 Journal of Environmental Protection, 2011, 2, 769-777 doi:10.4236/jep.2011.26089 Published Online August 2011 (http://www.SciRP.org/journal/jep) Copyright © 2011 SciRes. JEP 769 Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature J. Michael Kelly1, Frank C. Thornton2, J. Devereux Joslin3 1College of Natural Resources and Environment, Virginia Tech, Blacksburg, USA; 2Tennessee Valley Authority, Muscle Shoals, USA; 3Belowground Forest Research, Monteverde, Costa Rica. Email: jmkelly@vt.edu Received April 28th, 2011; revised June 14th, 2011; accepted July 27th, 2011. ABSTRACT A better understanding of the mechanisms that control nutrient acquisition in the context of plant and ecosystem re- sponses to climate change is needed. Mechanistic nutrient uptake models provide a means to investigate some of the impacts of temperature change on soil nutrient supply and root uptake kinetics through the simulation of key soil and plant processes. The NST 3.0 model, in combination with literature values on plant and soil parameters from a red spruce (Picea rubens L.) site in the southern Appalachians, was used to conduct a series of model simulations fo cused on the combined effects of changes to the maximal rate of nutrient influx at high concentrations (Imax), root growth rate (k), concentration of nutrient occurring in the soil solution (Cli), and th e ability of the so il solid phase to buffer chang es to the soil solution nutrient concentration (b). Previous research has indicated that these four parameters are respon- sive to changes in root zone temperature. Simulated uptake of NH4 increased by a factor of up to 2.6 in response to in- creases in soil temperature of 1˚C to 5˚C. The model also projected an increa se in P uptake coupled with up to an 80% reduction in solution P concentration in response to a 1˚C - 5˚C increase over a 147-d simulation period. These hypo- thetical changes, if validated, have interesting implications for plant growth and competition and point to a need for additional studies to better define the impacts of soil tempera ture on soil nutrient supply and root uptake. Keywords: Mechanistic Modeling, Imax, Root Growth Rate, Soil Buffer Power 1. Introduction Over the next 100 years, mean global temperature is pro- jected to increase by 1.5˚C to 4.5˚C [1]. An increase in atmospheric temperature will eventually lead to an in- crease in soil temperature [2], and this increase in soil temperature could lead to changes in soil supply and plant nutrient uptake rates. Most studies of soil warming in the forest environment have focused on changes in plant productivity and soil nutrient availability [3-6] with only Gessler et al. [7] and Adam et al. [8] assessing the potential changes in nutrient uptake as a function of changes in root zone temperature. In a laboratory study on the influence of temperature on solution phase nutrient concentrations in soil from a southeastern US spruce-fir stand, Kelly [9] found higher concentrations of NH4 and P as soil temperature in- creased from 4˚C to 24˚C. Conversely, NO3 concentra- tion in solution was greatest at 4˚C and declined as tem- perature increased. Since nutrient uptake is thought to occur largely through root interaction with the solution phase [10], changes in soil temperature, and associated changes in soil solution chemistry have the potential to alter both nutrient availability and plant uptake. And since nutrient uptake is a physiologically mediated proc- ess, it also follows that changes in soil temperature could have an impact on the rate at which plants take up nutri- ents. For example, Adam et al. [8] in a greenhouse study found that the uptake of NO3 by red maple plants in- creased by approximately 2% per degree of temperature increase as solution temperature increased to an optimum and then began to decline. Adam et al. [8] also report that root surface area was influenced by differences in root zone temperature, with both root length and root radius values showing a response pattern similar to that of ni- trate. Similarly, Weih and Karlsson [5] found that moun- tain birch root-N uptake rate and plant-N concentration were positively correlated with increases in soil tem- perature. In a field and laboratory study of Norway spruce, Gessler et al. [7] found that very little NO3 was taken up by roots when solution NH4 concentrations were elevated and that NH4 uptake increased by ap-  Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature 770 proximately 10% to 13% per ˚C increase in root zone temperature. Agronomic studies suggest that when soil P supplies are low, P availability increases with increasing soil temperature [11-13]. It stands to reason that this rela- tionship would also hold in unfertilized forest