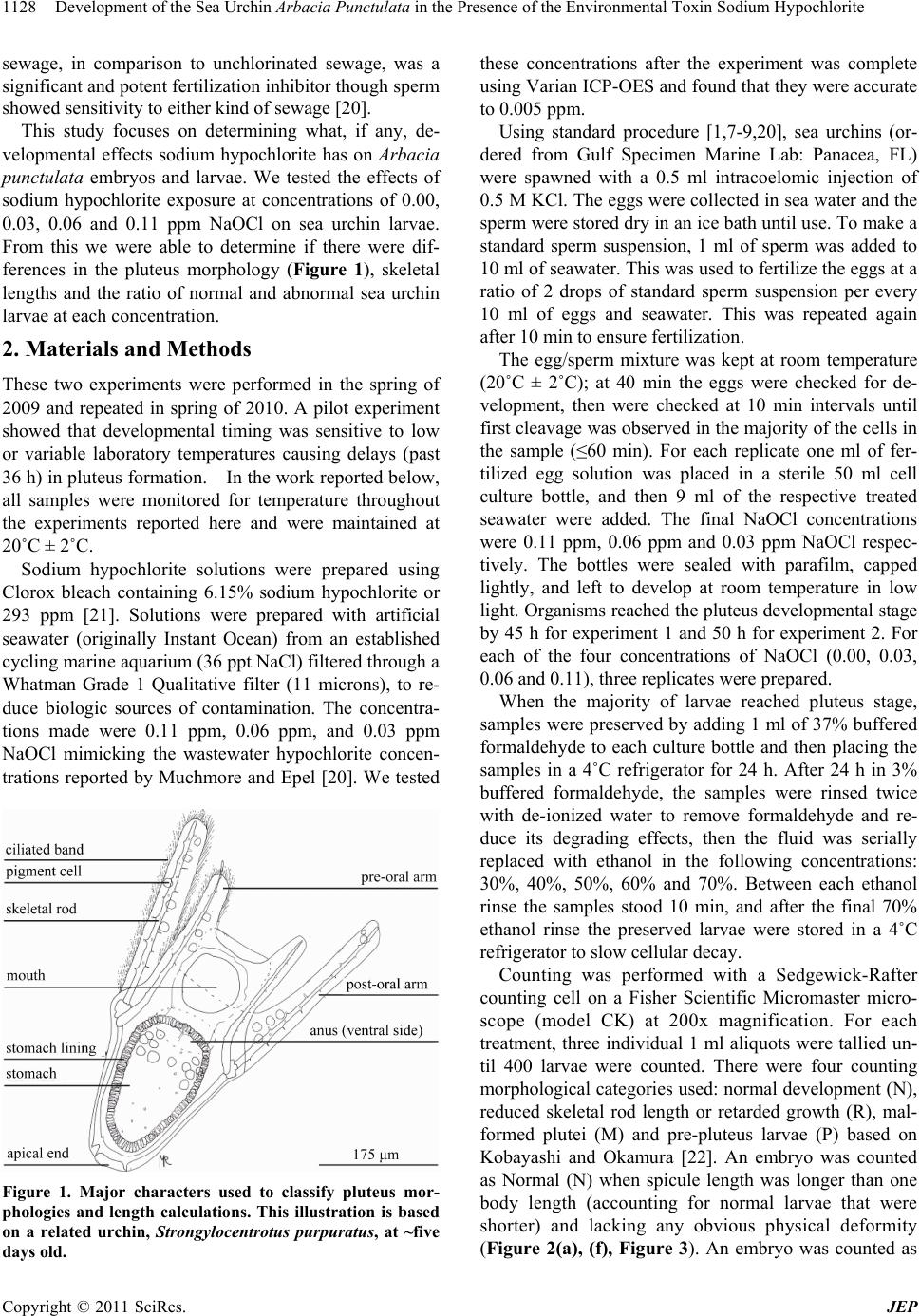
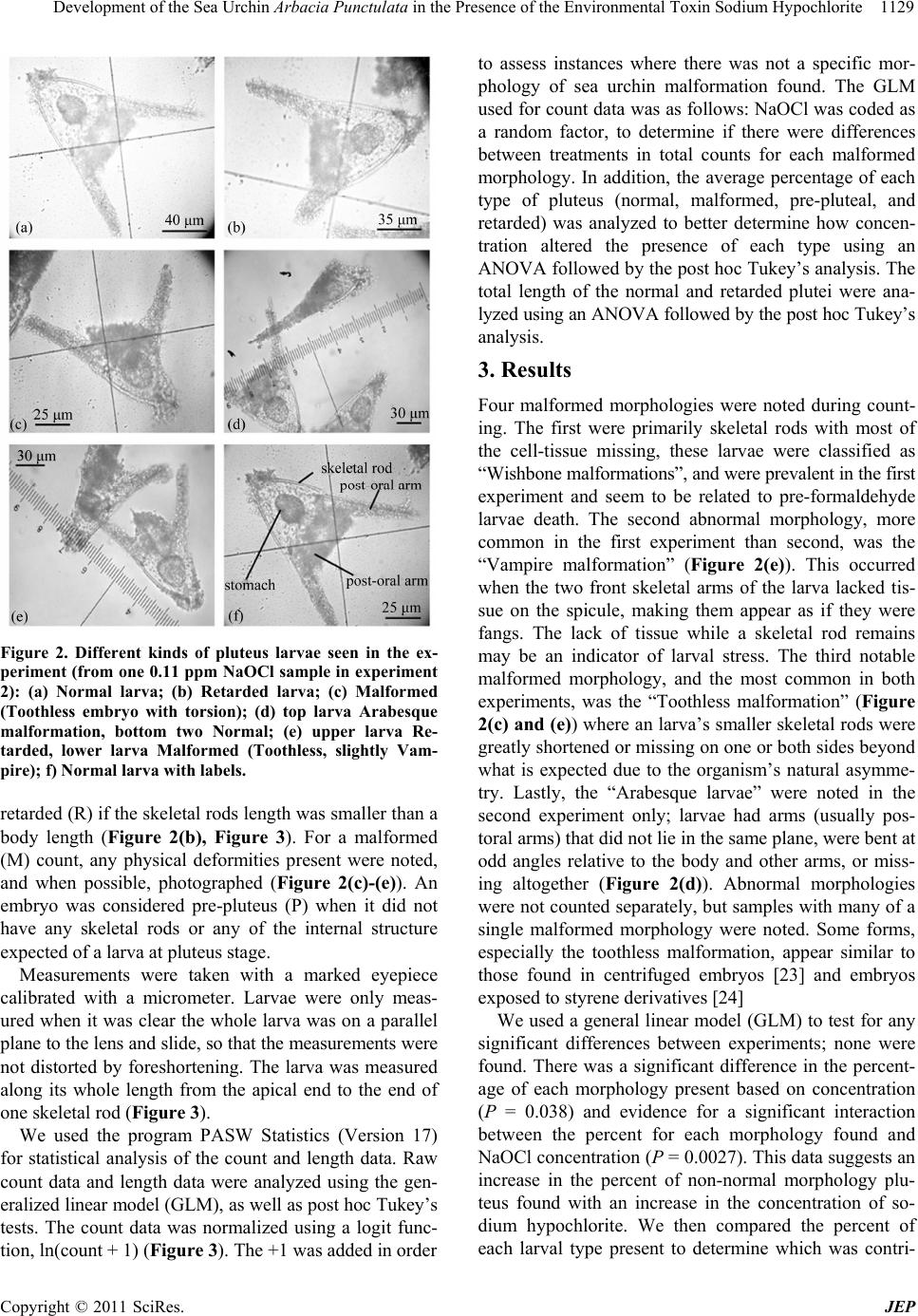
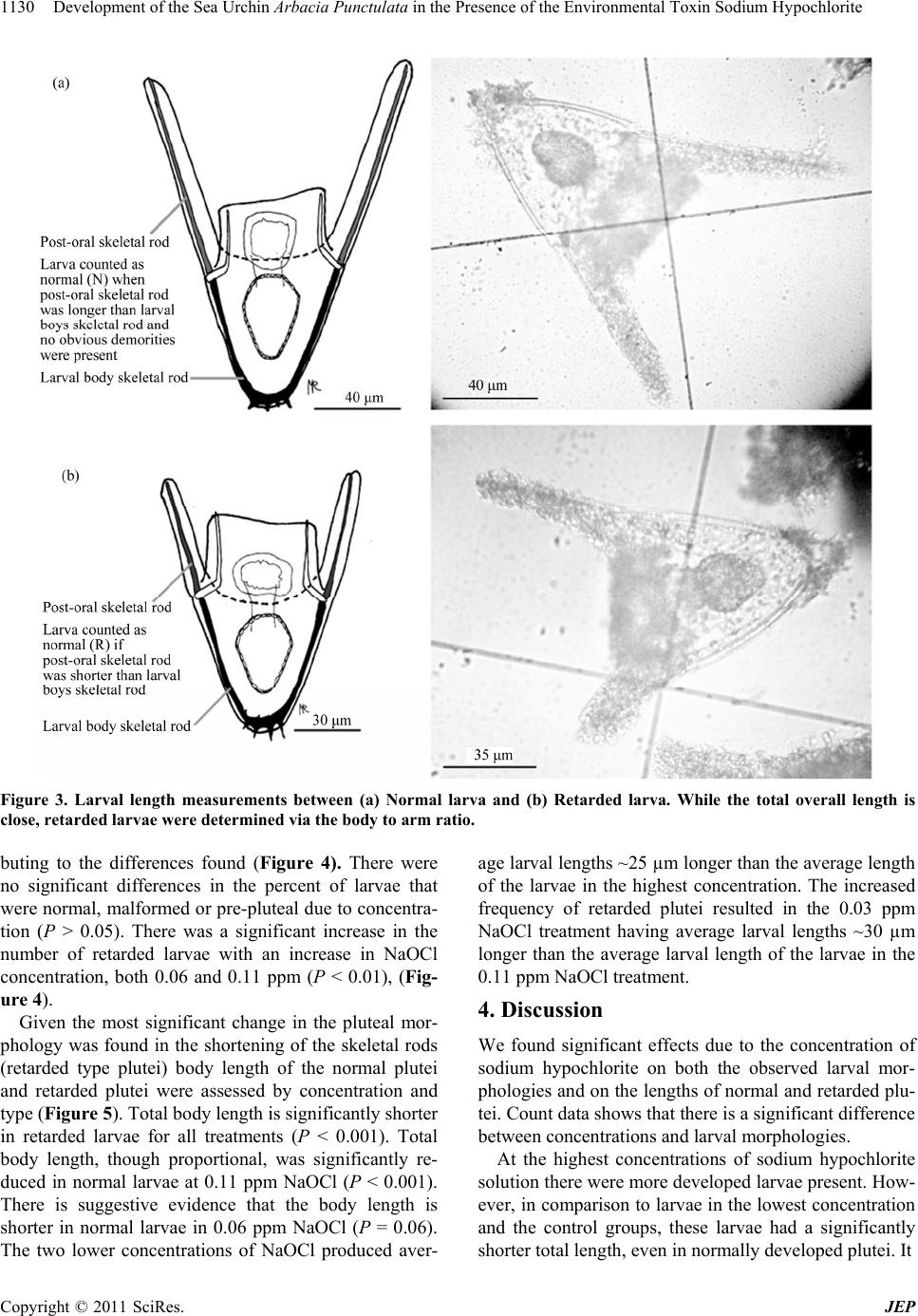
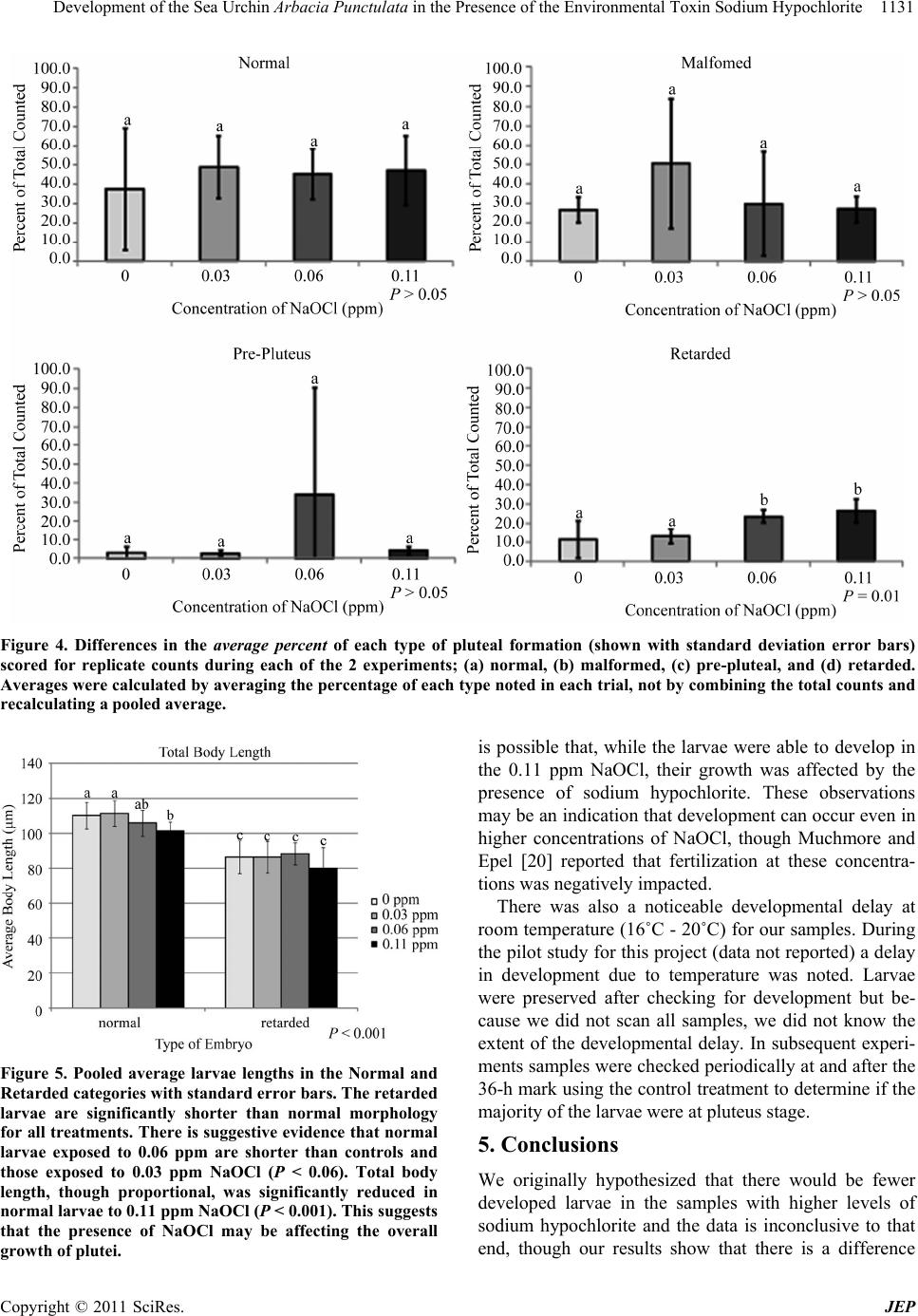
Development of the Sea Urchin Arbacia Punctulata in the Presence of the Environmental Toxin Sodium Hypochlorite
1132
between samples by concentration levels, especially at
the highest concentrations of NaOCl. At higher levels of
NaOCl exposure, A. punctulata larvae have shorter
skeletal lengths than those larvae in lower levels of
NaOCl. The significant increase in retarded type plutei
