Open Access Library Journal
Vol.04 No.06(2017), Article ID:76998,7 pages
10.4236/oalib.1103688
Clockwise Loops and Anticlockwise Loops Observed in a Stock-Recruitment Relationship
Kei Tanaka, Naoki Suzuki, Kazumi Sakuramoto*
Department of Ocean Science and Technology, Tokyo University of Marine Science and Technology, Tokyo, Japan

Copyright © 2017 by authors and Open Access Library Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: May 24, 2017; Accepted: June 16, 2017; Published: June 19, 2017
ABSTRACT
Loop-shaped trajectories are commonly observed in the stock recruitment relationship (SRR). A mechanism has been proposed that explains why the loops emerge in SRR. The aim of this study is to validate the mechanism using data for the stocks of 24 fish species that live in the waters around Japan. The following three questions are examined: 1) whether or not loop shapes are detected in SRRs for all 24 stocks; 2) whether or not the direction of the loops, i.e., clockwise or anticlockwise, changes depending on the age at maturity; and 3) whether or not the slope of the regression line adapted for SRR changes depending on the age at maturity. The results were as follows: 1) loop shapes in SRR were recognized for all 24 stocks analyzed in this study; 2) clockwise loops were dominant when the age at maturity was low, and anticlockwise loops were dominant when the age at maturity was high; 3) the slope of the regression line adapted for SRR has a negative relationship to the age at maturity. When the age at maturity was low, the slope was positive but less than unity. When the age at maturity was high, the slope had no trend and was recognized as statistically zero. These findings will drastically change the concept of SRR. All previous findings obtained by analyzing the SRR may be better to revise essentially.
Subject Areas:
Marine Biology
Keywords:
Stock Recruitment Relationship, Clockwise Loop, Anticlockwise Loop, Age at Maturity, Japanese Sardine, Japanese Anchovy, Chub Mackerel

1. Introduction
One of the most important tasks in fisheries resource management is to elucidate the fluctuation mechanisms in fish populations. One of the key factors in those mechanisms is the stock-recruitment relationship (SRR). Scientists in the field widely believe that a density-dependent effect is also an important factor in controlling population fluctuations [1] [2] . Typical, traditional SRR models include the well-known Ricker [3] and Beverton and Holt [4] models, which are based on a density-dependent mechanism in SRR. Recently, however, the importance of environmental factors has been reported in many studies. For instance, Su et al. [5] noted a significant positive effect of sea-surface temperature (SST) on the survival rate of northern pink salmon stocks; conversely, they observed a weak negative effect of SST on the survival rate of southern pink salmon stocks. Sundby [6] noted that the recruitment of Atlantic cod stocks showed the different responses to the temperature changes. The temperature influences the recruitment processes in a large number of ways; partly directly on vital rates in cod, and partly indirectly through trophic transfer. Chen and Irvine [7] proposed a new model in which environmental factors were incorporated and discussed the stock-recruitment relationship using fisheries data of southeast Alaska pink salmon (Oncorhynchus gorbuscha) and West Coast Vancouver herring (Clupea harengus). They concluded that the model proposed by Chen and Irvine [7] performed better than the traditional Ricker model and a Ricker model that was extended to include environmental effects.
Recently, Sakuramoto proposed a new concept of SRR mechanisms [8] - [13] and showed the mechanisms that explained clockwise or anticlockwise loops emerged in the SRR, using a simulation study. That is, when recruitment, R, fluctuates cyclically in response to environmental factors, and the spawning stock biomass (SSB) also fluctuates cyclically with a time lag (mainly determined by age at maturity), the SRR shows a clockwise loop or an anticlockwise loop for each period of environmental change. The former occurs when age at maturity is less than half of the environmental cycle, and the latter occurs when age at maturity is more than half of the environmental cycle.
