Food and Nutrition Sciences
Vol. 4 No. 9A1 (2013) , Article ID: 36120 , 6 pages DOI:10.4236/fns.2013.49A1008
Short Communication: Effect of Timing of Introduction to Pasture Post Calving and Supplementation with Saccharomyces cerevisiae on Milk Fatty Acid Profiles in Early Lactation Dairy Cows

1School of Agriculture and Food Science, University College Dublin, Dublin, Ireland; 2Arasco Feeds, Riyadh, KSA; 3Department of Animal Production, University of Buenos Aires, Buenos Aires City, Argentina; 4School of Veterinary Medicine, University College Dublin, Dublin, Ireland.
Email: stephen.c.whelan@ucd.ie
Copyright © 2013 Karina Mary Pierce et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received May 16th, 2013; revised June 16th, 2013; accepted June 24th, 2013
Keywords: Fatty Acid; Dairy Cow; Nutrition; Post-Partum
ABSTRACT
There is increased public awareness of the effect of dietary fatty acid (FA) profile on human health. Therefore, when devising nutritional management strategies for dairy cows it is important to evaluate the effects of said strategies on the FA profile of the milk. This experiment investigates the effects of two early PP nutritional management strategies (NM); abrupt introduction to pasture (AP) or a total mixed ration for 21 d followed by a gradual introduction to pasture over 7 d (GP), with (Y) or without (C) live yeast (YS) on milk fatty acid (FA) profile. Forty multiparous dairy cows were assigned to one of four dietary treatments in a two (AP vs. GP) by two (Y vs. C) factorial, randomized block design. The experiment was conducted from d 1 to 70 PP. Pasture, TMR and concentrate samples were taken weekly to assess the chemical and FA composition. Milk yield was recorded daily and individual milk samples were collected weekly to determine milk FA composition. There was no interaction between NM strategy and YS supplementation on milk FA. Similarly, YS supplementation did not affect milk FA profile. However, GP had higher concentrations of C10 (P = 0.04), C12 (P = 0.01), C14 (P = 0.02) and medium chain FA (P = 0.02) vs. AP. Whereas AP had higher concentrations of the FA cis-9, C18:1 (P < 0.01), long chain FA (P = 0.1) and unsaturated FA (P = 0.01) and lower concentrations of saturated FA (P = 0.01) vs. GP. These results suggest that abruptly introducing the early lactation dairy cow to a pasture based diets positively alters the FA composition of the milk produced when compared to the milk from a dairy cow gradually introduced to pasture.
1. Introduction
In recent years consumers have become increasingly aware of the potential health benefits from consuming dairy products rich in polyunsaturated fatty acids (PUFA), particularly those in the n-3 (omega-3) fatty acid (FA) group and the conjugated linoleic acid (CLA) isomer cis-9 trans-11 C18:2 (Ruminic acid, RA) [1]. Therefore, when devising nutritional management strategies for the early lactation dairy cow it is important to measure the effects of said strategies on the FA profile of the milk produced. Several factors are known to influence the FA composition of bovine milk including diet [2], rumen function [3], stage of lactation and body tissue mobilization [4]. In Ireland, early turn out to pasture post calving has been advocated as a means of maximizing milk production from grazed pasture, increasing the competitiveness of Irish dairy production systems [5]. In addition, the milk produced from pasture fed dairy cows has been reported to contain higher levels of CLA compared to cows offered total mixed ration (TMR) diets [2,6]. However, abrupt changes in the diet offered during early lactation can impair rumen function [7], depress dry matter intake (DMI) and exacerbate body tissue mobilization [8], thus altering the FA profile in milk.
Nutritional management (NM) strategies such as gradual introduction to pasture (GP) may ameliorate the negative effects of abrupt dietary changes [9]. However, the effect of this dietary strategy on milk FA profile is less well known. Supplementation with live yeast has also been suggested as a means of overcoming the risks associated with abrupt dietary change [10]. In addition, live yeast supplementation has been reported to alter rumen pH and volatile fatty acid (VFA) concentrations [11, 12]. However, there is a scarcity in literature reporting the effect of live yeast supplementation on milk FA profile.