soils, since in both agricultural and forest soils, decomposition of organic matter is the primary source of P. These and other studies have created a body of knowledge that, when combined through a mechanistic nutrient uptake model, provide a means to explore hypothetically the impacts of soil temperature change on soil nutrient sup- ply and nutrient uptake by an important tree species. The NST 3.0 mechanistic nutrient uptake model pro- vides a means to simulate the short-term impact of soil temperature change on the concentration of nutrients occurring in the soil solution as well as the impact of changes in root growth rate on nutrient acquisition. Similarly, the impacts of temperature mediated changes on physiological processes can be explored indirectly through alterations of the magnitude of the uptake kinet- ics parameters. NST 3.0 is available for download from the website of the Department of Crop Sciences at Göt- tingen University (http://wwwuser.gwdg.de/~uaac/). The NST 3.0 model, and its predecessor the Barber-Cushman model [14], have been used with a high degree of success to explore nutrient uptake under a variety of circum- stances [15-19]. As noted by Claassen and Steingrobe [18], a validated mechanistic model provides a means to extrapolate plant response beyond currently available data and also pro- vides a way to evaluate potential hypotheses. Thus, the objective of this study is to use a well-established mech- anistic nutrient uptake model, NST 3.0, to explore the potential impacts of 1˚C, 2˚C, 3˚C, and 5˚C increases in simulated soil temperature on estimates of nutrient up- take by red spruce seedlings over a growing season. We will do this by altering the magnitude of the values for the initial solution concentration (Cli), the ability of the soil solid phase to resupply the solution phase (b), the root growth rate (k), and the maximal rate of nutrient influx at the root surface (Imax). The results of these comparisons should provide initial theoretical insights into the potential changes in N and P supply and uptake that might occur in response to changes in soil tempera- ture as a consequence of global climate change. 2. Materials and Methods Published data taken from studies conducted in the spruce-fir forest at Whitetop Mountain in southeastern Virginia and reported by Kelly and Mays [20], Thornton et al. [21], Kelly et al. [22], and Kelly [9] will form the basic data set used in this analysis (Table 1). These data sets will be supported by additional information from the same site reported by Joslin and Wolfe [23,26]. In some cases the values listed in Table 1 are taken directly from Table 1. Transport, sorption, and root parameters used in the NST 3.0 model to describe N and P uptake by red spruce seed- lings under base conditions (Letters in parentheses following each value indicate the data source corresponding to the cita- tions listed in the footnote). Parameter Units N P DL Diffusion coefficient in H2O cm2·s−1 1.9E−5 (a)† 8.9E−6 (b) Θ Volumetric H2O content cm3 H2O/cm3 soil 0.22 (c) 0.22 (c) f Impedance factor unitless 0.178 (c) 0.178 (c) Vo H2O uptake at root cm·s−1 6.3E−8 (d) 6.3E−8 (d) Cli Initial solution concentration mol·cm-3 1.17E−6 (e) 3.42E−9 (e) b Buffer power unitless 133 (c) 37 (d) Imax Maximum influx at high concentration mol·cm−2·s−1 2.1E−11 (f) 2.68E−13 (g) Km Solution concentration when influx is 0.5 Imax mol·cm−3 2.06E−4 (f) 1.6E−8 (g) Cmin Solution concentration when influx is zero mol·cm−3 1.11E−9 (f) 6.0E−11 (g) ro Root radius cm 0.042 (h) 0.042 (h) r1 Half distance cm 0.31 (h) 0.31 (h) Lo Initial root length cm 37860 (h) 37860 (h) k Root growth rate cm·d−1 315 (h) 315 (h) † (a) [24]; (b) [25]; (c) [20]; (d) [9]; (e) calculated by model; (f) [19]; (g) [22]; (h) [21]. Copyright © 2011 SciRes. JEP  Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature771 the published work cited in Table 1 or re-calculated from these values when not available as a direct result of these studies. For parameters that could not be calculated or extrapolated from on-site data directly available to the authors, missing parameter values were obtained from the literature sources noted or calculated with the model using a process similar to that described in Kelly et al. [17]. Values for the NH4 uptake kinetics parameters Imax, Km, and Cmin were taken from a study of white spruce conducted by Hangs et al. [19]. Phosphorus values for the same parameters came from a study of loblolly pine conducted by Kelly et al. [22]. While it would be prefer- able to have actual values for red spruce, values from these two conifer species are thought to provide a rea- sonable first approximation. In this study we used the NST 3.0 model to focus on the impacts of temperature