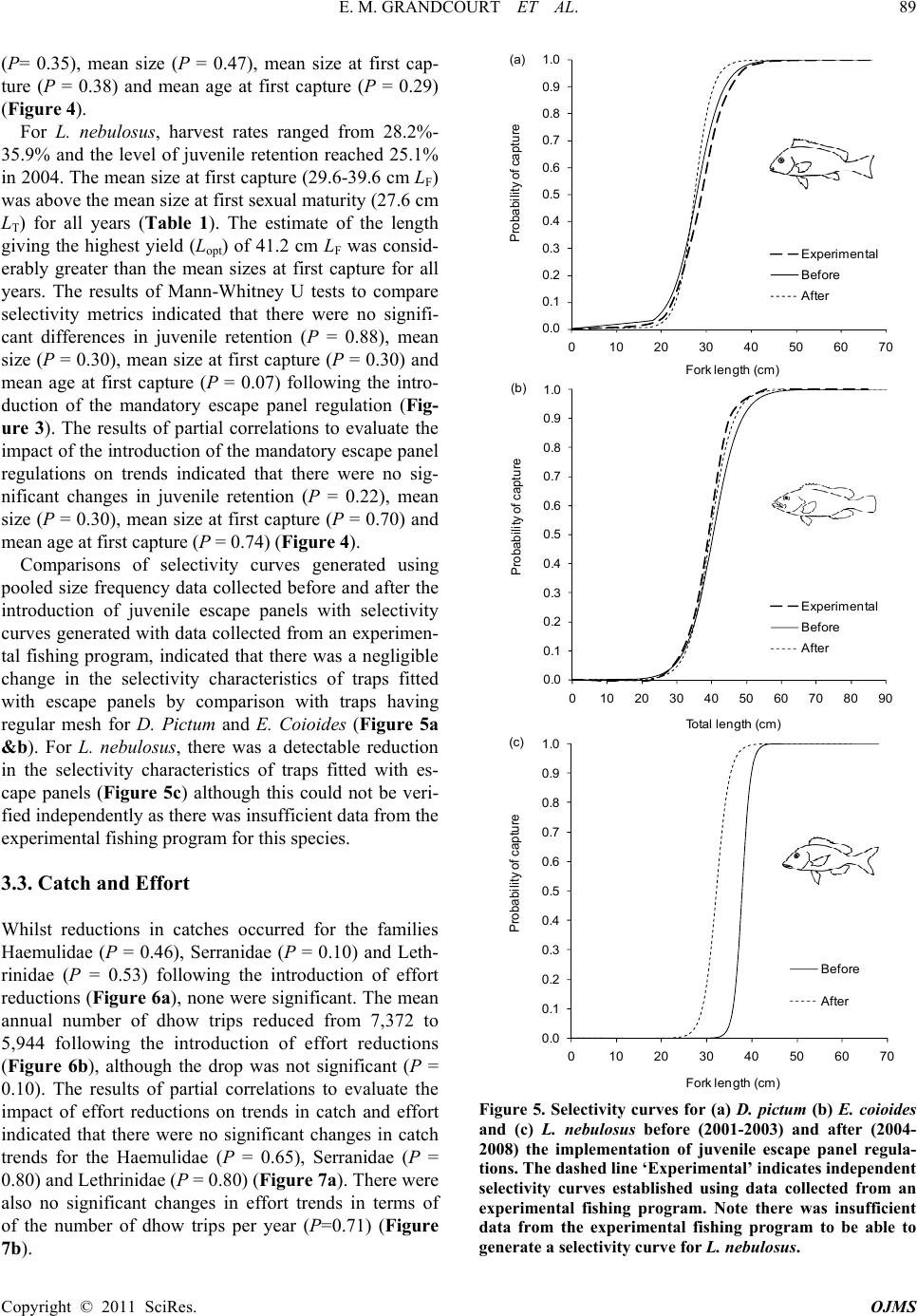
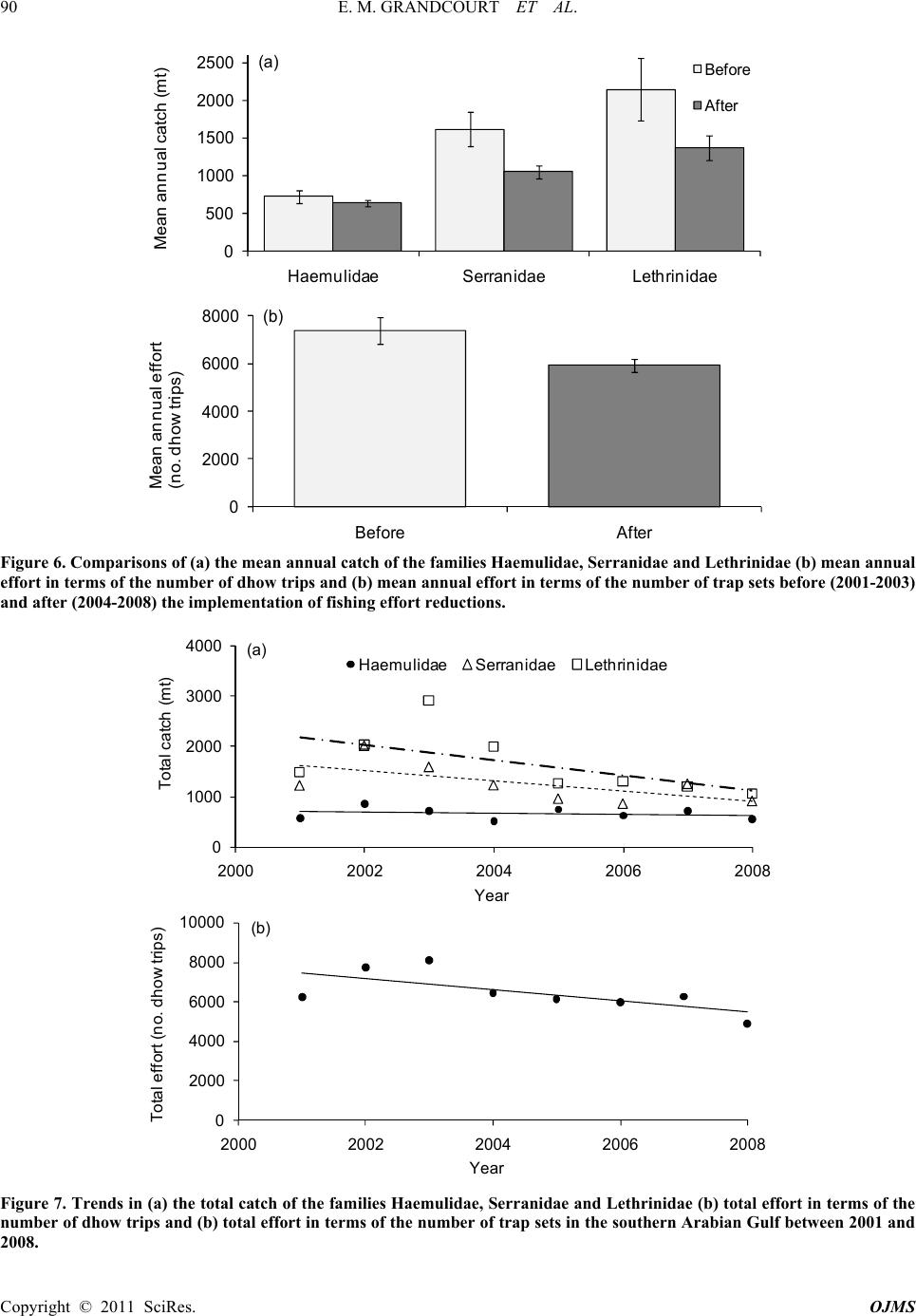
E. M. GRANDCOURT ET AL.
96
Copyright © 2011 SciRes. OJMS
introduction of regulations, in contrast to the intended
impact.