Sakuramoto [12] also discussed the slope of the regression line for the plot of ln(R) against ln(SSB). When the age at maturity is low enough relative to the length of the cycle of environmental factors, the slope of the regression line is high and close to unity. However, the higher the age at maturity becomes, the more the slope of the regression line decreases. When the age at maturity becomes greater than approximately half the length of the cycle of environmental factors, the slope decreases to almost zero. As a result, the relationship between R and SSB is masked and cannot be detected. Furthermore, when the age at maturity becomes greater than half the length of the cycle of environmental factors, the slope of the regression line becomes negative.
Sakuramoto [12] analyzed the Pacific stock of Japanese sardines and Pacific bluefin tuna and showed that, for the former, the slope of the regression line was close to unity, and three clockwise loops appeared. In contrast, for the latter stock, the slope of the regression line had no significant slope and showed anticlockwise loops. Sakuramoto concluded that no relationship between R and SSB for Pacific bluefin tuna could be detected could be explained by the mechanism mentioned above.
The aim of this study is to elucidate whether or not this concept can be applied to the stocks of 24 fish species that live in the waters around Japan. That is, we examine: 1) whether or not loops are detected in SRRs for all 24 stocks; and 2) whether or not the direction of the loops, i.e., clockwise or anticlockwise, changes depending on the age at maturity; and 3) whether or not the slope of the regression line adapted for SRR changes depending on the age at maturity.
2. Materials and Methods
2.1. Data
The data used in this study were provided by the Fisheries Agency and Fisheries Research Agency Japan [14] , and included: number of recruitments; spawning stock biomass; and age at maturity of fish. The R and SSB for snow crab was estimated by area density method. The R and SSB for other 23 stocks are all estimated by VPA of which method dose not assume any stock-recruitment relationship. The species names and stocks names are listed in Table 1.
2.2. A Mechanism Produces the Loops
Sakuramoto [12] used four simulation models to reproduce the SRR observed in the Pacific stock of Japanese sardine and Pacific bluefin tuna. Using the same models proposed by Sakuramoto [12] , we explain the mechanism that loops are necessary detected in SRR. The basic model of SRR is expressed by Equation (1).
(1)
where Rt, St−1 and f(.) denote the recruitment in year t, spawning stock biomass in year t − 1, and a function that evaluates the effects of environmental factors in year t. The vector is a list of environmental factors that affect the strength in R, which comprised not only of physical factors such as water temperature, but also biological interactions such as prey-predator relationships. Parameters α and k denote a proportional constant and the number of environmental factors, respectively. That is, equation (1) implies that Rt is proportionally determined by St−1, and simultaneously, Rt is affected by environmental factors in year t.
Model 1 is the case when environmental effects can be neglected. That is, f(xt) in Equation (1) can be assumed to be unity. That is,
. (2)
where α denotes the recruitment per spawning stock biomass (RPS). The survival process is expressed by
(3)
For simplicity, m denotes the age at maturity and longevity of the fish. That is, fish reach maturity age at m-year old, then, they spawn their eggs and die. In Equation (3), denotes the survival rate during m years or the spawning stock

Table 1. Lines which were judged to be parts of clockwise loops or parts of anticlockwise loops.
biomass per recruitment (SPR), i.e., . Therefore, when the population reproduces according to Model 1, Rt and St+m are constant regardless of year.
Model 2 is the case in which when f(xt) in Equation (1) can be expressed by 1 + r. That is,
(4)
The increasing or decreasing rate, r, is determined by environmental factors. When environmental factors are good for the stock, r takes positive values (r > 0) and R increases. On the contrary, when environmental factors are bad for the stock, r takes negative values (−1 < r < 0) and R decreases. In this model, the survival process is the same of that shown in Equation (3).
Model 3 is the case when r in Equation (4) changes cyclically. It can be expressed by a sine curve as defined below:
(5)
Thus,
(6)
Here, and denote the amplitude of the sine curve and angular velocity, respectively. In this model, the survival process is the same of that shown in Equation (3).
