Journal of Biomedical Science and Engineering
Vol.5 No.8(2012), Article ID:21461,7 pages DOI:10.4236/jbise.2012.58059
Gankyrin: A potential target for drug therapy against hepatocellular carcinoma
![]()
1Department of Biotechnology, University Institute of Engineering and Technology, Panjab University, Chandigarh, India
2Center for System Biology and Bioinformatics, U.I.E.A.S.T, Panjab University, Chandigarh, India
Email: *vpuri1825@yahoo.com
Received 14 May 2012; revised 2 July 2012; accepted 13 July 2012
Keywords: Hepatocellular Carcinoma; Murine Double Minute-2; p53; Tumor Suppressor Protein; Gankyrin
ABSTRACT
Hepatocellular Carcinoma is a primary malignant tumor of the liver and gankyrin is an oncoprotein over-expressed in hepatocellular carcinoma. It has been found that Gankyrin protein reduces the level of p53 protein by increasing its ubiquitylation and degradation, following a MDM-2 mediated pathway. Interaction of gankyrin with MDM2 enhances the ubiquitylation of p53. Independent study of this protein molecule revealed that it is identical to the p28 subunit of the 26S proteasome, having seven similar alpha helical ankyrin repeats. Gankyrin also binds to the Tumor Suppressor Protein (TSP) Retinoblastoma (RB), thereby accelerating its phosphorylation and proteasomal degradation. Blocking the expression of Gankyrin with MDM2 in cases of Hepatocellular Carcinoma (HCC) promoted apoptosis in cancer cells. Hence, Gankyrin can be used as a potential target for drug therapy against Hepatocellular Carcinoma.
1. INTRODUCTION
The most common type of cancer occurring in human beings, carcinoma, begins in the epithelial tissue, lining the inner and outer surfaces of the body. It is a form of cancer composed of cells that have developed the cytological appearance, histological architecture, or molecular characteristics of epithelial cells [1]. In multi-cellular organisms, cell growth is tightly regulated by check-points in the cell cycle that prevent over-proliferation of cells, enable repair of damaged DNA and selectively eliminate damaged cells by apoptosis. Hepatocellular Carcinoma, the most common primary malignant tumor of the liver, is of worldwide significance [2]. It may present itself with common symptoms like abdominal pain, clotting disorders resulting in easy bruising and bleeding, weight loss and jaundice. Detection of this carcinoma is difficult because the symptoms associated with it are no different than those related to a disease the patient may already be suffering. Hence, detection of this cancer at an early stage is very important. Over the past few years, several studies have been conducted to find an efficient detection technique for Hepatocellular Carcinoma. One of the techniques is to make use of a blood tumor marker, called alpha-fetoprotein (AFP) that has been known to detect HCC at an early stage. It is effective in spotting a small HCC but it has not been widely adopted because the cost-effectiveness of this technique is yet to be proven. Moreover the AFP levels in the body can also be elevated during pregnancy, which can be misleading.
There are mainly two tumor-suppressor proteins found in our body, Retinoblastoma (pRb) and p53. The retinoblastoma protein is associated with the check-point complex pRb-p16Ink4a-D cyclins whereas p53 is associated with the check-point complex p53-ARF-MDM2. The p53 tumor suppressor is a transcription factor that controls the expression of apoptotic proteins. The activity of p53 is regulated by proteins such as ubiquitin ligase MDM2 [3]. It is because of the activity of these protein molecules that the damaged cells lead to genomic instability. It has been found that gankyrin suppresses p53- dependent apoptosis in tumor cells. The ability of p53 to exert anti-proliferative effects, including cell cycle arrest and apoptosis, is strongly associated with its tumor suppressor function. The ability of p53 to bind DNA and activate transcription is required for p53 dependent growth arrest. P53 can mediate apoptosis by activating transcription of pro-apoptotic genes and repressing transcription of anti-apoptotic genes. p53 also controls apoptosis through transcription-independent mechanisms. Tight regulation of p53 activity is vital for maintaining normal cell growth and preventing tumorigenesis. The degradation of p53 is regulated by the ubiquitin-proteasome system. The 26S proteasome is a large multi-protein complex that selectively degrades proteins conjugated to ubiquitin. Foremost, ubiquitin is activated in an ATPdependent manner by an ubiquitin activating enzyme (E1). Then it is transferred to an ubiquitin conjugating enzyme (E2), and to an ubiquitin protein ligase (E3). For efficient polyubiquitylation, an additional conjugating enzyme (E4) is required.