exposed to NaOCl and overall shortening of body length
even in normal type plutei, suggests that NaOCl may be
impacting the development of the skeletal rods. Further
investigation into the physiological mechanisms behind
this finding needs to be addressed. Development may be
slowed by the presence of NaOCl as noted by the in-
creased number of pre-pluteus larvae present in the 0.11
ppm NaOCl treatment. Further research is necessary to
determine the impact NaOCl has on observed larval
morphologies.
6. Acknowledgements
This study would not have been possible without the help
of many people. Very special thanks to the Science and
Math department at Columbia College Chicago for their
continued financial support and encouragement through-
out the course of this experiment, in particular G. Adams,
B. Budy, O. Carnate, D. Jordan, M. & R. Rock, and M.
Welsh.
REFERENCES
[1] P. A. Dinnel, J. M. Link, Q. J. Stober, M. W. Letourneau
and W. E. Roberts, “Comparitive Sensitivity of Sea Ur-
chin Sperm Bioassays to Metals and Pesticides,” Archive
of Environmental Contamination Toxicology, Vol. 18, No.
5, 1989, pp. 748-755.doi:10.1007/BF01225012
[2] A. V. Ghiradini, A. A. Novelli, C. Losso and P. F. Ghetti,
“Sea Urchin Toxicity Bioassays for Sediment Quality
Assessment in the Lagoon of Venice (Italy),” Chem. and
Ecol, Vol. 19, No. 2-3, 2003, pp. 99-111.
doi:10.1080/0275754031000119870
[3] A.V. Ghiradini, A.A. Novelli, C. Losso and P.F. Ghetti,
“Sperm Cell and Embryo Toxicity Tests Using the Sea
Urchin Paracentrotus Lividus (LmK)”, In: K.O. Gary, Ed.,
Techniques in Aquatic Toxicology, Taylor & Francis,
New York., 2005. pp. 147-168.