change on the level of nutri- ent occurring in the soil solution (Cl), the ability of the soil solid phase nutrients to sustain the solution phase concentration (b), the root growth rate (k), and the max- imal rate of nutrient uptake under unlimited nutrient availability (Imax). These four parameters were chosen for evaluation because of their demonstrated sensitivity to change as a function of temperature [8,9]. Values used for these and the other parameters required for the base case simulations of N and P using NST 3.0 are listed in Table 1 and their source identified. Percentage modifications (+/−) to the base level values used for k and Imax to simulate changes in temperature (Table 2) are based on the relations observed by Adam et al. [8] for red maple. In a study of root zone temperature influence on root growth and nitrate uptake by red maple, Adam et al. [8] found that root growth rate increased by an average of 4.3% per degree of temperature change as the root zone temperature was increased from 14˚C to 24˚C and that the nitrate Imax value increased by 1.9% per degree increase over the same temperature range. Given the absence of actual measurements for red spruce, these two percentage values were assumed to represent the relative change rate in k and Imax for both nutrients (Ta- ble 2). It should be noted however, that the temperature range used by Adam et al. [8] exceeds the range of soil temperature increase likely to occur in soils currently supporting the growth of red spruce. Similarly, in a controlled environment investigation of changes in soil solution NH4 and P in the surface horizon of a soil from a spruce-fir stand on Whitetop Mountain, Kelly [9] found that NH4 concentration increased by 10% per degree as temperature increased from 4˚C to 24˚C while soil solution P concentration increased by 13.1% per degree C. Using the relationship between Cli and b reported by Kelly et al. [27], the corresponding b values for NH4 and P were estimated to decrease by 10% for Table 2. Percentage change factors for Tests I and II (+/− %) per degree centigrade increase in simulated soil tempera- ture for Cli, b, k, and Ima x for simulations by NST 3.0 of N and P uptake by red spruce seedlings. N P Parameter Test I Test II Test I Test II Change % Cli +10 † +13.1 b −10 † −6.5 k +4.3 +10 +4.3 +10 Imax +1.9 +10 +1.9 +10 †Cli and b values were unchanged in Test II, only the k and Imax values were modified according to the Q10 relationship. NH4 and 6.5% for P per degree of temperature increase (Table 2). For the purposes of this analysis all nitrogen uptake is assumed to be in the form of NH4 because of the established preference of conifers for this form of N [7,28]. Using the change values listed in Table 2 for the four model parameters, a suite of uptake values was cal- culated for each nutrient to represent simulated tempera- ture increases of 1˚C, 2˚C, 3˚C, and 5˚C. All four values for each nutrient-temperature scenario were entered concurrently while the values for the remaining nine pa- rameters in the model were held constant at the level specified for each nutrient in Table 1. In addition to the change factors for k and Imax identi- fied for Test I in Table 2, an additional set of values was calculated for these two physiologically mediated pa- rameters based on a simple Q10 relationship. Thus for the data sets based on the Q10 relationship (Test II, Table 2), k and Imax were each increased by 10% per ˚C of tem- perature increase. This level of change for Test II was chosen in part based on the study of Norway spruce by Gessler et al. [7] who found an increase in NH4 uptake of approximately 10% - 13% per ˚C increase in root zone temperature. The values for Cli and b in the Q10 simula- tions (Test II) continued to change at the rates indicated for Test I in Table 2, while the remaining values for the other nine parameters listed in Table 1 were again held constant at the levels indicated. Nine scenarios were run for each nutrient-temperature combination for a total of 18 simulations covering both nutrients. 