S. Behbahani et al. / J. Biomedical Science and Engineering 2 (2009) 480-483
SciRes Copyright © 2009
483
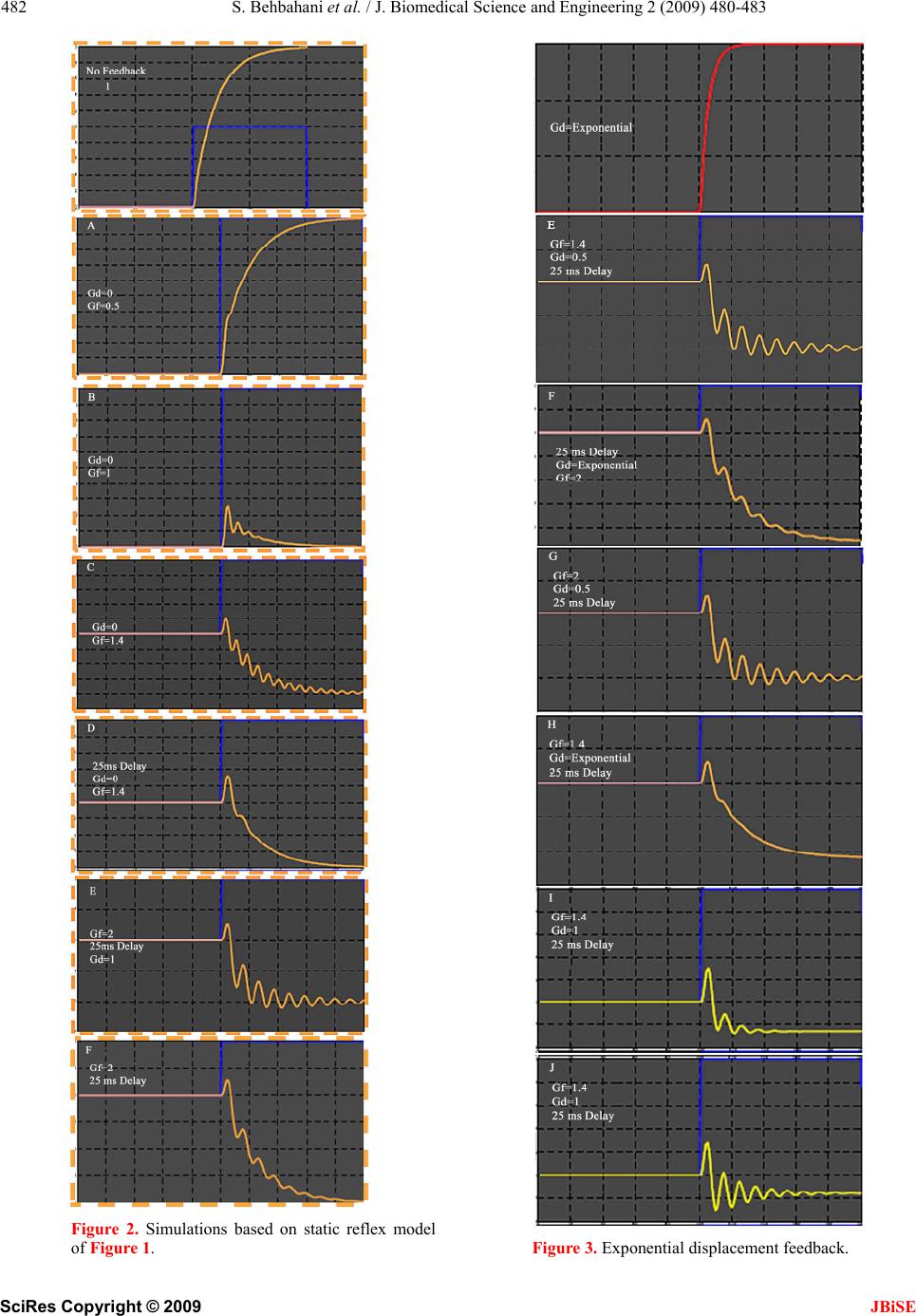
isometric frequency response characteristic of cat triceps
surae muscles of Rosenthal that positive force feedback
may be appropriate in some et al. 1970. A mass of 1 kg
represents the inertial load borne by a cat hind limb. A
force increment of 10 N represents the mean force de-
veloped by triceps surae dur ing the stance phase of slow
gait [11]. Inherent muscle properties are simplified to a
linear viscoelastic element with a stiffness of 0.5 N/mm.