doi:10.1201/9780203501597.ch8
[4] D. Nacci, E. Jackin and R. Walsh, “Comparative Evalua-
tion of Three Rapid Marine Toxicity Tests: Sea Urchin
Early Embryo Growth Test, Sea Urchin Sperm Cell Tox-
icity Test and Microtox,” Environmental Toxicology and
Chemistry, Vol. 5, No. 6, 1986, pp. 521-525.
doi:10.1002/etc.5620050603
[5] G. Pagano, et al., “The Sea Urchin: Bioassay for the As-
sessment of Damage from Environmental Contaminants,”
In: J. Cairns Jr., Ed., Community Toxicology Testing,
ASTM, Philadelphia, 1986. pp. 66-92.
doi:10.1520/STP23050S
[6] R. T. Hinegardner, “Growth and Development of the
Laboratory Cultured Sea Urchin,” Biological Bulletin,
Vol. 137, 1969, pp. 465-475.
[7] EPA, “Purple Urchin, Strongylocentrotus Purpuratus and
Sand Dollar, Dendraster Excentricus Larval Development
Test Method,” In: G. A. Chapman, D. L. Denton and J. M.
Lazorchak, Eds., Short-Term Methods for Estimating the
Chronic Toxicity of Effluents and Receiving Waters to
West Coast Marine and Estuarine Organisms EPA/600/
R-95-136, U.S. Environmental Protection Agency, Cin-
cinnati, 1995.
[8] R. T. Hinegardner, “Care and Handling of Sea Urchin
Eggs, Embryos and Adults (Principally North American
species),” In: G. Czihak, Ed., The Sea Urchin Embryo:
Biochemistry and Morphogenesis, Springer-Verglag,
New York, 1975. pp. 10-25.
[9] G. Radenac, D. Fichet and P. Miramand, “Bioaccumula-
tion and Toxicity of Four Dissolved Metals in Paracen-
trotus Lividus Sea-Urchin Embryo,” Marine Environ-
mental Research , Vol. 51, No. 2, 2001, pp. 151-166.
doi:10.1016/S0141-1136(00)00092-1
[10] G. Rosen, I. Rivera-Duarte, D.B. Chadwick, A. Ryan,
R.C. Santore and P.R. Paquin, “Critical tissue copper
residues for marine bivalve (Mytilus galloprovincialis)
and echinoderm (Strongylocentrotus purpuratus) embry-
onic development: conceptual, regulatory and environ-
mental implications”. Marine Environmental Research,
Vol. 66, 2008, pp. 327-336.
doi:10.1016/j.marenvres.2008.05.006
[11] G. M. Mateyko, “Developmental Modifications in Arba-
cia Punctulata by Various Metabolic Substances,” Bio-
logical Bulletin, Vol. 133, No. 1, 1967, pp. 184-228.
doi:10.2307/1539802
[12] M. G. Marin, V. Moshino, F. Cima and C. Celli, “Em-
bryotoxicity of Butyltin Compounds to the Sea Urchin
Paracentrotus Lividus,” Marine Environmental Research,
Vol. 50, 2000, pp. 231-235.
doi:10.1016/S0141-1136(00)00072-6
[13] J. Runnstrom, “An Analysis of the Action of Lithium of
Sea Urchin Development,” Biological Bulletin, Vol. 68,
No. 3, 1935, pp. 378-384. doi:10.2307/1537560
[14] J. V. Goldstone, et al., “The Chemical Defensome: Envi-
ronmental Sensing and Response Genes in the Strongy-
locentrotus Purpuratus Genome,” Developmental Biology,
Vol. 300, No. 1, 2006, pp. 366-384.
doi:10.1016/j.ydbio.2006.08.066
[15] J. E. Hose, “Potential Uses of Sea Urchin Embryos for
Identifying Toxic Chemicals: Description of a Bioassay
Incorporating Cytologic, Cytogenetic and Embryologic
Endpoints,” Journal of Applied Toxicology, Vol. 5, No. 4,
1985, pp. 245-254. doi:10.1002/jat.2550050406
[16] M. A. Lewis, C. B. Daniels, J. C. Moore and T. Chen,
“Potential Genotoxicity of Wastewater-Contaminated
Pore Waters with Comparison to Sediment Toxicity and
Macrobenthic Community Composition,” Environmental
Toxicology, Vol. 17, No. 1, 2002, pp. 63-73.
doi:10.1002/tox.10033
[17] E. Emmanuel, G. Keck, J. Blanchard, P. Vermande and Y.
Perrodin, “Toxicological Effects of Disinfections Using
Sodium Hypochlorite on Aquatic Organisms and Its Con-
C
opyright © 2011 SciRes. JEP