3. Results and Discussion 3.1. Validity of Assumptions Before proceeding further it is appropriate to consider the assumptions that have been made concerning the magni- tudes and directions of change associated with the Cli, b, k, and Imax values used for each nutrient. Unfortunately, Copyright © 2011 SciRes. JEP  Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature 772 studies of soil temperature influences focused directly on these parameters are very limited for woody species. Therefore we have depended heavily on the study of Adam et al. [8] even though their study was conducted with a deciduous species and used a temperature range likely to exceed temperatures in red spruce forest soils. We combined the insights from Adam et al. [8] with ac- tual seasonal totals of N and P uptake by red spruce seedlings from the work of Thornton et al. [21]. The Test I values are taken to be at the lower end of the potential response spectrum. The observations of Gessler et al. [7] were used to inform the other end (Test II) of what we feel is a conservative spectrum of input parameters that are consistent with observed growing season uptake based on the data of Thornton et al. [21]. Therefore, the focus of this study is to develop an assessment of “rela- tive change” rather than providing “absolute values”. While these uptake estimates are hypothetical, they have the potential to raise important research questions. These questions in turn could lead to the formulation of future studies specifically designed to address possible changes in nutrient uptake in response to changing soil tempera- ture. 3.2. Nitrogen Using the parameter values listed in Table 1, the model estimate of NH4 uptake over a 147-d period was on the order of 24.0 mmols. For the purpose of this study this value is considered to be the base level of N uptake and is equivalent to the level of uptake actually observed for the red spruce seedlings described by Thornton et al. [21] during their second year of growth. This level of corre- spondence between the model calculation and observed uptake was achieved by adjusting the Cli value to a level that, in conjunction with the other values entered into the model (Table 1), would combine to produce a level of model predicted uptake equivalent to the observed uptake. The same procedure was used to establish the base level of uptake for P as well. In the case of NH4, this led to a higher value for Cli (1.17E−6 mol·cm−3) being used in the model than was reported by Kelly [9] for a spruce-fir soil at the Whitetop Mountain site (5.27E−8 mol·cm−3). However, it is not unreasonable to assume that even though the soil used in the pot study conducted by Thornton et al. [21] came from a similar site on Whitetop Mountain, the handling of the soil and the warmer environment in the chambers used by Thornton et al. [21] led to a higher level of N availability to the study seedlings. This supposition is consistent with the observation of Kelly [9] that soil so- lution NH4 levels increased as soil incubation tempera- ture increased, as well as the findings of Rustad and Fer- nandez [4] who reported that a 4˚C - 5˚C increase in soil temperature in a red spruce forest stand stimulated de- composition and the subsequent release of N from or- ganic sources. Estimates of N uptake derived by the model in re- sponse to each of the temperature increase scenarios in- dicate steady increases in uptake with the highest esti- mate using the Test I parameters being 1.75 times that occurring under base conditions (Table 3). When Q10 values are used to represent further increases in Imax and k (Test II), the level of N uptake estimated at 5˚C increases by a factor of 2.6 over base conditions. This level of in- crease, if realized, could stimulate the growth of estab- lished red spruce, assuming that current growth rate is N limited. The predicted level of increased uptake could also have other impacts such as the creation of imbal- ances with other nutrients that could slow growth, or increased susceptibility to freeze damage due to early bud out or delayed senescence [29]. Alternatively, an increase in nitrogen availability could stimulate the growth or establishment of other species that might be better suited to a more favorable nitrogen supply and warmer soil than red spruce and ultimately lead to the displacement of red spruce by another species over time [30]. To explore these model results further, a series of sin- gle factor sensitivity analyses were conducted using the approach described by Silberbush and Barber [31] to see if a particular one of the four parameters evaluated was driving the response. Using the 1˚C increase scenario as an example, the results of the sensitivity analyses in- dicted that for NH4 the Imax and Cli values were the most sensitive to change and both produced essentially the same level of response (Figure 1). Root growth rate (k) responded to a lesser degree with NH4 uptake exhibiting a 19% decline when the k value was reduced by 50% and a 40% increase when the k value was doubled (Figure 1). Table 3. Estimates of N and P uptake by red spruce seed- lings over a 147-d period as a function of simulated soil temperature changes of 1˚C, 2˚C, 3˚C, or 5˚C based on NST 3.0 calculations. Uptake (mmols) N P Scenario Test I Test II Test I Test II Base 24.0 1.48 +1˚C 27.3 29.7 1.61 1.66 +2˚C 30.8 37.0 1.77 1.87 +3˚C 34.4 44.8 1.86 2.01 +5˚C 42.2 63.2 2.06 2.26 