Generally, the spawning stock biomass in year t − 1 (St−1) produces the recruitment in year t (Rt), and the recruitment in year t (Rt) becomes the spawning stock biomass in year t + m (St+m). Then the spawning stock biomass in year t + m (St+m) produces the recruitment in year t + m + 1 (Rt+m+1). This cycle repeats infinitely as shown in Figure 1. When the year is t + m + 1, the SRR can be expressed by Equation (7),
. (7)
As shown in Figure 1, Equation (7) can be modified by,
. (8)
That is, the relationship from St+m to Rt+m+1 is replaced by the relationship from Rt to Rt+m+1. Equations (7) and (8) reveal an important fact that is hidden behind a SRR. That is, the relationship from St+m to Rt+m+1, which is the SRR itself, is the relationship from Rt to Rt+m+1, which is so to speak “R to R relationship”. This relationship is the same in Model 3 as shown in Figure 1. Therefore, when the environmental factors cyclically fluctuate, such as a sine curve, the stock-recruitment relationship simply means the relationship between Rt+m+1 and Rt. In other words, a stock-recruitment relationship shows only a relationship between two different points at t and t + m + 1 on the same sine curve (Figure 2). Therefore, when the recruitment fluctuates cyclically in response to environmental factors, the stock-recruitment relationship necessary shows loop shapes. Further the time lag is small, the SRR shows clockwise loops, and the time lag is large, the SRR shows anticlockwise loops.

Figure 1. The stock-recruitment relationship in Model 2 and Model 3. The relationship from St+m to Rt+m+1 is replaced by the relationship from Rt to Rt+m+1.
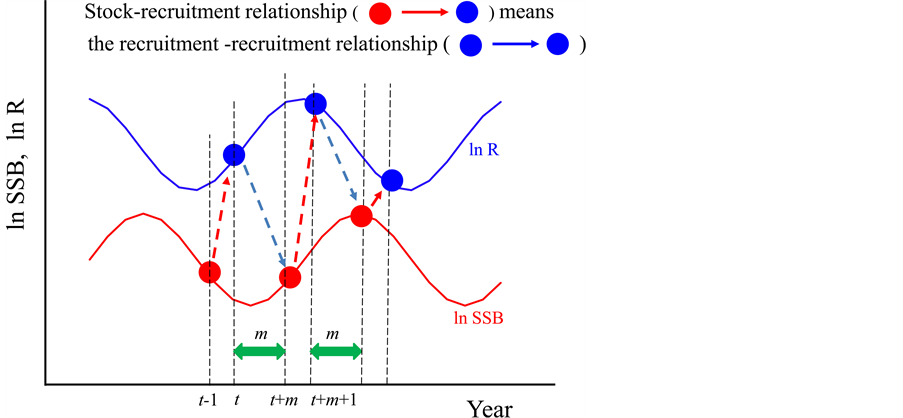
Figure 2. Stock-recruitment relationship. The relationship from St+m to Rt+m+1 merely implies the relationship from Rt to Rt+m+1.
2.3. Rule for Judging the Clockwise or Anticlockwise Direction of Loops in the SRR
Using actual data for 24 fish stocks, we investigated whether or not loop shapes emerge in SRRs by plotting ln(Rt+m) against ln(SSBt). Further, we investigated the directions of the loops depending on the age at maturity.
We constructed a rule that determined the direction of the loops, either clockwise or anticlockwise. The direction of the line from year t to year t + 1 is judged based on the direction of the line from year t + 1 to year t + 2 (Figure 3). When the direction of the line from year t + 1 to year t + 2 is “A” shown in Fig. 3, the line from year t to year t + 1 is judged to be part of a clockwise loop. When the direction of the line from year t + 1 to year t + 2 is “B”, the line from year t to year t + 1 is judged to be part of an anticlockwise loop. After all directions of the lines from year t to year t + 1 were determined, we modified the judgement regarding the direction. When three successive lines were judged to have clockwise, anticlockwise, and clockwise directions, respectively, we replaced the middle anticlockwise direction with a clockwise direction, and we replaced the judgement with clockwise, clockwise, and clockwise because this seemed to be the more reasonable conclusion. If the series of directions was anticlockwise, clockwise, and anticlockwise, the middle clockwise” was replaced with anticlockwise, and we concluded the directions to be anticlockwise, anticlockwise and anticlockwise.