MDM2, or the Murine double Minute 2, is a gene (as well as the protein which the gene encodes) which acts as a negative regulator of the p53 tumor suppressor [4]. It binds with p53 and represses its transcriptional activity. This is done when MDM2 binds to the N-terminal Trans —activation domain of p53. The transcription of MDM2 can be activated by p53, which means that if p53 is stabilized; the transcription of MDM2 is also induced, resulting in higher MDM2 protein levels. This is the reason why MDM2 is a p53 responsive gene. The full length transcript of MDM2 gene encodes a protein of 491 amino acids, with a predicted weight of 56kDa. At least 5% - 10% of all human tumors possess inappropriate over-expression of MDM2, which is an E3 ubiquitin ligase that negatively regulates p53.
Out of all the genes that have been found over expressed in Hepatocellular Carcinoma, two genes namely Ube2c and Gankyrin are of utmost importance. Ube2c gene, located on chromosome 20q13, belongs to the E2 gene family and codes for a 19.6 kDa protein involved in ubiquitin-dependent proteolysis [5]. Gankyrin was identified as the p28 component of the 26S proteasome [6-8]. Since gankyrin is involved in the ubiquitylation of tumor suppressor protein p53, gankyrin promises to be a potential target for drug therapy against liver cancer Hepatocellular Carcinoma. Gankyrin is identical to one of the component of the 26S proteasome. It contains an ankyrin repeat stack (6 repeats) with a 38-amino-acid N-terminal domain and the first letter “g” stands for “gann”, meaning cancer in Japanese [6]. It specifically interacts with the S6b ATPase of the 19S regulatory complex of 26S proteasome [7,8]. Gankyrin is highly conserved throughout evolution and is localized on human chromosome Xq22.3. Gankyrin expression is increased in all HCC’s compared with non-cancerous liver tissues [6]. Hence it was found that almost all HCC’s over-express this novel gene, gankyrin. Gankyrin binds to and potentiates the transcriptional activity of p53. It facilitates the binding of MDM2 to p53 and enhances ability of MDM2 to monoand poly-ubiquitylate p53. Further, gankyrin recruits a MDM2-p53 complex to the 26S proteasome and accelerates the degradation of p53 in an MDM2-dependent manner. In vitro, MDM2 catalyzes the addition of single ubiquitin moieties to a cluster of six C-terminal lysines in p53. MDM2 does not efficiently poly-ubiquitylate p53 under usual in vitro conditions, and the E4 activity of p300 is required in addition to MDM2 for p53 polyubiquitylation.
Viral oncoproteins such as HPV E7 and adenovirus E1A block the function of RB, which results in production of ARF (tumor suppressor), leading to stabilization of p53. The viruses counteract these cellular defenses by producing proteins that inhibit the function of p53. Interestingly, HPV E7, adenovirus E1A, and gankyrin all interact with ATPases of the 19S regulatory complex of the 26S proteasome in addition to their effects on RB. MAGE-A4, a member of the MAGE (melanoma antigen) family containing a well conserved region, binds to gankyrin and suppresses its oncogenic activity. Finally, gankyrin specifically binds to the C-terminus of the proteasomal S6 ATPase of the 26S proteasome. In HCC cells, down-regulation of gankyrin expression by RNAi promoted apoptosis and over-expression of MAGE-A4 suppresses activity of gankyrin [9-12]. Thus, gankyrin could be a potential target for cancer therapy. Blocking expression or function of gankyrin can be a valuable therapeutic or preventive strategy against human HCC’s. The main aim of this study is to computationally design a peptide that can capture MDM2 in a manner mimicking Gankyrin and can be used as a potential drug for the treatment of this disease.