Copyright © 2011 SciRes. JEP 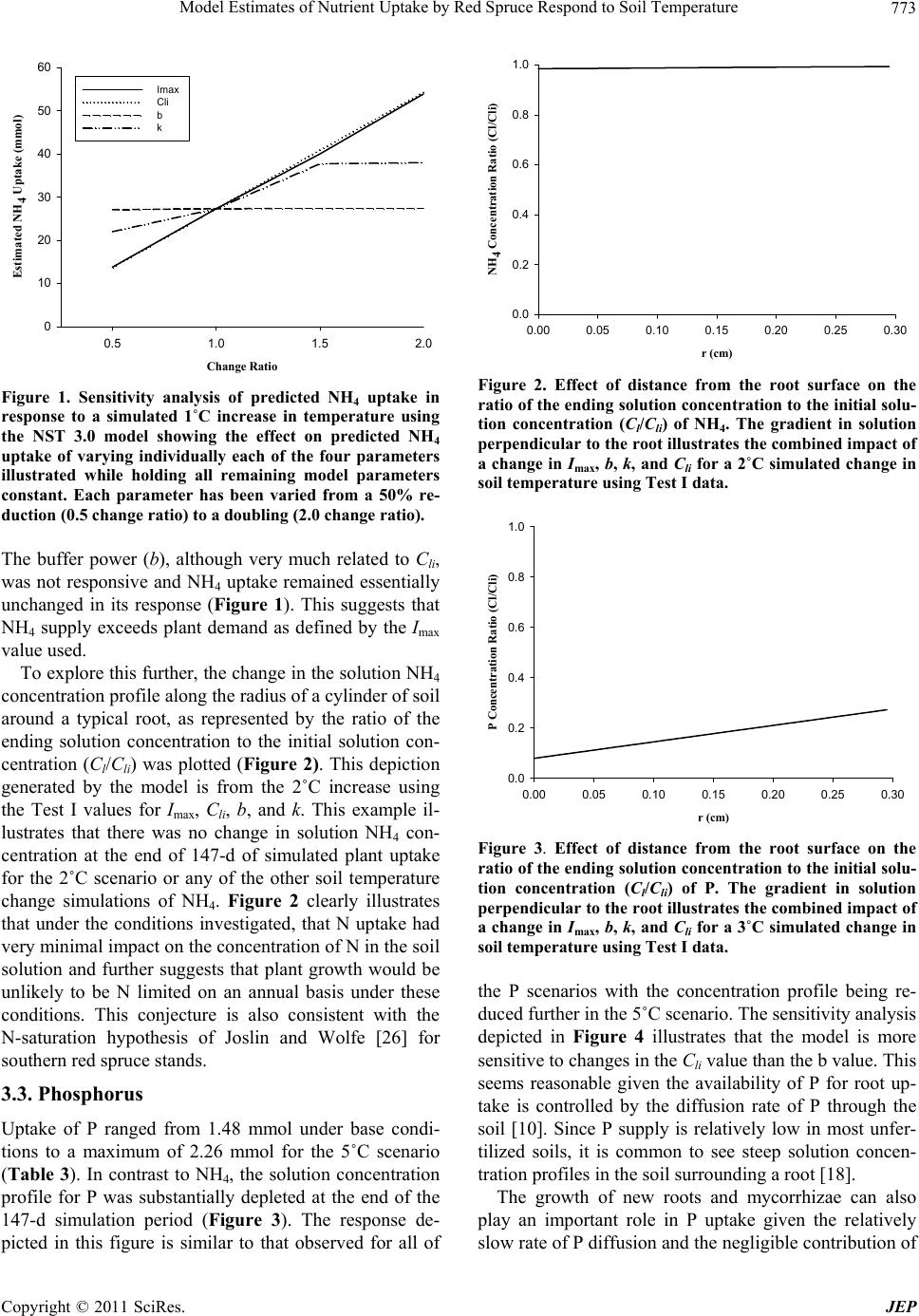 Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature773 Change Ratio 0.5 1.0 1.5 2.0 Estimated NH4 Uptake (mmol) 0 10 20 30 40 50 60 Imax Cli b k Figure 1. Sensitivity analysis of predicted NH4 uptake in response to a simulated 1˚C increase in temperature using the NST 3.0 model showing the effect on predicted NH4 uptake of varying individually each of the four parameters illustrated while holding all remaining model parameters constant. Each parameter has been varied from a 50% re- duction (0.5 change ratio) to a doubling (2.0 change ratio). The buffer power (b), although very much related to Cli, was not responsive and NH4 uptake remained essentially unchanged in its response (Figure 1). This suggests that NH4 supply exceeds plant demand as defined by the Imax value used. To explore this further, the change in the solution NH4 concentration profile along the radius of a cylinder of soil around a typical root, as represented by the ratio of the ending solution concentration to the initial solution con- centration (Cl/Cli) was plotted (Figure 2). This depiction generated by the model is from the 2˚C increase using the Test I values for Imax, Cli, b, and k. This example il- lustrates that there was no change in solution NH4 con- centration at the end of 147-d of simulated plant uptake for the 2˚C scenario or any of the other soil temperature change simulations of NH4. Figure 2 clearly illustrates that under the conditions investigated, that N uptake had very minimal impact on the concentration of N in the soil solution and further suggests that plant growth would be unlikely to be N limited on an annual basis under these conditions. This conjecture is also consistent with the N-saturation hypothesis of Joslin and Wolfe [26] for southern red spruce stands. 3.3. Phosphorus Uptake of P ranged from 1.48 mmol under base condi- tions to a maximum of 2.26 mmol for the 5˚C scenario (Table 3). In contrast to NH4, the solution concentration profile for P was substantially depleted at the end of the 147-d simulation period (Figure 3). The response de- picted in this figure is similar to that observed for all of r (cm) 0.00 0.05 0.10 0.15 0.20 0.25 0.30 NH4 Concentration Ratio (Cl/Cli) 0.0 0.2 0.4 0.6 0.8 1.0 Figure 2. Effect of distance from the root surface on the ratio of the ending solution concentration to the initial solu- tion concentration (Cl/Cli) of NH4. The gradient in solution perpendicular to the root illustrates the combined impact of a change in Imax, b, k, and Cli for a 2˚C simulated change in soil temperature using Test I data. r (cm) 0.00 0.05 0.10 0.15 0.20 0.25 0.30 P Concentration Ratio (Cl/Cli) 0.0 0.2 0.4 0.6 0.8 1.0 Figure 3. Effect of distance from the root surface on the ratio of the ending solution concentration to the initial solu- tion concentration (Cl/Cli) of P. The gradient in solution perpendicular to the root illustrates the combined impact of a change in Imax, b, k, and Cli for a 3˚C simulated change in soil temperature using Test I data. the P scenarios with the concentration profile being re- duced further in the 5˚C scenario. The sensitivity analysis depicted in Figure 4 illustrates that the model is more sensitive to changes in the Cli value than the b value. This seems reasonable given the availability of P for root up- take is controlled by the diffusion rate of P through the soil [10]. Since P supply is relatively low in most unfer- tilized soils, it is common to see steep solution concen- tration profiles in the soil surrounding a root [18]. The growth of new roots and mycorrhizae can also play an important role in P uptake given the relatively slow rate of P diffusion and the negligible contribution of Copyright © 2011 SciRes. JEP  Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature 774 Change Ratio 0.5 1.0 1.5 2.0 Estimated P Uptake (mmol) 0.0 0.5 1.0 1.5 2.0 2.5 3.0 Imax Cli b k Figure 4. Sensitivity analysis of predicted P uptake in re- sponse to a 1˚C increase in temperature using the NST 3.0 model showing the effect on predicted P uptake of varying individually each of the four parameters illustrated while holding all remaining model parameters constant. Each parameter has been varied from a 50% reduction (0.5 change ratio) to a doubling (2.0 change ratio). mass flow to P delivery. Under base case conditions, P uptake over the study period attributed to existing roots, as calculated by the model, was on the order of 912 µmols with new roots responsible for an additional 563 µmols. At the 5˚C level using Test II data, existing roots accounted for 1.17 mmols of uptake and new roots 1.09 mmols of P uptake. In the latter case, new roots account for approximately half of the