2.4. Three Regression Methods
In this study, we used three regression methods to plot ln(Rt+m) against ln(SSBt): the simple, Deming [15] , and Passing and Bablok [16] . A simple regression analysis is problematic, because it assumes that the independent variable contains no observation and process errors. Therefore, parameters estimated using a simple regression analysis usually have serious biases [17] [18] [19] [20] . When both independent and dependent variables contain observation and/or process errors, both the Deming and Passing and Bablok regression analyses can effectively re-

Figure 3. Rule used to judge the direction of the line from t to t + 1. When the line from t + 1 to t + 2 is A, the line from t to t + 1 is judged to be part of a clockwise loop. When the line from t + 1 to t + 2 is B, the line from t to t + 1 is judged to be part of an anticlockwise loop.
move the biases inherent in the results of a simple regression analysis. The programs developed by Aoki [21] [22] were used when these two regression analyses were applied to the data.
3. Results
Clockwise or Anticlockwise Loops in the SRR
Figure 4 shows the SRRs for 24 stocks. For almost all 24 stocks, loops were recognized. Table 1 shows the number of lines that shows a clockwise loop or an anticlockwise loop. We also show the age at maturity. Table 1 shows that when the age at maturity was equal to one year old, for all three stocks the number of lines that showed a clockwise loop was greater than the number that showed an anticlockwise loop. When the age at maturity was one-half or 2 years old, the number of lines that showed a clockwise loop was dominant except in two stocks. That is, the Seto Island Sea stock of Japanese Spanish mackerel and the Pacific stock of blue mackerel showed the opposite results, i.e., the number of lines that showed an anticlockwise loop was greater than the number that showed a clockwise loop. When the age at maturity was between 3 and 4 years old, loops in both directions appeared. For 8 out of 12 stocks, clockwise loops were dominant, and in the other 4 stocks anticlockwise loops were dominant. When the age at maturity was more than or equal to 6 years old, anticlockwise loops were dominant for both stocks.
Table 2 shows the results of three regression analyses. Figure 5 shows the plots of the slope estimated by a simple regression analysis against the age at maturity. The regression line in Figure 5 is shown below:
(9)
where b and m denote the slope of the regression line and the age at maturity. The p-value of slope b is 0.0125 and the 95% confidence interval of b is (−0.333, −0.00449). That is, a significant negative relationship was detected between the slope of the regression line (b) and the age at maturity (m). This result coincided well with the results of the simulation by Sakuramoto [12] .
It is generally believed that simple regression analyses commonly underestimate the slope of the regression line in response to the observed and/or process errors that exist in the independent variable (Table 2). Even in cases when the slope was estimated to be less than unity by a simple regression analysis, the slope estimated using Deming and/or Passing-Bablok regression analyses sometimes changed to unity or more than unity. Such changes occurred in 6 stocks, which are shaded blue in Table 2. Only 3 out of 24 stocks showed negative slopes for all three regression analyses, and these are shaded red in Table 2. The slopes for 8 out of 24 stocks were estimated to be unity or more than unity, and these are shaded green in Table 2. For the other 7 stocks, the slopes could not be estimated at least by one method. Therefore, only 3 out of 17 stocks showed negative density-dependent effects, and 14 out of 17 stocks showed no density-dependent effect or positive density-dependent effects.








Figure 4. Stock-recruitment relationships for all 24 stocks that live in the waters around Japan. Clockwise loops or anticlockwise loops emerged.
Table 2. Slopes and 95% confidence limits of the regression lines estimated by the simple (OLS), Deming, and Passing-Bablok (P-B) regression methods. Red shows the cases when the estimated slopes were negative for all three methods. Blue shows the cases when the estimated slopes estimated by the simple regression method were less than unity, but, the slopes estimated by the Demin and/or Passing-Bablok (P-B) regression methods were unity or greater than unity. Green shows the cases when the slopes estimated by all three regression methods were equal to unity or greater than unity.