The hypothesis of the current study is that abrupt turn out to pasture and supplementation with live yeast may improve the concentrations of CLA in the milk of early lactation dairy cows when compared to milk from dairy cows gradually introduced to pasture.
2. Materials and Methods
2.1. Animals and Management
All procedures involving the use of animals were conducted under experimental license from the Department of Health and Children, Ireland [13]. Forty multiparous dairy cows were selected from the dairy herd at UCD Lyons Research Farm, Dublin, Ireland (53˚17'56''N, 6˚32'18''W) and blocked by previous lactation 305 d milk yield, parity, BCS, and predicted calving date. Within each block, cows were randomly allocated to dietary treatments in a 2 × 2 factorial arrangement. Animals were either introduced to pasture gradually (GP, n = 20) or abruptly (AP, n = 20) post calving and were offered concentrates with (Y) or without (C) supplementary Yea-sacc1026 (YS) at a rate of 2.5 × 109 cfu of Saccharomyces cerevisiae, strain 1026 (Yea-sacc1026, Alltech, KY, United States of America). The experiment was conducted from d 0 to 70 postpartum (PP). From d 1 to 21 PP the GP group were housed in a free stall barn and offered 23 kg dry matter (DM)/day of a TMR containing on a DM basis; 27% maize silage, 16.5% grass silage, 3.5% wheat straw and 53% TMR concentrate with or without YS. Individual feed intake during this time was facilitated using computerized feed troughs (RIC System; Insentec B.V., Marknesse, The Netherlands). From d 21 to 28 PP, the GP group was allowed access to pasture between am and pm milking (0800 and 1530 h) and had access to the TMR between pm and am milking. During this time the GP groups were introduced to the pasture concentrates at an initial rate of 3.5 kg DM/d increasing by 0.5 kg DM/d until d 28. By d 28 the GP group were grazing full time with no supplementary TMR. Animals in the AP group were offered 20 kg DM/d of a perennial ryegrass based pasture plus 7 kg DM/d of a pasture concentrate with or without YS throughout the experiment.
2.2. Data and Sample Collection and Analysis
Samples of the TMR, pasture and concentrates were collected weekly and stored at −20˚C pending analysis. Dry matter, neutral detergent fiber (NDF), acid detergent fiber (ADF), crude protein (CP), gross energy, ether extract, starch and ash were determined as described in Whelan et al. [14]. The FA in feed was extracted using the one-step methylation procedure of Sukhija and Palmquist [15] as described in Palladino et al. [16]. Concentrations of individual FA were determined using gas chromatography (GC) (Varian CP-3800 GC; Varian Inc., Palo Alto, Canada) fitted with a 100 m capillary column with an internal diameter of 0.25 mm coated with 0.39 μm film (CP7420, Varian Inc.). Nitrogen was the carrier gas and the column flow was held at 2 ml/ min (Table 1). Cows were milked twice daily at 0700 and 1600 h. Milk output was recorded daily and sampled weekly using a milk metering system (Weighall, Dairymaster, Kerry, Ireland). Weekly milk samples were collected from consecutive am and pm milking and pooled according to output. Milk fat was extracted from these samples by centrifuging samples at 4˚C for 20 min at 978 × g, the cream was collected and stored at −20˚C pending analysis. Concentrations of individual milk FA were determined using GC (previously described) using the method of Christie [17] as described in Palladino et al. [16]. Data were checked for adherence to normal distribution and homogeneity of variance using histograms and formal statistical tests as part of the Univariate procedure of SAS [18]. The natural logarithmic transformations of C10:0, C12:0, C18:3 and saturated FA (SFA) were used to normalize data distribution since the preliminary analyses revealed that the distribution of values for these variables were positively skewed. Transformed data were used to calculate P-values. However, the corresponding least squares means and standard errors of the nontransformed data are presented in the results. Analysis of data was conducted using Proc Mixed of SAS [18] including terms for NM, YS supplementation, block, time and their interactions. Statistically significant differences between least squares means were tested using the PDIFF command incorporating the Tukey test for pairwise comparison of treatment means. Where interactions were not significant the term was excluded from the model. Statistical significance was assumed at a value of P < 0.05 and a tendency toward significance assumed at a value of P > 0.05 but <0.10.
