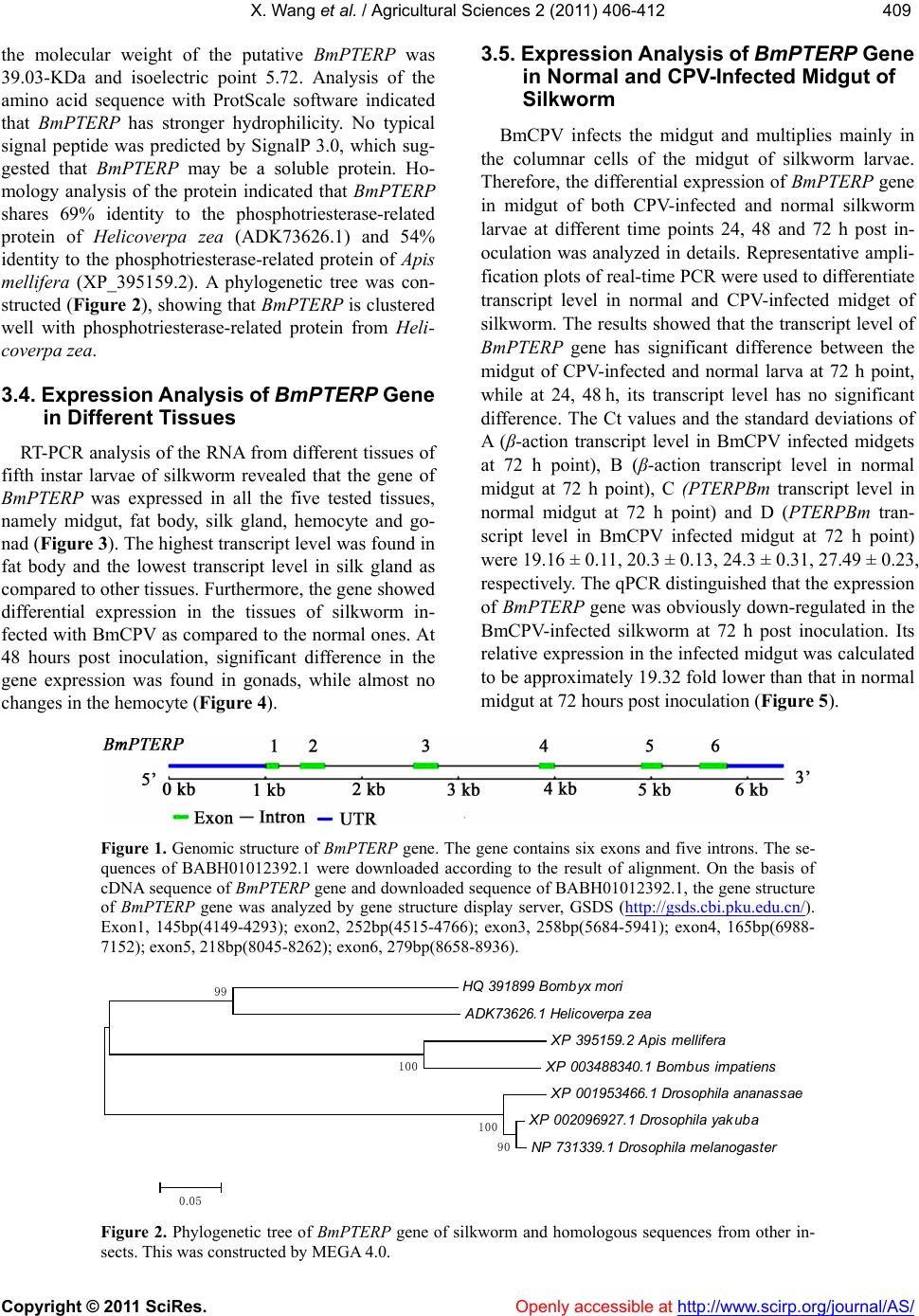
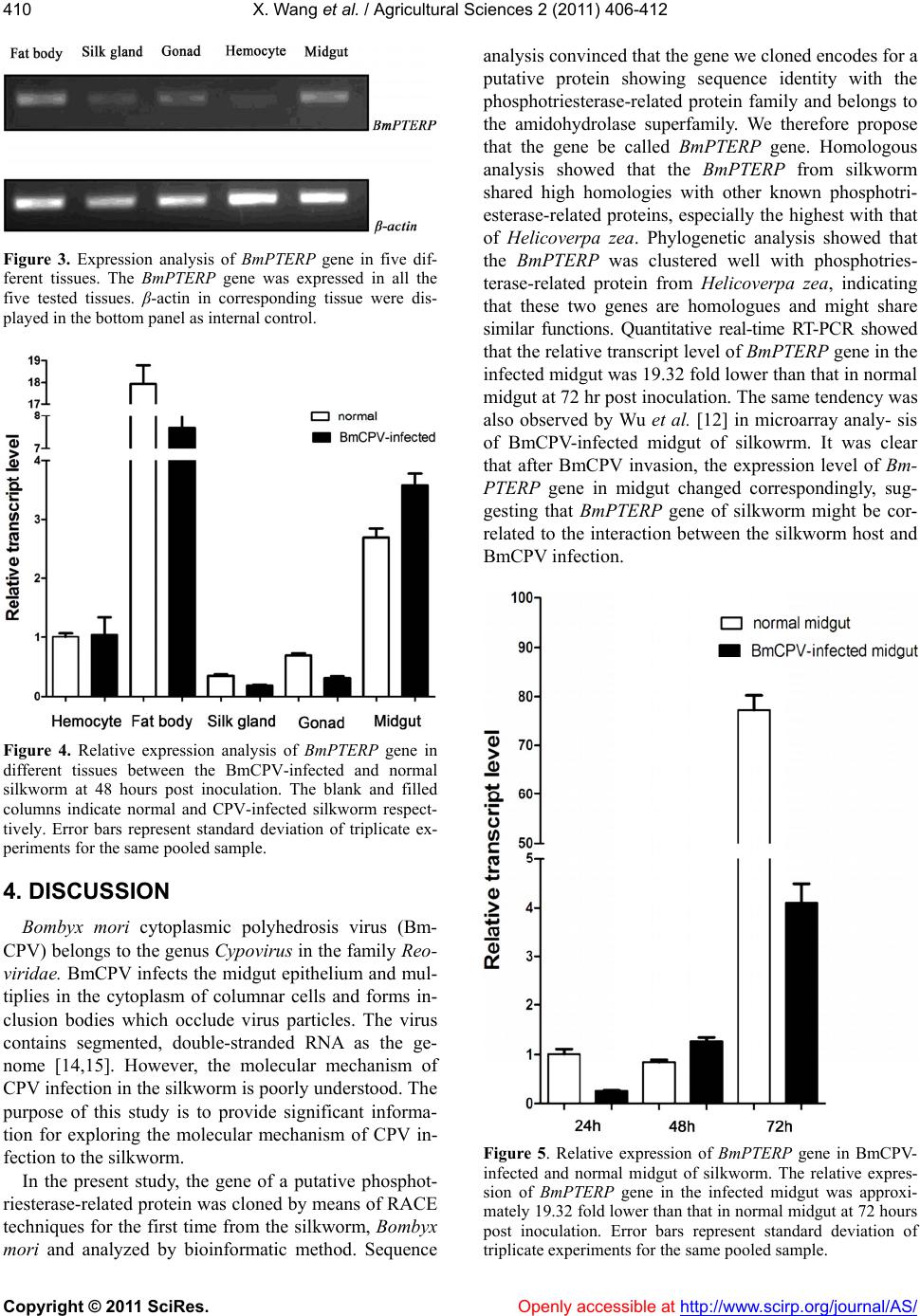
X. Wang et al. / Agricultural Sciences 2 (2011) 406-412
Copyright © 2011 SciRes. Openly accessible at http://www.scirp.org/journal/AS/
411411
Regarding the function of the phosphotriesterase-re-
lated protein, only few reports were available at present.
In mice, phosphotriesterase has hypothetical function
that catalyzes small cytoplasmic molecules which would
prove toxic and protects mice against organophosphate
neurotoxins [16,17]. Attempts to control agricultural and
forest insects and spread of insect-borne diseases, such
as malaria, using organophosphate insecticides are being
frustrated by the development of resistant strains of in-
sects. Some of these achieve resistance by modification
of acetylcholinesterase, others show a phosphotriesterase
activity apparently different from that of bacterial phos-
photriesterases [18]. However, no activity of the phos-
photriesterase-related protein was commonly detected,
except for a weak esterase activity and PTE activity in
an E. coli PHP mutant [1]. Recently, a PHP gene from
the thermophilic bacterium Geobacillus caldoxylosilyti-
cus TK4 was cloned and overexpressed in E. coli [19].
The recombinant protein showed activities with p-ni-
trophenyl acetate and p-nitrophenyl butyrate. This is the
first reported PHP having an extremely pH- and thermo-
stable esterase activity.
