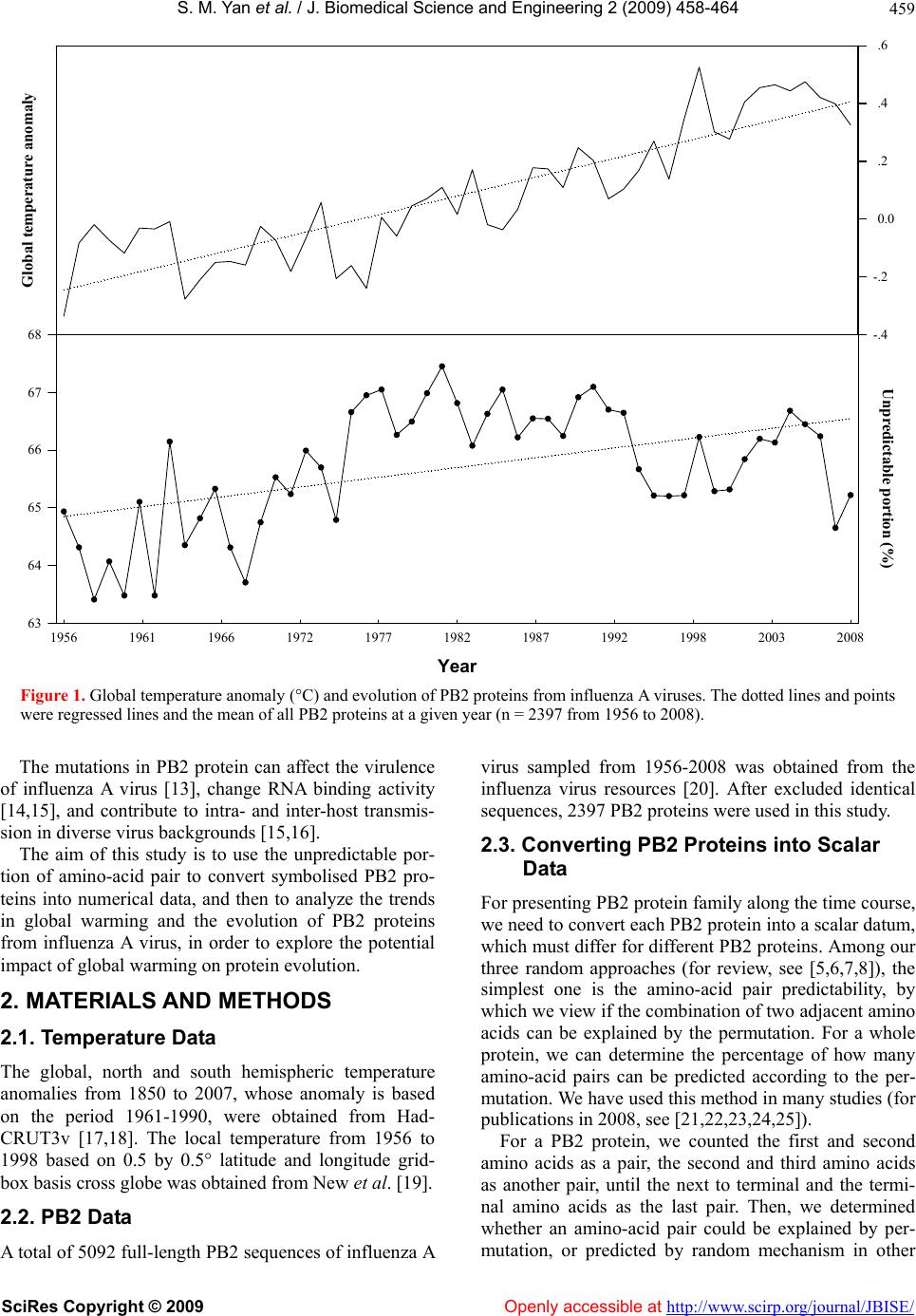
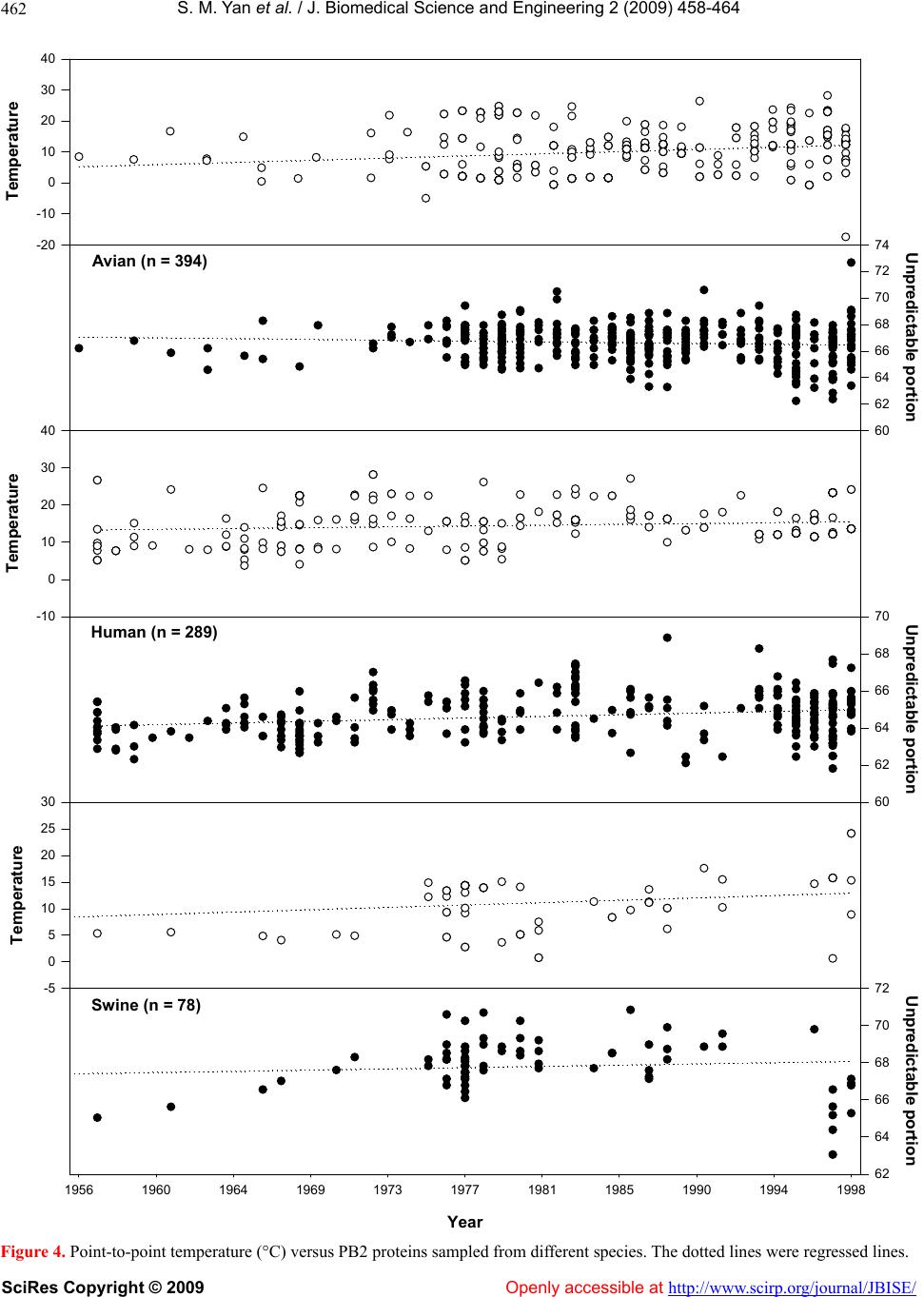
S. M. Yan et al. / J. Biomedical Science and Engineerin g 2 (2009) 458-464
SciRes Copyright © 2009 Openly accessible at http://www.scirp.org/journal/JBISE/
464
[6] G. Wu and S. Yan, (2006) Fate of influenza A virus pro-
teins, Protein Pept. Lett., 13, 399–406.
[7] G. Wu and S. Yan, (2006) Mutation trend of hemaggluti-
nin of influenza a virus: A review from computational
mutation viewpoint, Acta Pharmacol, Sin., 27, 513–526.
[8] G. Wu and S. Yan, (2008) Lecture notes on computational
mutation, Nova Science Publishers, New York.
[9] O. G. Engelhardt and E. Fodor, (2006) Functional asso-
ciation between viral and cellular transcription during in-
fluenza virus infection, Rev. Med. Virol., 16, 329–345.
[10] J. N. Heme rka, D. Wa ng, Y. Weng, W. Lu, R. S. Kaushik,
J. Jin, A. F. Harmon, and F. Li, (2009) Detection and
characterization of influenza A virus PA-PB2 interaction
through a bimolecular fluorescence complementation as-
say, J. Virol., 83, 3944–3955.
[11] N. Van Hoeven, C. Pappas, J. A. Belser, T. R. Maines, H.
Zeng, A. García-Sastre, R. Sasisekharan, J. M. Katz, and
T. M. Tumpey, (2009) Human HA and polymerase sub-
unit PB2 proteins confer transmission of an avian influ-
enza virus through the air, Proc. Natl. Acad. Sci., U. S. A.,
106, 3366–3371.
[12] T. Watanabe, S. Watanabe, K. Shinya, J. H. Kim, M.
Hatta & Y. Kawaoka, (2009) Viral RNA polymerase com -
plex promotes optimal growth of 1918 virus in the lower
respiratory tract of ferrets, Proc. Natl. Acad. Sci. U. S. A.,
106, 588–592.
[13] M. Hatta and Y. Kawaoka, (2002) The continued pan-
demic threat posed by avian influenza viruses in Hong
Kong. T rends Microbiol., 10, 340–344.