3. Results and Discussion
There is considerable interest in developing dairy prod-
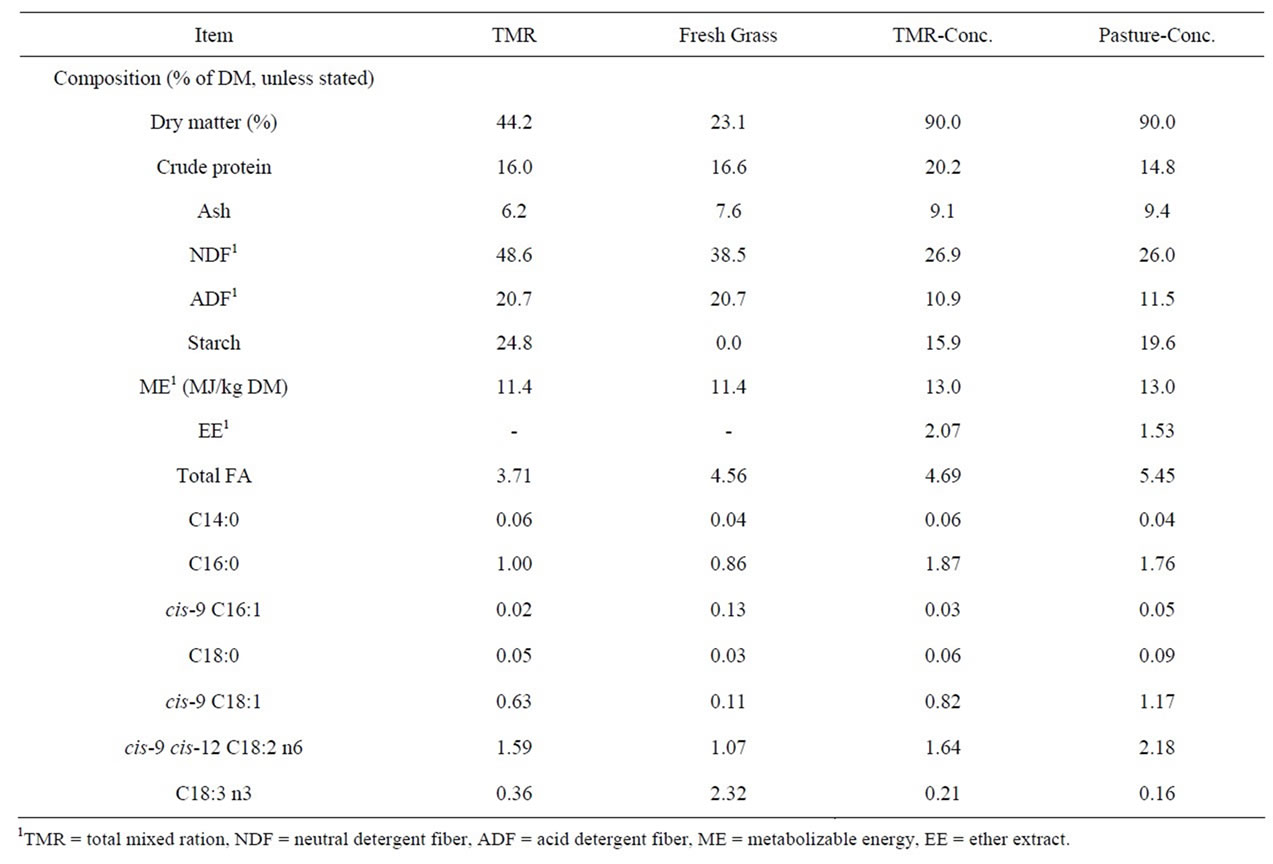
Table 1. Chemical and fatty acid composition of the total mixed ration (TMR), fresh perennial ryegrass, TMR-concentrate portion, and pasture-concentrate used as a supplement for grazing cows.