2. MATERIALS AND METHODS
2.1. Homology Modeling
The first step towards successful in-Silico drug designing is 3D structure modeling. Homology modeling of a protein refers to constructing an atomic-resolution model of a target protein from its amino acid sequence and an experimental three dimensional structure of a related homologous protein. The basic principle of homology modeling lies in the fact that the protein structures, being few in numbers are more conserved as compared to the protein sequences. But a sequence identity of greater than 20% has to be achieved [13]. Different models of MDM2 were constructed using MODELLER 9.10. It is used for homology modeling (also known as comparative modeling) of three dimensional structures of proteins [14,15]. In this tool, an alignment file of a target sequence to be modeled with known related structures is provided and MODELLER automatically calculates a model containing all non-hydrogen atoms. MODELLER can give as many number of structures as desired by the user in two easy steps. The script is written in “python”, a general purpose, high level, programming language. The FASTA [16] sequence of the target MDM2 was obtained from protein database of NCBI (Accession Number Q00987.1 and Gene id 266516). The protein consists of 491 amino acids. Subsequently, the template structure 1Z1M selected with protein BLAST [17] of target sequence and template structure was aligned using MODELLER 9.10 and hence construction of ten rough 3D models was done. The constructed models were evaluated for geometry, stereochemistry checks and energy distribution on the basis of DOPE score and Ramachandran plot (using PROCHECK). DOPE score stands for Discrete Optimized Protein Energy score. It is a statistical potential used to assess homology models in proteins structure prediction. It can give us information about the conformation of the protein. Lesser the DOPE score value, higher is the structural stability of the protein molecule. The MODELLER evaluates the DOPE value of the protein structures. The structure (PDB) file of different models and a command python file are required to calculate this energy. The python script for constructing models comes under the file name “evaluateDOPE.py”. Now, since stability is inversely proportional to the system energy, the structure having the lowest DOPE value has highest structural stability. If there are some residues in the fourth quadrant, then structure refinement using loop modeling has to be done.
The second parameter used for the detection of most favorable structure is Ramachandran Plot, this is available on an online server SAVES, which stands for Structure Analysis and Verification server. Ramachandran plot is a way to visualize the backbone torsion angles of amino acids residues in a protein structure, hence accounting for the structural stability of the molecule. It is a part of PROCHECK, a tool present on the SAVE server that checks the stereo-chemical properties of the protein structures. The Ramachandran plots for all ten MDM2 structures were obtained. There are four quadrants in a normal Ramachandran Plot. The first belongs to right-handed α helix, second belongs to β-sheets, third belongs to left-handed α helix, and the fourth one is the disallowed region.
2.2. Active Site Prediction
Since only a few amino acids present in MDM2 actually play an important role in interaction with gankyrin, thus we open this molecule in LIGSITEcsc [18], a web-server for the automatic identification of pockets on the query protein sequence. The input file can either be uploaded directly by the user, or the PDB code of the file along with chain ID. LIGSITEcsc uses the concept of surfacesolvent-events and involves surface residues conservation degree. The binding site prediction was based on 1.5 Å grid space, 5 Å probe radius and number of binding sites to ten. The server searches for pockets on the surface of the protein molecule and it shows the interacting amino acids residues as an output. In the present study, LIGSITEcsc step gave us five pockets as output. Out of these five pockets, only one was found to be repetitive in the structure of the protein molecule, which also contained the gankyrin binding domain. The binding site residues are LYS 412, GLU 413, SER 425 and VAL 426.
2.3. Peptide Designing
For designing a peptide, a sequence with 15 amino acid residues was selected. The FASTA format sequence is selected in SPDB viewer [19] v4.0.4. Since the selected active site is a “loop”, we make sure that the designed peptide also has a loop at its tail end. To achieve this, we refer to the atom file of the structure file of MDM2. Keeping in mind that amino acids LYS 412, GLU 413, SER 425 and VAL 426 are responsible for interactions, we assign the amino acids present in our peptides the same φ, ψ and ω angles in SPDB viewer v4.0.4. Based on the three dimensional geometry of amino acids, we introduce two Proline and two Glycine molecules in the peptide. These amino acids will account for the helical shape of the designed peptide. The amino acids responsible for interaction at the pocket (active site) of MDM2 have been taken as terminal points, with Lysine and Glutamic Acid on one end, while Serine and Valine on the other. For the rest of the seven amino acids, Alanine is introduced. This is done to avoid steric hindrance among the amino acids of our peptide. Once this sequence was finalized, it was saved as a FASTA format sequence.
2.4. Visualization
Visualization of the peptide and protein structures was performed on PyMol v1.3 molecular graphics program, a comprehensive software package for rendering and animating 3D-structures. This software produces high quality three dimensional images of small molecules, proteins and nucleic acids.