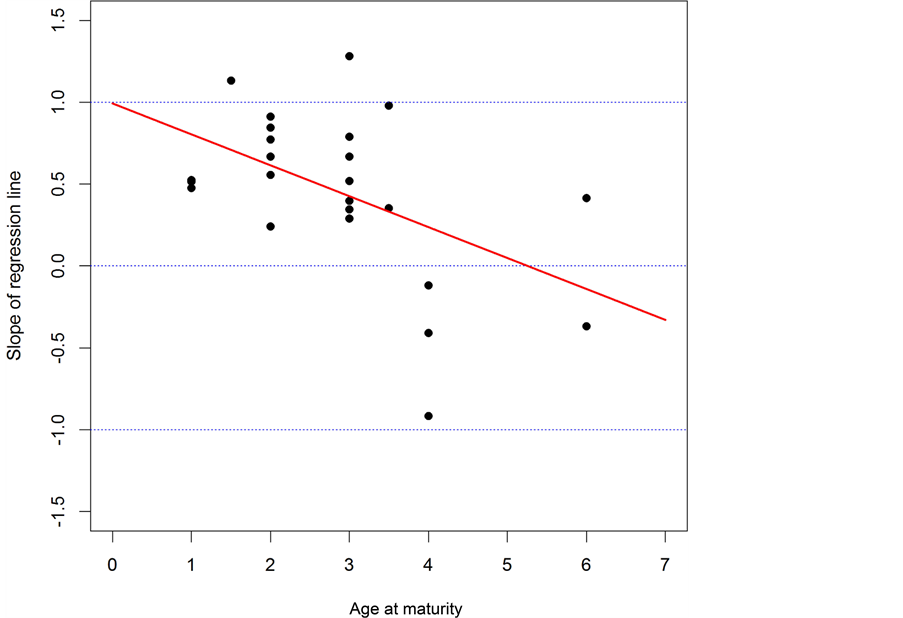
Figure 5. The relationship between the slope of the regression lines and the age at maturity. The regression line estimated is b = 0.992 - 1.89 m.
4. Discussion
4.1. Clockwise or Anticlockwise Loops in the SRR
Sakuramoto explained the mechanism that a clockwise loop or an anticlockwise loop in SRR were necessary detected [23] . In this paper, we again explained the mechanism that must appears loops in SRR. The plot of the S-R relationship merely implies the plot of the R-R relationship. Therefore, loops necessarily appear in SRR. The SRR appears to be a complex relationship. Loops should appear for all SRRs. This paper elucidated this fact using 24 stocks that live around Japan. We believe that the same results will be obtained for other stocks that live in other areas.
When the age at maturity was low, clockwise loops were dominant, and when the age at maturity was high, anticlockwise loops were dominant. However, with regard to the direction of the loop, two exceptions were observed. For the Seto Island Sea stock of Japanese Spanish mackerel and the Pacific stock of Blue mackerel, anticlockwise loops were dominant, although the ages of these stocks at maturity were low, i.e. 1.5 years old and 2 years old, respectively. In the case of the Seto Island Sea stock of Japanese Spanish mackerel, the first seven lines appear to show clockwise loops (See Figure 4(d)). Hasegawa et al. [24] noted the possibility that even when the age at maturity is low, the SRR could show anticlockwise loops when strong species interactions can be assumed to exist. Further, the direction of the loops would be strongly influenced by the cycle of environmental conditions. When the length of the cycle of environment conditions was short, an anticlockwise loop would likely occur. In almost all cases, the present results were coincided well with the theory proposed by Sakuramoto [12] [23] , with only two exceptions.
The time series data available is also an important factor to construct the loops. When the time series data is short, it is difficult to construct a loop such as the cases of a and x in Figure 4. On the contrary, when the time series data is too long, two or more than two loops are constructed and it is difficult to distinguish into each loop, such as the case in m in Figure 4. In this case, it is better to plot in each period of time such as the case done for Pacific bluefin tuna [12] . In the case for Pacific bluefin tuna [12] , the 60-year time series data available were separated into four periods and detected the three anti-clock loops.
