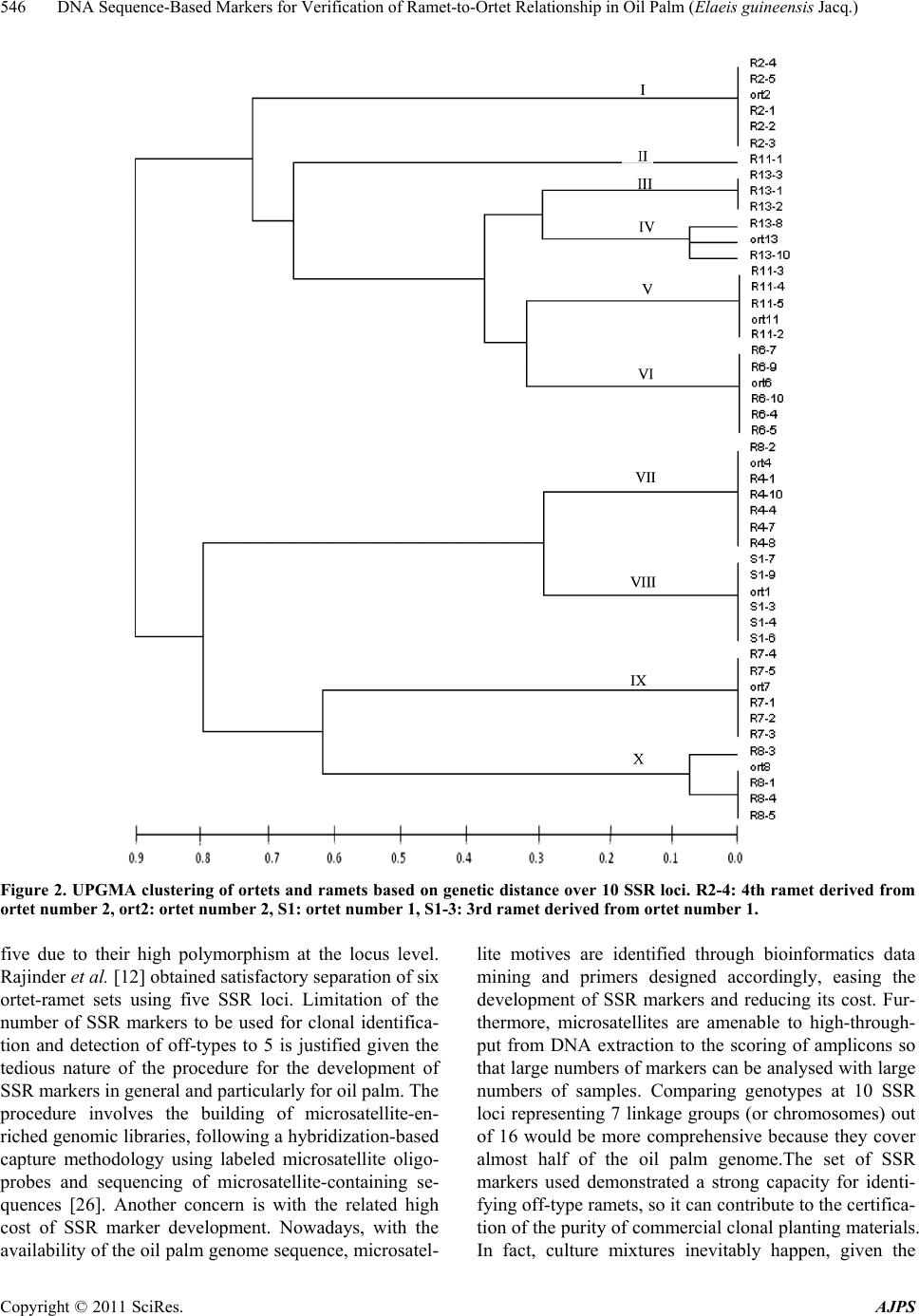
DNA Sequence-Based Markers for Verification of Ramet-to-Ortet Relationship in Oil Palm (Elaeis guineensis Jacq.)
Copyright © 2011 SciRes. AJPS
548
of the PORIM Oil Palm Congress, Kuala Lumpur, 1999,
pp. 330-343.
[7] A. C. Soh, Y. Y. Yong, Y. W. Ho and N. Rajanaidu,
“Commercial Potential of Oil Palm Clones: Early Results
of Their Performances,” In: V. Rao, I. E. Henson and N.
Rajanaidu, Eds., Recent Developments in Oil Palm Tissue
Culture and Technology, Palm Oil Research Institute of
Malaysia, Kuala Lumpur, 1995, pp. 134-144.
[8] A. C. Soh, G. Wong, C. C. Tan, P. S. Chew, T. Y. Hor, S.
P. Chong and K. Gopal, “Recent Advances towards Com-
Mercial Production of Elite Oil Palm Clones,” Proceed-
ings of the MPOB 2001 International Palm Oil Con-
gress―Agriculture, Kuala Lumpur, 2001, pp. 33-44.
[9] C. J. Eeuwens, S. Lord, C. R. Donough, V. Rao, G.
Vallejo and S. Nelson, “Effects of Tissue Culture Condi-
Tions during Embryoid Multiplication on the Incidence of
‘Mantled’ Flowering in Clonal Propagated Oil Palm,”
Cell Plant, Tissue Organ Culture, Vol. 70, 2002, pp.
311-323. doi:10.1023/A:1016543921508
[10] S. Jouannic, X. Argout, F. Lechauve, C. Fizames, A.
Borgel, F. Morcillo, F. Aberlec-Bertossi, Y. Duval and J.