3. RESULTS
Since an accurate structure was not available for the protein MDM2, homology modeling was used to generate 3D protein models of the desired protein i.e. MDM2 using a template structure with PDB code 1Z1M. The constructed 3D models were checked for DOPE score and Ramachandran plot respectively with Modeller 9.10 and PROCHECK. Results have been shown in Table 1. Model number 10 was chosen as the best model with parameters like Ramachandran plot (78.5% core, 15.7% allowed, 2.7% generously allowed and 3.1% disallowed) and DOPE score (-21745.01172). Figure 1 shows the Ramachandran plot analysis of the model selected. Model structure prepared for visualization by PyMol v1.3 is shown in Figure 2. Details of pockets present in the protein model are described in Table 2. Pocket chosen for ligand binding was pocket 17.
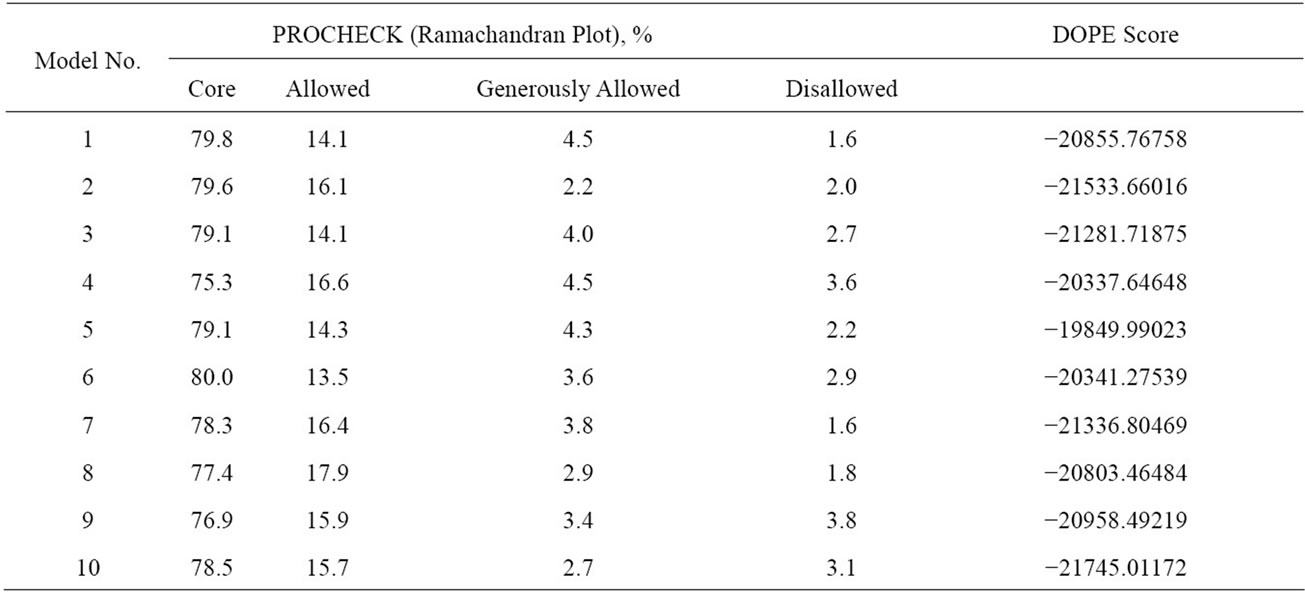
Table 1. analysis of generated models with procheck and modeler 9.10.

Figure 1. Ramachandran plot of selected MDM2 structure. Total number of residues: 491; residues in most favoured regions: 350 (78.5%); Residues in additional allowed regions: 70 (15.7%); Residues in generously allowed regions: 12 (2.7%); Residues in disallowed regions: 14 (3.1%); Number of non-glycine and non-proline residues 446; Number of end-residues (excl. Gly and Pro) 1; Number of glycine residues (shown as triangles) 21; Number of proline residues 23.

Figure 2. MDM2 favorable structure obtained from modeller v9.10.

Table 2. Binding pockets in the protein found by Ligsitecsc.