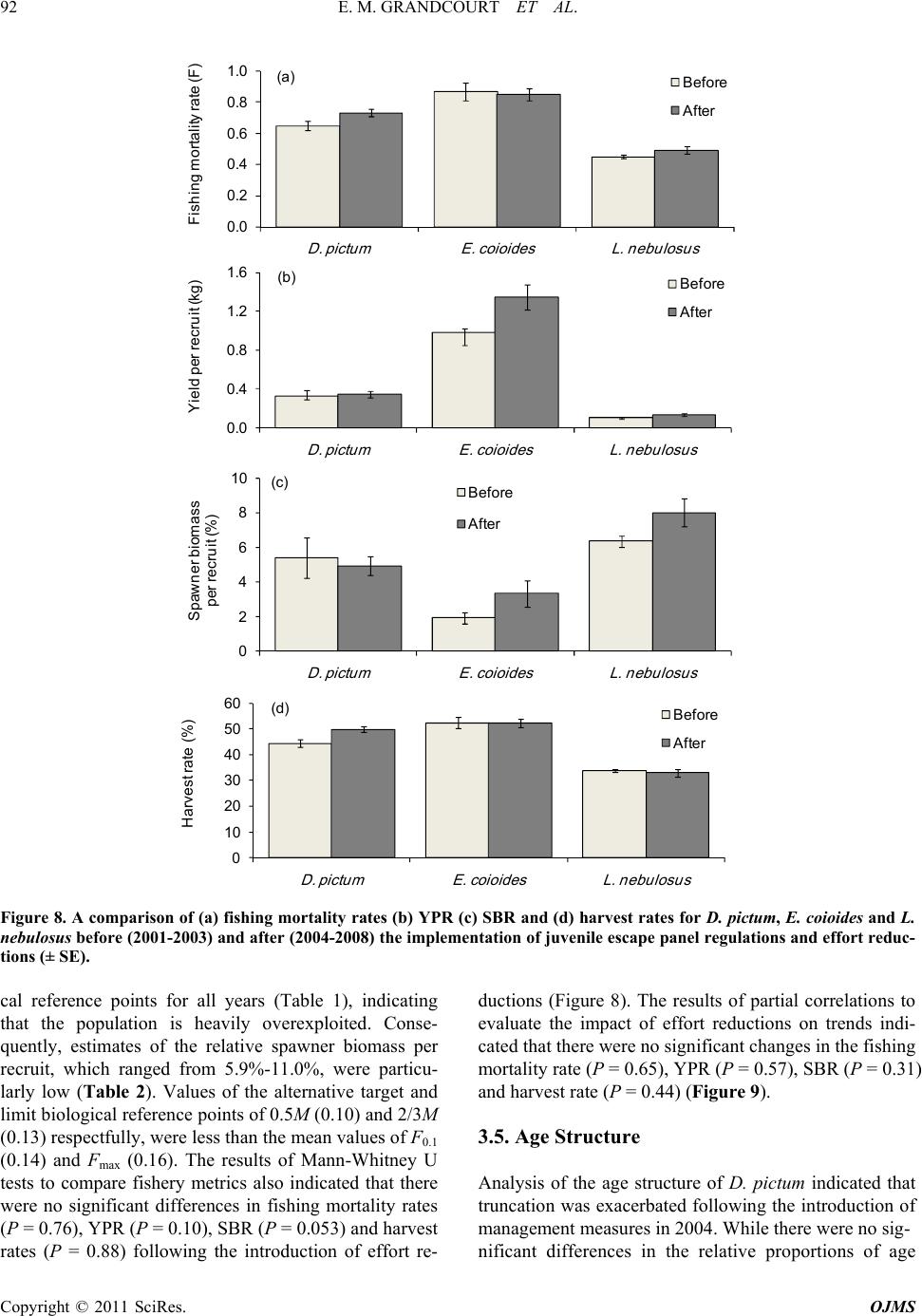
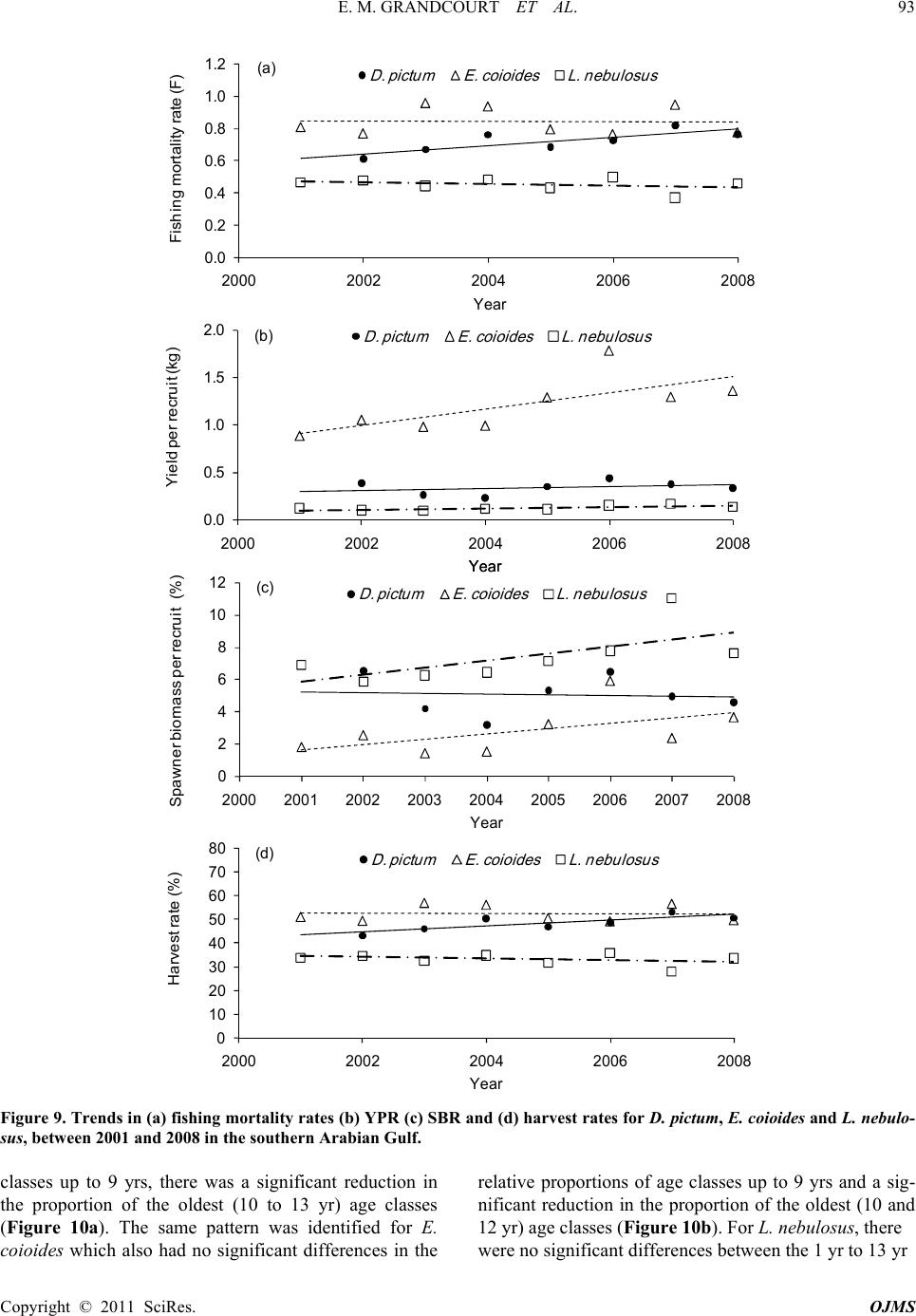
YPR and SBR analyses indicated that the study spe-
cies were all heavily over-exploited. As the existing fi-
shing mortality rates were all well in excess of the limit
biological reference points, growth over-fishing had
clearly occurred. Furthermore, with current values of the
relative spawner biomass per recruit being less than 10%
in all cases, recruitment over-fishing would also have
occurred, based on meta-analyses [22]. The critical ma-
nagement issues that relate to the stock status established
in earlier assessments [3,7] are clearly still pertinent.
There is widespread recognition that target species are
over-exploited in places where wire-mesh fish traps have
been used extensively [23,24]. Given the failure of ex-
isting regulations to modify gear selectivity, reduce ef-
fort and rebuild stocks, management authorities should
consider alternative measures including a moratorium on
the use of traps in the off-shore demersal fishery of Abu
Dhabi.
5. Acknowledgements
This study forms part of the activities of the ‘Fish Land-
ings and Population Dynamics Project’ (project # 02-
23-0008-09) which is implemented by the Biodiversity
Management Sector―Marine, of the Environment Agen-
cy―Abu Dhabi. The management of the Environment
Agency―Abu Dhabi is thanked for their support for this
work. The Abu Dhabi Fishermen’s Cooperative Society
facilitated size frequency sampling and catch data collec-
tion. The UAE Coast Guard and later CNIA (Critical
National Infrastructure Authority) provided effort re-
cords. All the EAD enumerators are thanked for their
contribution to data collection; Khalid Al Ali, Sultan Al
Ali, Hamad Al Shamsi, Mohamed Al Zaabi, Sabah Has-
san, Jihad Hassan, Mohamed Ahmed Hassan, Abdulla Al
Blooki, Abdulla Muhyiddeen, Eid Al Romaithi, Yaqoob
Yousif Al Hammadi.
6. References
[1] E. M. Grandcourt, “Fish and Fisheries,” In: T. Z. Al Ab-
dessalaam, Ed., Marine Environment and Resources of
Abu Dhabi, Motivate Publishing, Dubai, 2008, pp. 200-
225.