W. Tregear, “Analysis of Expressed Sequence Tags from
Oil Palm (Elaeis guineensis),” FEBS Letters, Vol. 579,
2005, pp. 2709-2714. doi:10.1016/j.febslet.2005.03.093
[11] A. C. Soh, G. Wong, C. C. Tan, T. Y. Hor and C. K.
Wong, “Revisited: Cloning Seedling of Reproduced Best
Dxp Cross Strategy,” Paper Presented at ISOPB Seminar
on the Progress of Oil Palm Breeding and Selection,
Medan, 6-9 October 2003.
[12] S. Rajinder, N. Jayanthi, T. Soon-Guan, M. P. Jothi and S.
C. Cheah, “Development of Simple Sequence Repeat
(SSR) Markers for Oil Palm and Their Application in
Genetic Mapping and Fingerprinting of Tissue Culture
Clones,” Asia Pacific Journal of Molecular Biology and
Biotechnology, Vol. 15, 2007, pp. 121-131.
[13] M. Delseny, M. Laroche and P. Penon, “Detection of
Sequences with Z-DNA Forming Potential in High
Plants,” Biochemical and Biosphysical Research Com-
mununications, Vol. 116, 1983, pp. 113-120.
doi:10.1016/0006-291X(83)90388-1
[14] W. Amos, S. J. Sawcer, R. W. Feakes and D. C.
Rubinsztein, “Microsatellites Show Mutational Bias and
Heterozygote Instability,” Nature Genetics, Vol. 13, 1996,
pp. 390-391. doi:10.1038/ng0896-390
[15] J. S. C. Smith, E. C. L. Chin, H. Shu, O. S. Smith, S. J.
Wall, M. L. Senior, S. E. Mitchel, S. Kresovich and J.
Ziegle, “An Evaluation of the Utility of SSR Loci as Mo-
Lecular Markers in Maize (Zea mays L.), Comparisons
with Data from RFLPs and Pedigree,” Theoretical and
Applied Genetics, Vol. 95, 1997, pp. 163-173.
doi:10.1007/s001220050544
[16] J. Doyle and L. Doyle, “Isolation of Plant DNA from
Fresh Tissue,” Focus, Vol. 12, 1990, pp. 13-15.
[17] N. Billotte, N. A. M. Risterucci, E. Barcelos, L. Noyer, P.
Amblard and F. C. Baurens, “Development, Characterisa-
tion, and Across-Taxa Utility of Oil Palm (Elaeis
guineensis Jacq.) Microsatellite Markers,” Genome, Vol.
44, 2001, pp. 413-425.
[18] M. Nei and N. Takezaki, “Estimation of Genetic Dis-
tances and Phylogenetic Trees from DNA Analysis,”
Proceedings of the 5th World Congress on Genetics. Ap-
plied to Livestock Production, Vol. 21, 1983, pp. 405-
412.
[19] P. H. A. Sneath and R. R. Sokal, “Numerical Taxonomy,”
W. H. Freeman Press, San Francisco, 1973.
[20] B. S. Weir, “Genetic Data Analysis II,” MA: Sinauer As-
sociates, Inc., Sunderland, 1996.
[21] M. Nei, “Analysis of Gene Diversity in Subdivided
Populations,” Proceedings of the National Academy of
Science of the USA, Vol. 70, 1973, pp. 3321-3323.
doi:10.1073/pnas.70.12.3321
[22] K. Liu and S. V. Muse, “PowerMarker: Intergrated
Analysis Environment for Genetic Marker Data,” Bioin-
formatics, Vol. 21, 2005, pp. 2128-2129.
doi:10.1093/bioinformatics/bti282
[23] B. Cochard, “Etude de la Diversité Génétique et du
Déséquilibre de Liaison au Sein de Populations
Améliorées de Palmier à Huile (Elaeis guineensis Jacq.),”
Ph.D. Dissertation, SUPAGRO, Montpellier, 2008.
[24] F. C. Yeh, “Population Genetics,” In: A. Young, D. Bosh-
ier and T. Boyle, Eds., Forest Conservation Genetics:
Principles and Practice, CSIRO Publishing, Collingwood,
2000, pp. 21-37.
[25] R. V. H. Corley, “Illegitimacy in Oil Palm Breeding,”
Journal of Oil Palm Research, Vol. 17, 2005, pp. 64-69.
[26] N. Billotte, P. J. L. Lagoda, A. M. Risterucci and F. P.
Baurens, “Microstellite-Enriched Libraries: Applied
Methodology for the Development of SSR Markers in
Tropical Crops,” Fruits, Vol. 54, 1999, pp. 277-288.
[27] C. Bakoumé, R. Wickneswari, N. Rajanaidu, A. Kushairi,
P. Amblard and N Billotte, “Allelic Diversity of Natural
Oil Palm (Elaeis guineensis Jacq.) Populations Detected
by Microsatellite Markers: Implication in Conservation,”
Plant Genetic Resources Characterization and Utilization,
Vol. 5, 2007, pp. 104-107.
doi:10.1017/S1479262107710870
[28] C. W. S. Hartley, “The Oil Palm (Elaeis guineensis
Jacq.),” Longman Scientific and Technical, New York,
1988.
[29] C. Bakoumé, “Genetic Diversity of Natural Oil Palm
(Elaeis guineensis Jacq.) Populations Using Microsatel-
lite Markers,” unpublished Ph.D. dissertation, Universiti
Kebangsaan Malaysia, Bangi, 2006.
[30] S. Diabate, H. De Franqueville, D. Allou, K. E. Konan, O.
A. Coulibaly and W. P. N’Guessan, “Phenolic Diversity
in the Defense Reaction of the Oil Palm against Vascular
Wilt Disease,” Proceedings of the 2009 International
Palm Oil Congress on Palm Oil: Balancing Ecologics
with Economics, Kuala Lumpur, 2009, pp. 1027-1034.