, W.A, Brown, D.E., Tague, B.W., Muday, G.K. and
Murpky, A.S. (2001) Flavonoid accumulation patterns of
transparent testa mutants of Arabidopsis. Plant Physiol-
ogy, 126, 536-548. doi:10.1104/pp.126.2.536
[14] HESS, C.E. (1962) A physiological analysis of root ini-
tiation in easy and difficult to root cuttings. 16th Interna-
tional Horticutureae Congress, 4, 375-381.
[15] Bachelard, E.P. and Stowe, B. (1963) Rooting of cuttings
of Acer rubrum L. and eucalyptus camaldulensis Delh.
Australian Journal of Biological Science, 16, 751-767.
[16] Arnold, A.W. and Albert, L.S. (1964) Chemical factors
affecting anthocyanin formation and morphogenesis in
cultured hypocotyl segments of impatiens balsamina.
Plant Physiology, 39, 307-312.
doi:10.1104/pp.39.3.307
[17] Stafford, H.A. (1968) Relationships between the devel-
opment of adventitous roots and biosynthesis of antho-
cyanins in first internodes of Sorghum. Plant Physiology,
43, 318-326. doi:10.1104/pp.43.3.318
[18] Tomaszewski, M. (1964) The mecanism of synergistic
effects between auxin and some natural phenolic sub-
stances. Colloque International C.N.R.S., 123, 335-351.
[19] Bachelard, E.P. (1965) The interrelations between root
formation and anthocyanin synthesis in red maple cutting:
Effects of gibberelic acid, CCC and 8 azaguanine. Aus-
tralian Journal of Biol o g i ca l Science, 18, 699-702.
[20] Fernqvist, I. (1966) Studies on factors in adventitious
root formation. Lantbruskshgskol Ann als, 32, 109-244 .
[21] Paupardin, C. (1969) Sur le moment d’action de l’acide
caféique dans la rhizogenèse de fragments de tubercules
de Tapinambour cultivé in vitro. Comptes. Rendues de
l’Académie des Sciences, Paris, 269, 1532-1534 .
[22] Kraeim, Z., Bouzazi, R. and Ezzili, B. (2006) Influence
of collection date on rhyzogenetic potentiality of Muscat
d’Italy (Vitis vinifera L.) cultivated on sandy substrate.
Arid Zones Workshop Communications, December 2006,
Djerba T un isia.
[23] Ezzili, B. (2001) Essai de régulation du fonctionnement
du méristème terminal, de la dormance des bourgeons et
de l’initiation florale de Vitis vinifera L. Thèse de
doctorat d’état, Faculté Des Sciences, Université Tunis El
Manar, 187.
[24] Friml, J. and Murphy, A.S. (2006) Nomenclature for
apical/basal polarity in plants. In: Taiz, L. and Zeiger, E.,
Eds., Plant Physiology, 4th Edition, Web Essay, Sinauer
Associates, Sunderland.
[25] Dewanto, V., Wu, X., Adom, K.K and Liu, R.H. (2002)
Thermal processing enhaces the nutritional value of to-
matoes by increasing total antioxidant activity. Journal of
Agricultural and Food Chemistry, 50, 3010-3014.
doi:10.1021/jf0115589
[26] Mau, J.L., Chao, G.R. and Wu, K.T. (2001) Antioxidant
properties of methanolic extracts from several ear mush-
rooms. Journal of Agricultural and Food Chemistry, 49,
5461-5467. doi:10.1021/jf010637h
[27] Statsoft, (1998) STATISTICA for Windows (Computer