ucts rich in long chain fatty acids (LCFA) and CLA because of their beneficial effects on human health [17]. In the current study, milk produced by the GP group had higher concentrations of caprinic acid (C10, P = 0.04), laurinc acid (C12, P = 0.01), meristic acid (C14, P = 0.02) and medium chain FA (P = 0.02) than that of the AP group indicating a reduction in de-novo synthesis in pasture fed cows [2] (Table 2). In a companion study, Al Ibrahim et al. [18] reported a lower DMI (15.8 vs. 17.0 kg/d, P = 0.04) and energy intake (15.1 vs. 16.3 units of energy for lactation/d, P = 0.04) in the AP group versus the GP group. It is possible that this reduction in DMI reduced the supply of substrates and energy required for de-novo synthesis in the AP group as evidenced by the reduction in milk fat yield (1.09 vs. 1.17 kg/d, P = 0.08) and consistent with the observations of Kelly et al. [2] where a similar dietary regime was imposed. The hormonal status of an animal is also known to affect FA synthesis [6]. In the companion study, Al Ibrahim et al. [18] reported higher insulin concentrations in the GP group, thus contributing to increased production of C10:0 to C14:0 FA [19]. As the concentrations of C12:0 and C14:0 have been reported to contribute to plasma low density lipoprotein cholesterol levels, reducing the portion of these FA in bovine milk may have important human health implications [6].