As the active site was discovered, a peptide was designed that was expected to simply fit into this active site, and then may lead to some conformational changes that prevent gankyrin molecule to bind with MDM2. If this happens, then the tumor suppressor p53 will start acting on the tumor cells and induce apoptosis in them, hence giving a potential therapy against Hepatocellular Carcinoma. For designing a peptide, a sequence having 15 amino acid residues was selected. Based on the knowledge of amino acids, we introduce two Prolines and two Glycine molecules in the peptide. The amino acids responsible for interaction at the pocket (active site) of MDM2 have been taken as terminal points, with Lysine and Glutamic Acid on one end, while Serine and Valine on the other. For the rest of the seven amino acids, Alanine is introduced.
This is done to avoid steric hindrance among the amino acids of our peptide. Once this sequence was finalized, it was saved as a FASTA format sequence. The sequence combination designed is “KEAAAGPGPAAAASV”. The PDB structure of the mentioned sequence has been shown in Figure 3.
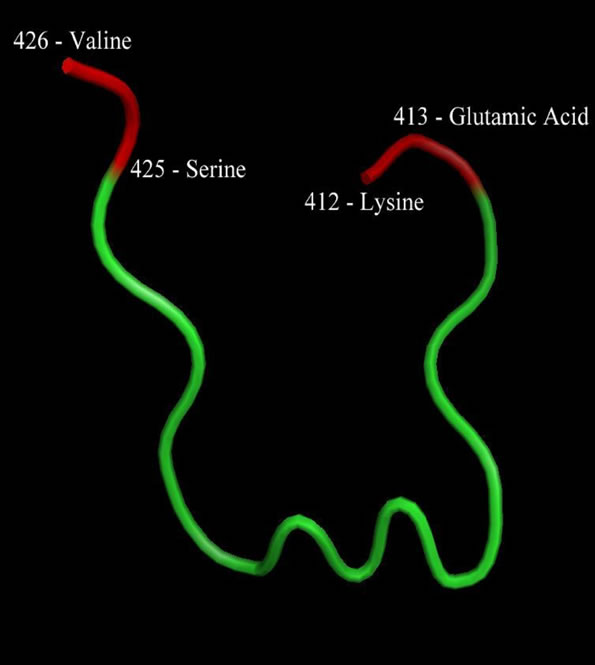
Figure 3. Structure of the designed peptide with highlighted terminal points.
4. DISCUSSION
In order to find out an effective treatment for hepatocellular carcinoma, many efforts have been made by different groups of people. But till now, there is no effective drug for this disease. The main idea behind identifying the structure of MDM2 was to study the interaction of Gankyrin with MDM2. Identification of the amino acid residues forming the active site on the surface of MDM2 can help us determine the interacting domain present in gankyrin molecule. For that, homology modeling has been done using a template structure and it was aligned with the sequence of MDM2.
We obtained ten similar structures for MDM2 as a result of homology modeling. Brief analysis of these models with Ramachandran plot and DOPE score was done to decide the stability in protein models.
More number of residues in disallowed region gives instability to the protein structure. To make a protein stable to configuration, it is necessary to remain all the amino acid residues in allowed region in the Ramachandran plot. In the selected model, least number of amino acids was present in disallowed region and the DOPE score was considerable relative to other models. The model number 10 was considerable as structure of MDM2 protein with 3.1% disallowed amino acid residues and a DOPE score of −21,745.01172.
The functional domains of MDM2, also known as ubiquitin protein ligase have been showed in Figure 4. The purple colored band shows the region where gankyrin binds with MDM2 resulting in its enhanced activity. The amino acid residues from 411 to 438 were involved in interaction with gankyrin.