4.2. Slope of the Regression Lines in SRR
Sakuramoto noted that the slopes of the regression lines adapted for SRR were determined by the age at maturity and the length of the cycle of environmental factors [12] [23] . When the age at maturity is low, the slope estimated by a simple regression method in which ln(Rt+m) is plotted against ln(SSBt) shows a positive value, although the slope is statistically less than unity. When the age at maturity is high, the slope of the regression line is close to zero. Furthermore, the age at maturity becomes higher, and the slope of the regression line becomes negative. Sakuramoto showed that the slope of the Pacific stock of Japanese sardines was close to unity, although it was statistically less than unity. This finding is very important because it shows that a density-dependent effect is easily detected for stocks which have a low age at maturity. In contrast, the results of this paper show that no relationship between recruitment and SSB could be easily detected for stocks which have a high age at maturity. For instance, the SSR of Pacific bluefin tuna showed no relationship between R and SSB, and the plots were widely scattered without any trend [12] [13] . The Beverton and Holt model or hockey stick model is usually applied as the SSR for Pacific bluefin tuna, and the management procedure has been widely discussed. However, this paper suggests that the Beverton and Holt model or hockey stick model is not appropriate for this stock, and so a discussion based on those SRR model would not be fruitful.
In general, the clockwise or anti-clockwise loop observed in SRR cannot be explained using the density-dependent effect. This fact indicates that the most important relationship between stock and recruitment is likely to be the interspecific relationship and/or environmental conditions, not the density-dependent effect, and the density-dependent effect observed in SRR is not real. All the management procedures that are constructed based on the density-dependent effect should the revised, because the density-dependent effect observed in SRR is not valid.
5. Conclusions
The results elucidated in this paper can be summarized as follows:
1) Loop shapes emerged in SRR for almost all 24 stocks investigated in this paper.
2) When the age at maturity was low, clockwise loops were dominant, and when the age at maturity was high, anticlockwise loops were dominant.
3) The slope of the regression line for SRR had a negative correlation with the age at maturity. When the age at maturity was low, the slope was positive but less than unity. When the age at maturity became high, the slope decreased to zero, and as the age at maturity increased further, the slope became negative.
Acknowledgements
I would like to thank KN International for improving the manuscript.
Cite this paper
Tanaka, K., Su- zuki, N. and Sakuramoto, K. (2017) Clock- wise Loops and Anticlockwise Loops Ob- served in a Stock-Recruitment Relation- ship. Open Access Library Journal, 4: e3688. https://doi.org/10.4236/oalib.1103688
References
- 1. Beamish, R.J. and Bouillon, D.R. (1993) Pacific Salmon Production Trends in Relation to Climate. Canadian Journal of Fisheries and Aquaculture Sciences, 50, 1002-1016.
https://doi.org/10.1139/f93-116 - 2. Myers, R.A. and Cadigan, N.G. (1993) Density-Dependent Juvenile Mortality in Marine Demersal Fish. Canadian Journal of Fisheries and Aquaculture Sciences, 50, 1576-1590.
https://doi.org/10.1139/f93-179 - 3. Ricker, W.E. (1954) Stock and Recruitment. Journal of the Fisheries Research Board of Canada, 11, 559-623.
https://doi.org/10.1139/f54-039 - 4. Beverton, R.J.H. and Holt, S.J. (1957) Fishery Investigations. Series 2, 19, HMSO, London.
- 5. Su, Z., Peterman, R.M. and Haeseker, S.L. (2004) Spatial Hierarchical Bayesian Models for Stock-Recruitment Analysis of Pink Salmon (Oncorhynchus gorbuscha). Canadian Journal of Fisheries and Aquaculture Sci-ences, 61, 2471-2486.
https://doi.org/10.1139/f04-168 - 6. Sundby, S. (2000) Recruitment of Atlantic Cod Stocks in Relation to Temperature and Advection of Copepod Populations. Sarsia, 85, 277-298.
https://doi.org/10.1080/00364827.2000.10414580 - 7. Chen, D.G. and Irvine, J.R. (2001) A Semiparametric Model to Examine Stock-Recruitment Relationships Incorporating Environmental Data. Canadian Journal of Fisheries and Aquaculture Sciences, 58, 1178-1186.