uptake, while in the base case simulation new roots provide approximately 38% of the P uptake. Increasing the degree to which roots ramify the soil has a definite impact on the uptake of P as a re- sult of shorter diffusion distance to a root. Hyphae of mycorrhizae can serve much the same function as roots and are known to be important to P ac- quisition in P limited situations. We did not have infor- mation on mycorrhizal growth rates or mycorrhizal up- take kinetics values for P or NH4. Consequently, we choose not to consider their potential contribution. Provi- sions have been made in NST 3.0 to estimate the contri- bution of root hairs to uptake. To do so, additional values for k and Imax are entered into the model. This same pro- vision could be used to get an estimate of mycorrhizal contributions as well, assuming appropriate k and Imax values were available. However, under the conditions considered here it is likely that the addition of my- corrhizae would have increased further the rate of P up- take and contributed to the further decline of plant avail- able P. 3.4. Implications of Results In this study, we simulate NH4 and P uptake using data from a southeastern US red spruce forest stand where there is evidence that growth is not consistently N-li- mited. The absence of red spruce responses to N fertili- zation on Whitetop Mt. [23] and the observation of net N-mineralization rates that exceed tree uptake require- ments in various other high-elevation red spruce-fir for- est sites [32,33] would indicate the absence of an N limi- tation for red spruce in some southeastern US locations. In light of this evidence, it appears prudent to question related assumptions given our limited knowledge of the impact that a lack of N-limitation might have. While we were able to calibrate the model to existing data on the total uptake of N by red spruce seedlings over a 147-d period, direct information on uptake responses to changes in Cl under current conditions in the southern Appalachians was not available and the assumptions was made that NH4 concentration in the soil solution would increase in response to increasing soil temperature in a manner similar to that observed by Kelly [9]. Both Karlsson and Nordell [3] and Weih and Karlsson [5] found an increase in N uptake and tissue concentration in mountain birch as soil temperature increased. In a study of oats, Nielsen et al. [34] found an increase in N uptake as soil temperature increased. Similarly, both Whitfield and Smika [35] and Gavito et al. [1] found an increase in N uptake in several wheat varieties as soil temperature increased. All of these responses are consistent with the findings of Adam et al. [8]. However, none of these studies, except for the work of Adam et al. [8], was con- ducted in a manner that allows the role of increased soil supply and increased root uptake to be assessed sepa- rately. Consequently, we assumed that Imax for modeled NH4 would increase with temperature either at rates ob- served by Adam et al. [8] (Test I) or according to a Q10 relationship (Test II). However, under conditions where N is not limiting growth, source-sink relationships could result in reduced NH4 uptake capacity and lower Imax values than those used in our modeling effort. Studies have documented reductions in uptake capacity for NH4 with increases in N availability for a variety of tree species, including two species closely related to red spruce—Norway spruce [36] and Engelmann spruce [28]. If these observations hold for red spruce under conditions of high N availability, the values for the increase in N uptake with increases in temperature would be lower than those predicted here. Possible secondary effects of a reduction in N uptake by red spruce include: 1) increases in the availability of NH4 for other plant species, 2) increased production of NO3,3) attendant elevation in cation concentrations in soil solu- tion, and 4) leaching losses of cations. In the case of P, we again assumed an increase in soil supply with increasing temperature based on the findings Copyright © 2011 SciRes. JEP  Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature775 of Kelly [9]. This assumption is supported by the work of Sheppard and Rocz [11] who found that the availability of P increased as soil temperature increased from 10˚C to 25˚C. They attributed this increase to desorption of bound P. Studies of P uptake by oats [13,34] and wheat [35] found a general increase in P uptake with increases in soil temperature. Del Valle and Harmon [37] found a 300% increase in P uptake in the shoots and a 200% in- crease in the roots of turnip as soil temperature increased from 7˚C to 23˚C. Conversely, Nielsen and Cunningham [38] did not find an increase in P uptake by ryegrass in response to increased soil temperature while De Lucia et al. [39] found that foliar P in big bluestem increased but root P declined in response to soil temperature increase. Given that the P response is mixed we chose to use an increasing scenario given that in most forest soils supply of P is generally low and thus an increase in availability is most likely to lead to an increase in uptake. 