The force feedback and displacement feedback signals
are represented without dynamics. It was clear that the
reflex mechanism in question represented positive feed-
back and this was normally associated with instability. It
was tacitly assumed that the nervous system would
somehow always limit positive force feedback gain
within a range consistent with stability. Our results sug-
gest that a combination of intrinsic muscle properties,
concomitant negative displacement feedback, and reflex
delays found in neuromuscular may provide this auto-
matic gain control.
JBiSE
[3] Donelan, J. M. and Pearson, K. G., (2004) Contribution
of sensory feedback to ongoing ankle extensor activity
during the stance phase of walking, Can. J. Physiol.
Pharmacol, 82, 589–598.
Our experiments and analysis verify that positive
force feedback in the neuromuscular system can provide
stable and effective load compensation. The analysis
also shows that the conclusions regarding the stabilizing
influence of muscle intrinsic properties, leng th feedback,
and delays in positive feedb ack pathways were robust in
the face of large parametric and structural variations in
the systems considered. Stable behavior for large values
of positive feedback gains was unexpected and initially
quite puzzling. However, it became apparent that loop
gain did not remain high, but rather it was automatically
attenuated when muscles shortened and thereby reduced
their force producing capability.
REFERENCES
[1] Brownstone, R. M., Gossard, J. P., and Hultborn, H.,
(1994) Voltage-dependent excitation of motoneurones
from spinal locomotor centres in the cat, Springer-Verlag,
Exp. Brain Res., 102, 3444.
[2] Grey, M. J., Mazzaro, N., Nielsen, J. B., and Sinkjær, T.,
(2004) Ankle extensor proprioceptors contribute to the
enhancement of the soleus EMG during the stance phase
of human walking1, Can. J. Physiol. Pharmacol., 82,
610–616.
[4] Frigon, A. and Rossignol, S., (2006) Experiments and
models of sensorimotor interactions during locomotion,
Springer Berlin/Heidelberg, Saturday, DOI: 10.1007/s
00422-006-0129-x.
[5] Prochazka, A. and Gillard, D., (1997) Sensory control of
locomotion, Proceeding of American Control Confer-
ence.
[6] Prochazka, A., Gillard, D., and Bennet, D. J., (1997)
Implications of positive feedback in the control of
movement, the American Physiological Society
0022-3077/97, 5.00 Copyright.
[7] Taylor, A., Ellaway, P. H., Durbaba, R., and Rawlinson,
S., (2000) Distinctive patterns of static and dynamic
gamma motor activity during locomotion in the decere-
brate cat, Journal of Physiology, 529(3), pp. 825–836.
[8] Raya, J. G., Ramı´rez, A., and Mun˜oz-Martı´nez, E. J.,
(2004) Gamma 3 alpha linkage and persistent firing of Ia
fibers by pudendal nerve stimulation in the decerebrate
cat, J. Neurophysiol, 92, 387–394.
[9] Krakauer, J. W., MD, (2006) Spinal cord physiology:
Receptors and reflexes.
[10] Csercsik, D., (2006) Construction of simple dynamic
models of the gamma-loop mechanism, Research Report
SCL-001.
[11] Knutson, G. A., Edward, D. C., and Owens, F., MS, DC,
(2003) Active and passive characteristics of muscle tone
and their relationship to models of subluxation, 0008-
3194/2003/269–283/$2.00/\JCCA.