Improving the concentrations of LCFA and CLA in bovine milk is also likely to benefit human health [17]. However, to improve the concentrations of LCFA and CLA in bovine milk the dairy cow must be offered diets that are also rich in LCFA as these FA cannot be synthesized de-novo [20]. The pasture offered in this experiment contained a higher portion of linolenic acid (C18:3) than the TMR diet. Additionally, pasture based diets have been reported to favor the proliferation of Butyrivibrio fibrisolvens in the rumen, a species responsible for the biohydrogenation of C18:3 to produce linoleic (C18:2) and oleic acid (C18:1) [21]. Analysis of the milk FA on a weekly basis shows an increase in concentrations of cis-9, C18:1 once the GP group was turned out to pasture (11.0 vs. 20.9% of milk FA for week 3 vs. week 4, P < 0.05). Thus, it is likely that a combination of increased dietary supply of C18:3 and rumen biohydrogenation contributed to the increased concentrations of cis-9, C18:1 (P < 0.01) in the milk of the AP versus the GP group.
Mobilization of body adipose tissue can contribute up
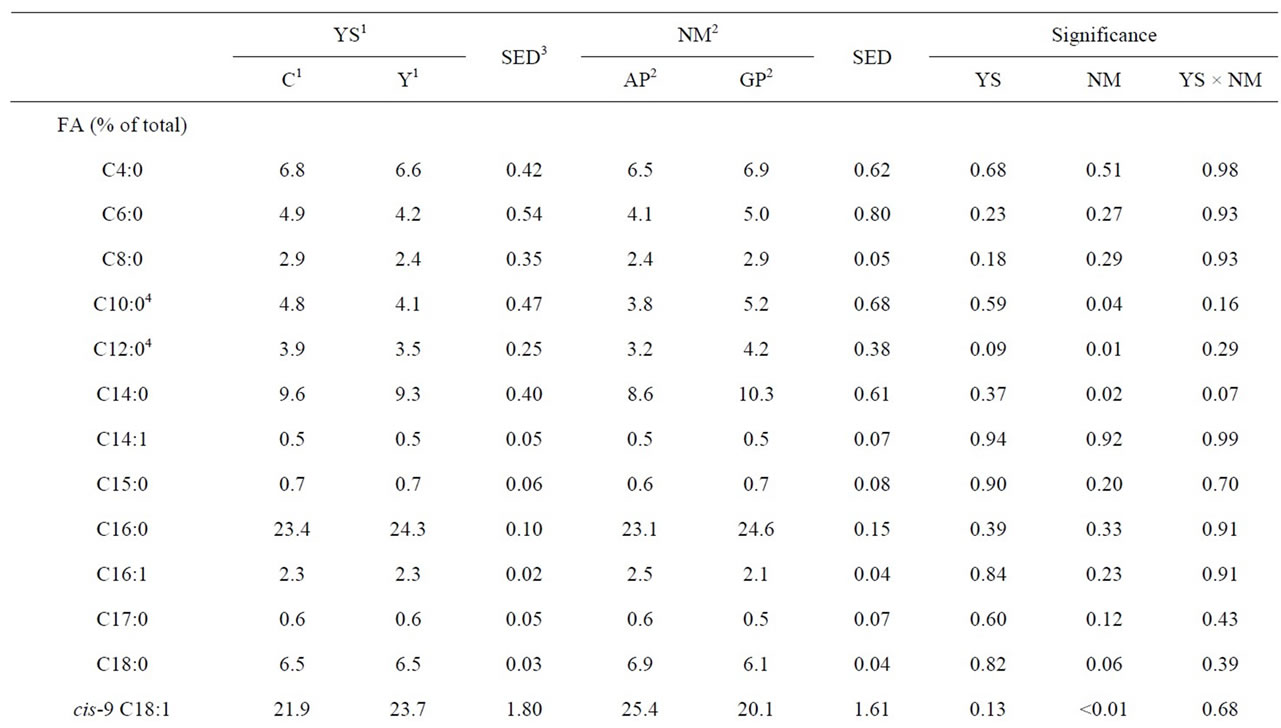
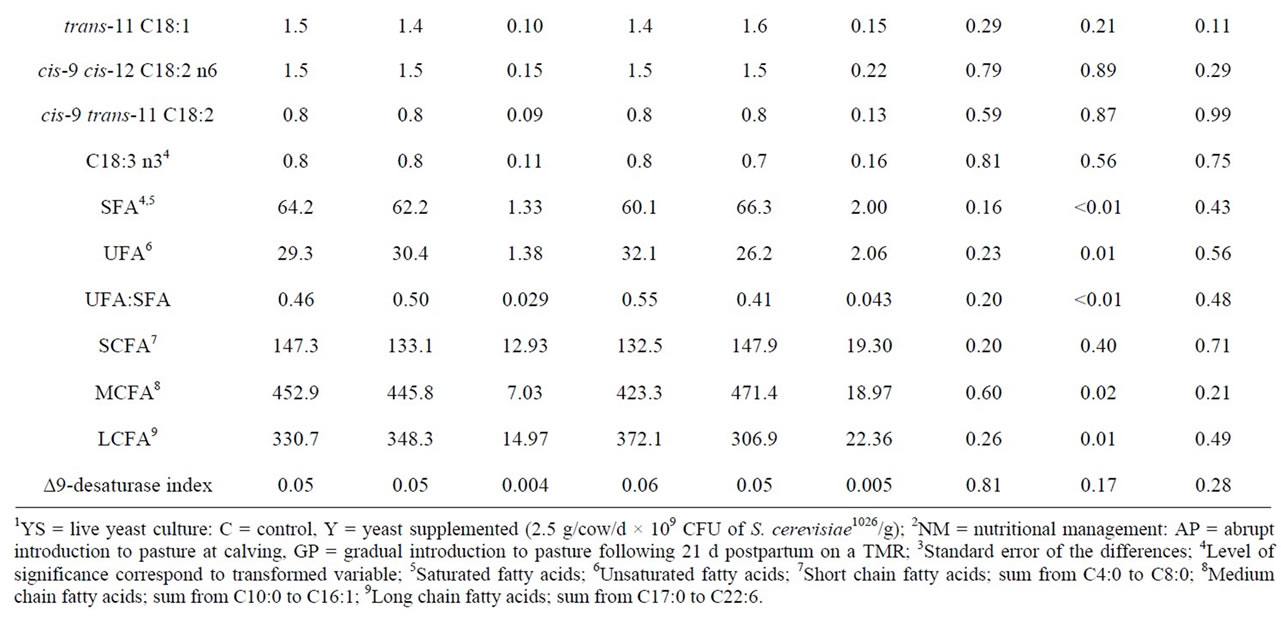
Table 2. Effect of live yeast culture (YS) supplementation and early postpartum nutritional management (NM) on milk fatty acid (FA) composition of early lactation dairy cows.