- 8. Sakuramoto, K. (2005) Does the Ricker or Beverton and Holt Type of Stock-Recruitment Relationship Truly Exist? Fisheries Science, 71, 577-592.
https://doi.org/10.1111/j.1444-2906.2005.01002.x - 9. Sakuramoto, K. (2013) A Recruitment Forecasting Model for the Pacific Stock of the Japanese Sardine (Sardinops melanostictus) That Does Not Assume Density-Dependent Effects. Agricultural Sciences, 4, 1-8.
https://doi.org/10.4236/as.2013.46A001 - 10. Sakuramoto, K. (2013) A Common Concept of Population Dynamics Applicable to Both Thrips imaginis (Thysanoptera) and the Pacific Stock of the Japanese Sardine (Sardinops melanostictus). Fisheries and Aquaculture Journal, 4, 140-151.
- 11. Sakuramoto, K. (2015) Illusion of a Density-Dependent Effect in Biology. Agricultural Sciences, 6, 479-488.
https://doi.org/10.4236/as.2015.65047 - 12. Sakuramoto, K. (2015) A Stock-Recruitment Relationship Applicable to Pacific Bluefin Tuna and the Pacific Stock of Japanese Sardine. American Journal of Climate Change, 4, 446-460.
https://doi.org/10.4236/ajcc.2015.45036 - 13. Sakuramoto, K. (2016) A Simulation Model of the Spawning Stock Biomass of Pacific Bluefin Tuna and Evaluation of Fisheries Regulations. American Journal of Climate Change, 5, 245-260.
https://doi.org/10.4236/ajcc.2016.52021 - 14. NPAFC.
http://www.npafc.org/new/science_statistics.html - 15. Deming, W.E. (1943) Statistical Adjustment of Data [1-261]. Dover Publication Inc., New York.
- 16. Passing, H. and Bablok, W. (1983) New Biometrical Procedure for Testing the Equality of Measurements from Two Different Analytical Methods. Application of Linear Regression Procedures for Method Comparison Studies in Clinical Chemistry, Part I. Journal of Clinical Chemistry and Clinical Biochemistry, 21, 709-720.
- 17. Maelezer, D.A. (1970) The Regression of Log Ni+1 on log Ni as a Test of Density Dependence: An Exercise with Computer-Constructed Density-Dependent Populations. Ecology, 51, 810-822.
https://doi.org/10.2307/1933973 - 18. Kuno, E. (1971) Sampling Error as a Misleading Artifact in “Key Factor Analysis”. Researches on Population Ecology, 13, 28-45.
https://doi.org/10.1007/BF02522011 - 19. Ito, Y. (1972) On the Methods for Determining Density-Dependence by Mean of Regression. Oecologia, 10, 347-372.
- 20. Sakuramoto, K. and Suzuki, N. (2012) Effect of Process and/or Observation Errors on the Stock-Recruitment Curve and the Validity of the Proportional Model as a Stock-Recruitment Relationship. Fisheries Science, 78, 41-45.
https://doi.org/10.1007/s12562-011-0438-4 - 21. Aoki, S. (2009) Parameter of a Re-gression Line Estimated by Deming Regression Method.
http://aoki2.si.gunmau.ac.jp/R/Deming.html - 22. Aoki, S. (2009) Parameter of a Regression Line Estimated by Passing Bablok Regression Method.
http://aoki2.si.gunma-u.ac.jp/R/PassingBablok.html - 23. Sakuramoto, K. (2016) Density-Dependent Effect Occurs Regardless of Density. Open Access Library Journal, 3, e3112.
https://doi.org/10.4236/oalib.1103112 - 24. Hasegawa, S., Suzuki, N. and Sakurqmoato, K. (2017) On a Catch-Forecasting Model for the Pink Salmon Oncorhynchus gorbuscha in the Maritime Province of Siberia. Open Access Library Journal, 4, e3406.
https://doi.org/10.4236/oalib.1103406