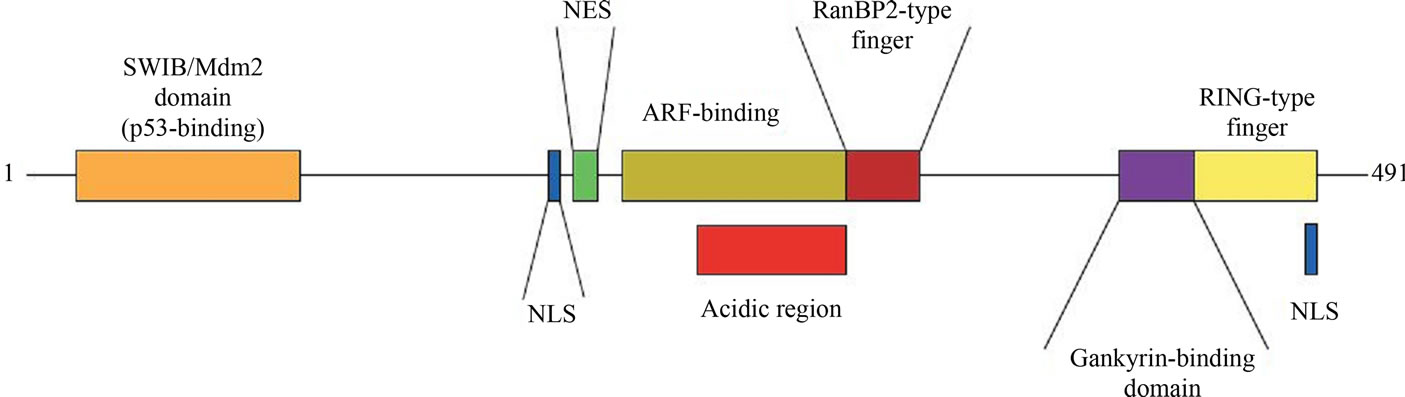
Figure 4. Functional domains of MDM2.

Figure 5. Delivering polyubiquitylated substrates to the 26S proteasome.
This region is just N-terminal to the RING (Really Interesting New Gene) domain. This puts the gankyrin binding region of MDM2 in a central position, as the N terminus of MDM2 binds p53 and the C terminus contains a RING site that enables association with an ubiquitin conjugating enzyme [20].
The de novo drug designing approach has been used to generate a peptide that may have the ability to bind itself at the “gankyrin binding domain” of MDM2, hence preventing the ubiquitylation of tumor suppressor p53. Figure 5 shows the mechanism by which ubiquitylated p53 is delivered to the proteasome by binding to MDM2 [9].
If the interaction between gankyrin and MDM2 is blocked, it might prevent the degradation of p53. Gankyrin plays a potential role in HCC. Therefore, the structure of gankyrin and MDM2 is important to begin the strategies of designing peptides that block their activities and provide a basis for drug development for therapeutic intervention in HCC.
5. CONCLUSION
Based on the various analyses performed in the present study, it may be concluded that the peptide generated in the study might have the capability of binding to the active site of MDM2, the N-terminal to the RING domain, hence preventing the interaction between gankyrin and MDM2. The same peptide can also be used to block the fifth ankyrin repeat present in gankyrin, which we know is responsible for the interaction between gankyrin and MDM2. This will result in the induction of apoptosis among cancer cells, and can be used as a potential drug against Hepatocellular Carcinoma.
REFERENCES
- Berman, J.J. (2004) Tumor taxanomy for the developmenttal classification of neoplasms. BMC Cancer, 4, 88-93. doi:10.1186/1471-2407-4-88.
- Thomas, M.B. (2005) Hepatocellular carcinoma: The need for progress. Journal of Clinical Oncology, 23, 2892-2899. doi:10.1200/JCO.2005.03.196.
- Quelle, D.E., Zindy, F., Ashmun, R.A. and Sherr, C.J. (1995) Alternative reading frames of the INK4a tumor suppressor gene encode two unrelated proteins capable of inducing cell cycle arrest. Cell, 83, 993-1000. doi:10.1016/0092-8674(95)90214-7.
- Iwakuma, T. and Lozano, G. (2003) MDM2, an introducetion. Molecular Cancer Research, 1, 993-1000. doi:10.1158/1541-7786..
- Ieta, K., Ojima, E., Tanaka, F., Nakamura, Y., Haraguchi, N., Mimori, K., Inoue, H., Kuwano, H. and Mori, M. (2007) Identification of overexpressed genes in hepatocellular carcinoma, with special reference to ubiquitinconjugating enzyme E2C gene expression. International Journal of Cancer, 121, 33-38. doi:10.1002/ijc.22605.