to 20% of FA in bovine milk during early lactation, particularly the LCFA [20]. In the companion study, Al Ibrahim et al. [18] reported concentrations of blood nonesterified FA (0.7 mmol/L) in the AP group that were indicative of negative energy balance, suggesting that part of the increase in LCFA in the milk from the AP group may have been a result of body tissue mobilization. This is important because managing the degree of body tissue mobilization experienced by the dairy cow is vital in maintaining the health of the dairy cow [8].
Lastly, a lower rumen pH has also been associated with increases in LCFA, unsaturated FA and the ratio of unsaturated FA to saturated FA [22]. In a companion study Al Ibrahim et al. [9] reported an increase in rumen pH in YS supplemented cows. However, there was no effect of YS supplementation on DMI or milk yield and it is likely that the differences in rumen pH was not sufficient to cause a change in milk FA profile as observed in this experiment.
4. Conclusion
The findings of the present study indicate that abruptly introducing animals to pasture can alter the FA profile of bovine milk toward a product with potential human health benefits.
5. Acknowledgements
The authors would like to acknowledge the contribution of the laboratory and farm staff of UCD Lyons Research Farm for their contribution to this work. This work was co-funded by Alltech as part of an Enterprise Ireland innovation partnership.
REFERENCES
- R. G. Jensen, “The Composition of Bovine Milk Lipids: January 1995 to December 2000,” Journal of Dairy Science, Vol. 85, No. 2, 2002, pp. 295-350. doi:10.3168/jds.S0022-0302(02)74079-4
- M. L. Kelly, et al., “Effect of Intake of Pasture on Concentrations of Conjugated Linoleic Acid in Milk of Lactating Cows,” Journal of Dairy Science, Vol. 81, No. 6, 1998, pp. 1630-1636. doi:10.3168/jds.S0022-0302(98)75730-3
- R. J. Dewhurst, et al., “Increasing the Concentrations of Beneficial Polyunsaturated Fatty Acids in Milk Produced by Dairy Cows in High-Forage Systems,” Animal Feed Science and Technology, Vol. 131, No. 3-4, 2006, pp. 168-206. doi:10.1016/j.anifeedsci.2006.04.016
- P. C. Garnsworthy, et al., “Variation of Milk Citrate with Stage of Lactation and De Novo Fatty Acid Synthesis in Dairy Cows,” Journal of Dairy Science, Vol. 89, No. 5, 2006, pp. 1604-1612. doi:10.3168/jds.S0022-0302(06)72227-5
- P. A. T. Dillon, et al., “Future Outlook for the Irish Dairy Industry: A Study of International Competitiveness, Influence of International Trade Reform and Requirement for Change,” International Journal of Dairy Technology, Vol. 61, No. 1, 2008, pp. 16-29. doi:10.1111/j.1471-0307.2008.00374.x
- A. L. Lock and P. C. Garnsworthy, “Seasonal Variation in Milk Conjugated Linoleic Acid and Δ9-Desaturase Activity in Dairy Cows,” Livestock Production Science, Vol. 79, No. 1, 2003, pp. 47-59. doi:10.1016/S0301-6226(02)00118-5
- G. A. Donovan, et al., “Influence of Transition Diets on Occurrence of Subclinical Laminitis in Holstein Dairy Cows,” Journal of Dairy Science, Vol. 87, No. 1, 2004, pp. 73-84. doi:10.3168/jds.S0022-0302(04)73144-6
- F. J. Mulligan and M. L. Doherty, “Production Diseases of the Transition Cow,” The Veterinary Journal, Vol. 176, No. 1, 2008, pp. 3-9. doi:10.1016/j.tvjl.2007.12.018
- R. M. Al Ibrahim, et al., “The Effect of Abrupt or Gradual Introduction to Pasture after Calving and Supplementation with Saccharomyces cerevisiae (Strain 1026) on Ruminal pH and Fermentation in Early Lactation Dairy Cows,” Animal Feed Science and Technology, Vol. 178, No. 1-2, 2012, pp. 40-47. doi:10.1016/j.anifeedsci.2012.09.011
- R. M. Al Ibrahim, et al., “The Effect of Nutritional Management and Supplementation with Saccharomyces cerevisiae on Ruminal pH and Fermentation in Early Lactation Dairy Cows,” Animal Feed Science and Technology, 2012 (accecpted).