[14] T. Kuzuhara, D. Kise, H. Yoshida, T. Horita, Y. Murazaki,
A. Nishimura, N. Echigo, H. Utsunomiya, and H. Tsuge,
(2009) Structural basis of the influenza A virus RNA po-
lymerase PB2 RNA-binding domain containing the
pathogenicity-determinant lysine 627 residue, J. Biol.
Chem., 284, 6855–6860.
[15] J. Steel, A. C. Lowen, L. Pena, M. Angel, A. Solórzano,
R. Albrecht, D. R. Perez, A. García-Sastre, and P. Palese,
(2009) Live attenuated influenza viruses containing NS1
truncations as vaccine candidates against H5N1 highly
pathogenic avian influenza. J. Virol., 83, 1742–1753.
[16] Q. M. Le, Y. Sakai-Tagawa, M. Ozawa, M. Ito, and Y.
Kawaoka, (2009) Selection of H5N1 influenza virus PB2
during replication in humans, J. Virol., 83, 5278–5281.
[17] N. A. Rayner, P. Brohan, D. E. Parker, C. K. Folland, J. J.
Kennedy, M. Vanicek, T. Ansell, and S. F. B. Tett, (2006)
Improved analyses of changes and uncertainties in ma-
rine temperature measured in situ since the mid nine-
teenth century: The HadSST2 dataset, J. Clim., 19, 446–
469.
[18] Climatic Research Unit, (2009)
http://www.cru.uea.ac.uk/cru/data/temperature/.
[19] M. New, M. Hulme, and P. Jones, (2000) Representing
twentieth-century space-time climate variability, Part II:
Development of 1901-96 monthly grids of terrestrial
surface climate, J. Clim., 13, 2217–2238.
[20] Influenza virus resources, (2009)
http://www.ncbi.nlm.nih.gov/genomes/FLU/Database/mu
ltiple.cgi.
[21] G. Wu and S. Yan, (2008) Prediction of mutations engi-
neered by randomness in H5N1 neuraminidases from in-
fluenza a virus . Amino Acids, 34, 81–90.
[22] G. Wu and S. Yan, (2008) Prediction of mutations initi-
ated by internal power in H3N2 hemagglutinins of influ-
enza a virus from North America, Int. J. Pept. Res. Ther.,
14, 41–51.
[23] G. Wu and S. Yan, (2008) Prediction of mutation in
H3N2 hemagglutinins of influenza A virus from North
America based on different datasets, Protein Pept. Lett.,
15, 144–152.
[24] G. Wu and S. Yan, (2008) Prediction of mutations engi-
neered by randomness in H5N1 hemagglutinins of influ-
enza a virus, Amino Acid, 35, 365–373.
[25] G. Wu and S. Yan, (2008) Three sampling strategies to
predict mutations in H5N1 hemagglutinins from influ-
enza A virus. Protein Pept. Lett., 15, 731–738.
[26] Amino-acid pair predictability, (2009)
http://www.dreamscitech.com/Service/rationale.htm.
[27] Get Lat Lon, (2009) http://www.getlatlon.com/.
[28] S. Krauss, D. Walker, S. P. Pryor, L. Niles, L. Chenghong,
V. S. Hinshaw, and R. G. Webster, (2004) Influenza A vi-
ruses of migrating wild aquatic birds in North America,
Vector Borne Zoonotic. Dis., 4, 177–189.
[29] L. Z. Garamszegi and A. P. Møller, (2007) Prevalence of
avian influenza and host ecology, Proc. Biol. Sci., 274,
2003–2012.
[30] T. P. Weber & N. I. Stilianakis, (2007) Ecologic immu-
nology of avian influenza (H5N1) in migratory birds,
Emerg. Infect. Dis., 13, 1139–1143.
[31] A. Jahangir, Y. Watanabe, O. Chinen, S. Yamazaki, K.
Sakai, M. Okamura, M. Nakamura & K. Takehara, (2008)
Surveillance of avian influenza viruses in Northern pin-
tails (Anas acuta) in Tohoku District, Japan. Avian Dis.,
52, 49–53.
[32] M. Gilbert, J. Slingenbergh & X. Xiao, (2008) Climate
change and avian influenza, Rev. Sci. Tech., 27, 459–
466.
[33] Louchart, (2008) Emergence of long distance bird migra-
tions: A new model integrating global climate changes,
Naturwissenschaften, 95, 1109–1119.
[34] S. de La Rocque, J. A. Rioux & J. Slingenbergh, (2008)
Climate change: effects on animal disease systems and
implications for surveillance and control, Rev. Sci. Tech.,
27, 339–354.
[35] S. Morand & J. F. Guégan, (2008) How the biodiversity
sciences may aid biological tools and ecological engi-
neering to assess the impact of climatic changes, Rev. Sci.
Tech., 27, 355–366.
[36] E. A. Gould & S. Higgs, (2009) Impact of climate change
and other factors on emerging arbovirus diseases, Trans.
R. Soc. Trop. Med. Hyg., 103, 109–121.