4. Conclusions Model predictions from the study indicate that even modest changes in soil temperature in response to global warming could lead to increased levels of N and P uptake in the near term. While the actual magnitude of the po- tential response is uncertain, if the underlying assump- tions prove to be correct, there is reason to feel that trends depicted by the model are credible given that the NST 3.0 model is based on mechanistic principles and its outputs have proven reliable in a variety of situations [18,22]. Possibly the most useful outcome of this analysis is that model results point to a number of areas that deserve further exploration through experimentation. For exam- ple, the sensitivity analyses indicate that a better under- standing of the impact of increasing temperature on plant nutrient uptake kinetics could be informative. This ap- pears particularly true in the case of NH4, since red spruce may respond differently to increases in soil supply if it is not N-limited. Similarly, changes in soil supply as reflected by changes in soil solution nutrient concentra- tions might be the most informative parameter and the least complicated to explore experimentally. As noted by Bassirirad [40], there is an urgent need to generate more data in this area and future studies of plant response to global warming should integrate measurements of soil nutrient supply and absorption into study plans. Although it was not the intent of this study to explore the ecological or whole plant implications of the simu- lated changes, the potential ecological response to re- duced P supply, in conjunction with an increase in P up- take, seems the most provocative. The potential impact of increased nitrogen availability on competition with other species for site dominance, possible changes in red spruce cold tolerance, and/or increases in nitrate produc- tion and nitrate leaching of cations also have important ecological implications and warrant further evaluation. 5. Acknowledgements The authors express their appreciation to Dr. Norbert Claassen for making the NST 3.0 Model available for public use. Thanks also to Dr. J. K. Kelly for his assis- tance in the preparation of the figures and to Alan Mays for his assistance with most of the field studies that un- derlie the papers from which these data were taken. REFERENCES [1] M. E. Gavito, P. S. Curtis, T. N. Mikkelsen and I. Ja- kobsen, “Interactive Effects of Soil Temperature, At- mospheric Carbon Dioxide and Soil N on Root Develop- ment, Biomass and Nutrient Uptake of Winter Wheat during Vegetative Growth,” Journal of Experimental Bi- ology, Vol. 52, No. 362, 2001, pp. 1913-1923. [2] J. S. King, K. S. Pregitzer and D. R. Zak, “Clonal Varia- tion in above- and below-Ground Growth Responses of Populus Tremuloides Michaux: Influence of Soil Warm- ing and Nutrient Availability,” Plant and Soil, Vol. 217, No. 1-2, 1999, pp. 119-130. doi:10.1023/A:1004560311563 [3] P. S. Karlsson and K. O. Nordell, “Effects of Soil Tem- perature on the Nitrogen Economy and Growth of Moun- tain Birch Seedlings Near Its Presumed Low Temperature Distribution Limit,” Ecoscience, Vol. 3, 1996, pp. 183- 189. [4] L. E. Rustad and I. J. Fernandez, “Soil Warming: Conse- quences for Foliar Litter Decay in a Spruce-Fir Forest in Maine, USA,” Soil Science Society of America Journal, Vol. 62, No. 4, 1998, pp. 1072-1080. doi:10.2136/sssaj1998.03615995006200040031x [5] M. Weih and P. S. Karlsson, “Growth Response of Mountain Birch to Air and Soil Temperature: Is Increas- ing Leaf-Nitrogen Content an Acclimation to Lower Air Temperature?” New Phytologist, Vol. 150, No. 1, 2001, pp. 147-155. doi:10.1046/j.1469-8137.2001.00078.x [6] M. Stromgren and S. Linder, “Effects of Nutrition and Soil Warming on Stemwood Production in a Boreal Nor- way Spruce Stand,” Global Change Biology, Vol. 8, No. 12, 2002, pp. 1195-1204. doi:10.1046/j.1365-2486.2002.00546.x [7] A. Gessler, S. Schneider, D. Von Sengbusch, P. Weber, U. Hanemann, C. Huber, A. Rothe, K. Kreutzer and H. Ren- nenberg, “Field and Laboratory Experiments on Net Up- take of Nitrate and Ammonium by the Roots of Spruce (Picea abies) and Beech (Fagus sylvatica) Trees,” New Phytologist, Vol. 138, No. 2, 1998, pp. 275-285. doi:10.1046/j.1469-8137.1998.00107.x [8] M. L. Adam, J. M. Kelly, W. R. Graves, and P. M. Dixon, “Net