- C. M. Guedes, et al., “Effects of a Saccharomyces cerevisiae Yeast on Ruminal Fermentation and Fibre Degradation of Maize Silages in Cows,” Animal Feed Science and Technology, Vol. 145, No. 1-4, 2008, pp. 27-40. doi:10.1016/j.anifeedsci.2007.06.037
- M. Thrune, et al., “Effects of Saccharomyces cerevisiae on Ruminal pH and Microbial Fermentation in Dairy Cows: Yeast Supplementation on Rumen Fermentation,” Livestock Science, Vol. 124, No. 1-3, 2009, pp. 261-265. doi:10.1016/j.livsci.2009.02.007
- S.I.613, “Cruelty to Animals Act 1876 (as Amended by European Communities. Regulations 2002 and 2005),” Statutory Instruments No. 613 of 2005, Department of Health and Children, Dublin, 2005.
- S. J. Whelan, et al., “Effect of Forage Source and a Supplementary Methionine Hydroxy Analogue on Nitrogen Balance in Lactating Dairy Cows Offered a Low Crude Protein Diet,” Journal of Dairy Science, Vol. 94, No. 10, 2011, pp. 5080-5089. doi:10.3168/jds.2011-4174
- P. S. Sukhija and D. L. Palmquist, “Rapid Method for Determination of Total Fatty Acid Content and Composition of Feedstuffs and Feces,” Journal of Agricultural and Food Chemistry, Vol. 36, No. 6, 1988, pp. 1202- 1206. doi:10.1021/jf00084a019
- R. A. Palladino, et al., “Fatty Acid Intake and Milk Fatty Acid Composition of Holstein Dairy Cows under Different Grazing Strategies: Herbage Mass and Daily Herbage Allowance,” Journal of Dairy Science, Vol. 92, No. 10, 2009, pp. 5212-5223. doi:10.3168/jds.2009-2404
- A. Lock and D. Bauman, “Modifying Milk Fat Composition of Dairy Cows to Enhance Fatty Acids Beneficial to Human Health,” Lipids, Vol. 39, No. 12, 2004, pp. 1197-1206. doi:10.1007/s11745-004-1348-6
- R. M. Al Ibrahim, et al., “Effect of Timing of PostPartum Introduction to Pasture and Supplementation with Saccharomyces cerevisiae on Milk Production, Metabolic Status, Energy Balance and Some Reproductive Parameters in Early Lactation Dairy Cows,” Journal of Animal Physiology and Animal Nutrition, Vol. 97, 2013, pp. 105-114. doi:10.1111/jpn.12048
- D. Yin, M. J. Griffin and T. D. Etherton, “Analysis of the Signal Pathways Involved in the Regulation of Fatty Acid Synthase Gene Expression by Insulin and Somatotropin,” Journal of Animal Science, Vol. 79, No. 5, 2001, pp. 1194-200.
- D. E. Bauman and J. M. Griinari, “Nutritional Regulation of Milk Fat Synthesis,” Annual Review of Nutrition, Vol. 23, No. 1, 2003, pp. 203-227. doi:10.1146/annurev.nutr.23.011702.073408
- L. Biondi, et al., “Changes in Ewe Milk Fatty Acids Following Turning Out to Pasture,” Small Ruminant Research, Vol. 75, No. 1, 2008, pp. 17-23. doi:10.1016/j.smallrumres.2007.07.004
- Y. Chilliard, et al., “Diet, Rumen Biohydrogenation and Nutritional Quality of Cow and Goat Milk Fat,” European Journal of Lipid Science and Technology, Vol. 109, No. 8, 2007, pp. 828-855. doi:10.1002/ejlt.200700080