- Higashitsuji, H., Itoh, K., Nagao, T., Dawson, S., Nonoguchi, K., Kido, T., Mayer, R.J., Arii, S. and Fujita, J. (2000) Reduced stability of retinoblastoma protein by gankyrin, an oncogenic ankyrin-repeat protein overexpressed in hepatomas. Nature Medicine, 6, 96-99. doi:10.1038/71600
- Manjasetty, B.A., Quedenau, C., Sievert, V., Büssow, C., Niesen, F., Delbrück, H. and Heinemann, U. (2004) X-ray structure of human gankyrin, the product of a gene linked to hepatocellular carcinoma. PROTEINS: Structure, Function and Bioinformatics, 55, 214-217. doi:10.1002/prot.20028
- Hori, T., Kato, S., Saeki, M., DeMartino, G.N., Slaughter, C.A., Takeuchi, J., Toh-e, A. and Tanaka, K. (1998) cDNA cloning and functional analysis of p28 (Nas6p) and p40.5 (Nas7p), two novel regulatory subunits of the 26S proteasome. Gene, 216, 113-122. doi:10.1016/S0378-1119(98)00309-6
- Higashitsuji, H., Itoh, K., Sakurai, T., Nagao, T., Sumitomo, H., Masuda, T., Dawson, S., Shimada, Y., Mayer, R.J. and Fujita, J. (2005) The oncoprotein gankyrin binds to MDM2/HDM2, enhancing ubiquitylation and degradation of p53. Cancer Cell, 8, 75-87. doi:10.1016/j.ccr.2005.06.006
- Li, H., Fu, X., Chen, Y., Hong, Y., Tan, Y., Cao, H., Wu, M. and Wang, H. (2005) Use of adenovirus-delivered siRNA to target oncoprotein p28GANK in hepatocellular carcinoma. Gastroenterology, 128, 2029-2041. doi:10.1053/j.gastro.2005.03.001.
- Nagao, T., Higashitsuji, H., Nonoguchi, K., Sakurai, T., Dawson, S., Mayer, R.J., Itoh, K. and Fujita, J. (2003) MAGE-A4 interacts with the liver oncoprotein gankyrin and suppresses its tumorigenic activity. The Journal of Biological Chemistry, 278, 10668-10674. doi:10.1074/jbc.M206104200
- Sakurai, T., Itoh, K., Higashitsuji, H., Nagao, T., Nonoguchi, K., Chiba, T. and Fujita, J. (2004) A cleaved form of MAGE-A4 binds to Miz-1 and induces apoptosis in human cells. The Journal of Biological Chemistry, 279, 15505-15514. doi:10.1074/jbc.M310437200
- Chothia, C. and Lesk, A. M. (1986) The relation between the divergence of sequence and structure in proteins. The EMBO Journal, 5, 823-826. doi:10.1002/prot.22927/abstract.
- Eswar, N., Marti-Renom, M.A., Webb, B., Madhusudhan, M.S., Eramian, D., Shen, M., Pieper, U. and Sali, A, (2006) Comparative protein structure modeling with MODELLER. John Wiley & Sons, New York. doi:10.1002/0471140864.ps0209s50.
- Marti-Renom, M.A., Stuart, A., Fiser, A., Sánchez, R., Melo, F. and Sali, A. (2000) Comparative protein structure modeling of genes and genomes. Annual Review of Biophysics, 29, 291-325. doi:10.1146/annurev.biophys.29.1.291
- Lipman, D.J. and Pearson, W.R., (1985) Rapid and sensitive protein similarity searches. Science, 227, 1435-1441. doi:10.1126/science.2983426
- Altschul, S.F., Gish, W., Miller, W., Myers, E.W. and Lipman, D.J. (1990) Basic local alignment search tool. Journal of Molecular Biology, 215, 403-410. doi:10.1006/jmbi.1990.9999
- Bingding, H. and Schroeder, M. (2006) LIGSITEcsc: predicting protein binding sites using the Connolly surface and degree of conservation. BMC structural Biology, 6, 19. doi:10.1186/1472-6807-6-1
- Schwede, T., Diemand, A., Guex, N. and Peitsch, M.C. (2000) Protein structure computing in the genomic era. Research in Microbiology, 151, 107-112. doi:10.1016/S0923-2508(00)00121-2.
- Dawson, S., Higashitsuji, H., Wilkinson, A.J., Fujita, J. and John Mayer, R. (2006) Gankyrin: a new oncoprotein and regulator of pRb and p53. Trends in Cell Biology, 16, 229-233. doi:10.1016/j.tcb.2006.03.001
NOTES
*Corresponding author.