Our result in the present study is the first report that a
phosphotriesterase-related protein gene was cloned from
the silkworm. While, its in vivo function still remains
unclear. At present, the only clue comes from its strong
homology to PHPs of bacteria and other organisms, but
there is not yet any experimental proof that BmPETRP is
itself a phosphotriesterase-related protein. As the disease
of silkworm caused by the infection with BmCPV ad-
vanced, the expression of the BmPETRP gene was ob-
viously down-regulated. This might be attributed to the
facts that a series of physiological and pathological
changes takes place due to the infection. Further func-
tional experimental research should be addressed in our
future work. If BmPETRP does turn out to have the ac-
tivities of PHP or phosphotriesterase predicted from its
homology, it would be very interesting to establish
whether insects have homologous genes, whether these
genes might confer resistance to organophosphate pesti-
cides and most importantly whether these genes might
be involved in the interaction between the silkworm and
the BmCPV infecton.
5. ACKNOWLEDGEMENTS
This research was supported by the National Natural Science Foun-
dation of China (Grant No.30972143) and Natural Science Foundation
of Jiangsu Province (Grant No. BK2010353).
REFERENCES
[1] Roodveldt, C. and Tawfik, D.S. (2005) Shared promis-
cuous activities and evolutionary features in various
members of the amidohydrolase superfamily. Biochemi s-
try, 44, 12728-12736. doi:10.1021/bi051021e
[2] Chen, S.L., Fang, W.H. and Himo, F. (2007) Theoretical
study of the phosphotriesterase reaction mechanism. The
Journal of Physical Chemistry, 111, 1253-1255.
doi:10.1021/jp068500n
[3] Porzio, E., Merone, L., Mandrich, L., Rossi, M. and
Manco, G. (2007) A new phosphotriesterase from Sul-
folobus acidocaldarius and its comparison with the
homologue from Sulfolobus solfata. Biochimie, 89, 625-
636. doi:10.1016/j.biochi.2007.01.007
[4] Merone, L., Mandrich, L. and Rossi, M. (2005) A ther-
mostable phosphotriesterase from the archaeon Sul-
folobus solfataricus: Cloning, overexpression and prop-
erties. Extremophile, 9, 297-305.
doi:10.1007/s00792-005-0445-4
[5] Griffiths, A.D. and Tawfik, D.S. (2003) Directed evolu-
tion of an extremely fast phosphotriesterase by in vitro
compartmentalization. The EMBO Journal, 22, 24-35.
doi:10.1093/emboj/cdg014
[6] Roodveldt, C. and Tawfik, D.S. (2005b) Directed evolu-
tion of phosphotriesterase from Pseudomonas diminuta
for heterologous expression in Escherichia coli results in
stabilization of the metal-free state. Protein Engineering,
Design & Selection, 18, 51-58.
doi:10.1093/protein/gzi005
[7] Buchbinder, J.L., Stephenson, R.C., Dresser, M.J., Pitera,
J.W., Scanlan, T. S. and Fletterick, R.J. (1998) Bio-
chemical characterization and crystallographic structure
of an Escherichia coli protein from the phosphotri-
esterase gene family. Biochemistry, 37, 5096-5106.
doi:10.1021/bi971707+
[8] Scanlan, T.S. and Reid, R.C. (1995) Evolution in action.
Chemistry & Biology, 2, 71-75.
doi:10.1016/1074-5521(95)90278-3
[9] Hou, X.Y., Maser, R.L., Magenheimer, B.S. and Calvet,
J.P. (1996) A mouse kidney- and liver-expressed cDNA
having homology with a prokaryotic parathion hydrolase
(phosphotriesterase)-encoding gene: Abnormal expres-
sion in injured and polycystic kidneys. Gene, 168, 157-
163. doi:10.1016/0378-1119(95)00746-6
[10] Ikeda, K., Nagaoka, S., Winkler, S., Kotani, K., Yagi, H.,
Nakanishi, K., Miyajima, S., Kobayashi, J. and Mori, H.
(2001) Molecular characterization of Bombyx mori cyto-
plasmic polyhedrosis virus genome segment 4. Journal
of Virology, 75, 988-995.
doi:10.1128/JVI.75.2.988-995.2001
[11] Qanungo, K.R., Kundu, S.C., Mullins, J.I. and Ghosh,
A.K. (2002) Molecular cloning and characterization of
Antheraea mylitta cytoplasmic polyhedrosis virus ge-
nome segment 9. Journal of General Virology, 83, 1483-
1491.
[12] Wu, P., Wang, X., Qin, G.X., Liu, T., Jiang, Y.F., Li, M.W.
and Guo, X.J. (2011) Microarray analysis of gene ex-
pression profile in the midgut of silkworm infected with
cytoplasmic polyhedrosis virus. Molecular Biology Re-
ports, 38, 333-341.
doi:10.1007/s11033-010-0112-4
[13] Livak, K.J. and Schmittgen, T.D. (2001) Analysis of
relative gene expression data using real-time quantitative
PCR and the 2−ΔΔCT Method. Methods, 25, 402-408.
doi:10.1006/meth.2001.1262