[2] B. Shallard, “Distribution and Abundance of Demersal
Fish Stocks in the UAE. Technical Report 1. Fish Re-
sources Assessment Survey Project of Abu Dhabi and
UAE Waters,” Environmental Research and Wildlife
Development Agency, Government of Abu Dhabi, Abu
Dhabi, 2003, p. 211.
[3] E. M. Grandcourt, T. Z. Al Abdessalaam, A. T. Al Shamsi
and F. Francis, “Biology and Assessment of the Painted
Sweetlips (Diagramma pictum (Thunberg, 1792)) and
Spangled Emperor (Lethrinus nebulosus (Forsskål, 1775))
in the Southern Arabian Gulf,” Fisheries Bulletin, Vol.
104, No. 1, 2006, pp. 75-88.
[4] E. M. Grandcourt, T. Z. Al Abdessalaam, F. Francis and
A. Al Shamsi, “Population Biology and Assessment of
the White-Spotted Spinefoot, Siganus Canaliculatus (Park,
1797), in the southern Arabian Gulf,” Journal of Applied
Ichthyology, Vol. 23, No. 1, 2007, pp. 53-59.
doi:10.1111/j.1439-0426.2006.00796.x
[5] E. M. Grandcourt, T. Z. Al Abdessalaam, F. Francis and
A. Al Shamsi, “Biology and Stock Assessment of the
Sparids, Acanthopagrus Bifasciatus and Argyrops Spini-
fer (Forsskål, 1775), in the Southern Arabian Gulf,” Fi-
sheries Research, Vol. 69, No. 1, 2004, pp. 7-20.
doi:10.1016/j.fishres.2004.04.006
[6] H. Al Masroori, H. Al Oufi, J. McIlwain and E. McLean,
“Catches of Lost Fish Traps (ghost fishing) from Fishing
Grounds near Muscat, Sultanate of Oman,” Fisheries Re-
search, Vol. 69, No. 3, 2004, pp. 407-414.
doi:10.1016/j.fishres.2004.05.014
[7] E. M. Grandcourt, T. Z. Al Abdessalaam, A. T. Al Shamsi
and F. Francis, “Population Biology and Assessment of
the Orange-Spotted Grouper, Epinephelus coioides (Ha-
milton, 1822), in the Southern Arabian Gulf,” Fisheries
Research, Vol. 74, No. 1, 2005, pp. 55-68.
doi:10.1016/j.fishres.2005.04.009
[8] R. J. Beamish and D. A. Fournier, “A Method for Com-
paring the Precision of a Set of Age Determinations,”
Canadian Journal of Fisheries and Aquatic Sciences, Vol.
38, No. 8, 1981, pp. 982-983. doi:10.1139/f81-132
[9] W. E. Ricker, “Computation and Interpretation of Bio-
logical Statistics of Fish Populations,” Bulletin of the
Fisheries Research Board of Canada, Vol. 191, 1975, pp.
2-6.
[10] R. J. H. Beverton and S. J. Holt, “On the Dynamics of
Exploited Fish Populations,” Chapman and Hall, London,
1957, p. 179.
[11] F. C. Gayanilo Jr. and D. Pauly, “FAO-ICLARM Stock
Assessment Tools, Reference Manual, ICLARM,” Food
and Agricultural Organization of the United Nations, Rome,
1997.
[12] D. Pauly, “Length-Converted Catch Curves: A Powerful
Tool for Fisheries Research in the Tropics (Part 1),”
Fishbyte, Vol. 1, No. 2, 1983, pp. 9-13.
[13] R. J. H. Beverton, “Patterns of Reproductive Strategy
Parameters in Some Marine Teleost Fish,” Journal of
Fish Biology, Vol. 41, No. B, 1992, pp. 137-160.
[14] E. M. Grandcourt, T. Z. Al Abdessalaam, F. Francis, A. T.
Al Shamsi and S. A. Hartmann, “Reproductive Biology
and Implications for Management of the Orange-Spotted
Grouper, Epinephelus coioides (Hamilton, 1822), in the
southern Arabian Gulf,” Journal of Fish Biology, Vol. 74,
2009, pp. 820-841.
doi:10.1111/j.1095-8649.2008.02163.x
[15] K. Patterson, “Fisheries for Small Pelagic Species: An
Empirical Approach to Management Targets,” Reviews in
Fish Biology and Fisheries, Vol. 2, No. 4, 1992, pp.