Nitrate Uptake by Red Maple Is a Function of Root-Zone Temperature,” Journal of Plant Nutrition, Vol. Copyright © 2011 SciRes. JEP  Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature 776 26, No. 1, 2003, pp. 203-222. doi:10.1081/PLN-120016505 [9] J. M. Kelly, “Temperature Affects Solution-Phase Nutri- ent Concentrations and Subsequent Calculation of Supply Parameters,” Soil Science Society of America Journal, Vol. 57, 1993, pp. 527-531. doi:10.2136/sssaj1993.03615995005700020038x [10] S. A. Barber, “Soil Nutrient Bioavailability: A Mechanist Approach,” 2nd Edition, John Wiley & Sons, Inc., Ho- boken, 1995. [11] S. C. Sheppard and G. J. Racz, “Effects of Soil Tempera- ture on Phosphorus Extractability. I. Extractions and Plant Uptake of Soil and Fertilizer Phosphorus,” Canadian Journal of Soil Science, Vol. 64, No. 2, 1984, pp. 241-254. doi:10.4141/cjss84-025 [12] K. Sardi, P. Csatho and E. Osztoics, “Changes in the Soil Phosphorus Content of a Long-Term Fertilization Field Trial Studied in Laboratory Incubations,” Communica- tions in Soil Science and Plant Analysis, Vol. 37, No. 15-20, 2006, pp. 2833-2840. [13] K. Simpson, “Factors Influencing Uptake of Phosphorus by Crops in Southeast Scotland,” Soil Science, Vol. 92, 1961, pp. 1-14. doi:10.1097/00010694-196107000-00002 [14] K. Oats and S. A. Barber, “Nutrient Uptake: A Mini- computer Program to Predict Nutrient Absorption from Soil by Roots,” Journal of Agronomic Education, Vol. 16, 1987, pp. 65-68. [15] A. R. Gillespie and P. E. Pope, “Rhizosphere Acidifica- tion Increases Phosphorus Recovery of Black Locust. II. Model Predictions and Measured Recovery,” Soil Science Society of America Journal, Vol. 54, No. 2, 1990, pp. 538-541. doi:10.2136/sssaj1990.03615995005400020042x [16] K. C. J. Van Rees, N. B. Comerford and W. W. McFee, “Modeling Potassium Uptake by Slash Pine Seedlings from Low-Potassium-Supplying Soils,” Soil Science So- ciety of America Journal, Vol. 54, No. 5, 1990, pp. 1413-1421. doi:10.2136/sssaj1990.03615995005400050034x [17] J. M. Kelly, A. H. Chappelka and B. G. Lockaby, “Meas- ured and Estimated Parameters for a Model of Nutrient Uptake by Trees,” New Zealand Journal of Forest Sci- ence, Vol. 24, 1994, pp. 213-225. [18] N. Claassen and B. Steingrobe, “Mechanistic Simulation Models for a Better Understanding of Nutrient Uptake from Soil,” In: Z. Rengel, Ed., Mineral Nutrition of Crops: Fundamental Mechanisms and Implications, Food Prod- ucts Press, New York, 1999, pp. 327-367. [19] R. D. Hangs, J. D. Knight and K. C. J. Van Rees, “Nitro- gen Uptake Characteristics for Roots of Conifer Seedlings and Common Boreal Forest Competitor Species,” Cana- dian Journal of Forest Research, Vol. 33, No. 1, 2003, pp. 156-163. doi:10.1139/x02-169 [20] J. M. Kelly and P. A. Mays, “Root Zone Physical and Chemical Characteristics in Southeastern Spruce-Fir Stands,” Soil Science Society of America Journal, Vol. 53, No. 4, 1989, pp. 1248-1255. doi:10.2136/sssaj1989.03615995005300040043x [21] F. C. Thornton, P. A. Pier and C. McDuffie Jr., “Re- sponse of Growth, Photosynthesis, and Mineral Nutrition of Red Spruce Seedlings to Ozone and Acidic Cloud Deposition,” Environmental and Experimental Botany, Vol. 30, No. 3, 1990, pp. 313-323. doi:10.1016/0098-8472(90)90043-4 [22] J. M. Kelly, S. A. Barber and G. S. Edwards, “Modeling Magnesium, Phosphorus and Potassium Uptake by Lob- lolly Pine Seedlings Using a Barber-Cushman Approach,” Plant and Soil, Vol. 139, No. 2, 1992, pp. 209-218. doi:10.1007/BF00009312 [23] J. D. Joslin and M. H. Wolfe, “Red Spruce Soil Solution Chemistry and Root Distribution across a Cloud Water Deposition Gradient,” Canadian Journal of Forest Re- search, Vol. 22, No. 6, 1992, pp. 893-904. doi:10.1139/x92-119 [24] R. Parsons, “Handbook of Electro-Chemical Constants,” Academic Press, Waltham, 1959. [25] O. W. Edwards and E. O. Huffman, “Diffusion of Aque- ous Solutions of Phosphoric Acid at 25˚,” Journal of Physical Chemistry, Vol. 63, 1959, pp. 1830-1833. doi:10.1021/j150581a011 [26] J. D. Joslin and M. H. Wolfe, “Foliar Deficiencies of Mature Southern Appalachian Red Spruce Determined from Fertilizer Trials,” Soil Science Society of America Journal, Vol. 58, No. 5, 1994, pp. 1572-1579. doi:10.2136/sssaj1994.03615995005800050042x [27] J. M. Kelly, J. D. Scarbrough and P. A. Mays, “Hard- wood Seedling Root and Nutrient Parameters for a Model of Nutrient Uptake,” Journal of Environmental Quality, Vol. 30, 2001, pp. 427-439. doi:10.2134/jeq2001.302427x [28] R. D. Yanai, K. J. McFarlane, M. S. Lucash, S. E. Kulpa and D. M. Wood, “Similarity of Nutrient Uptake and Root Dimensions of Engelmann Spruce and Subalpine Fir at Two Contrasting Sites in Colorado”, Forest Ecology and Management, Vol. 258, No. 10, 2009, pp. 2233-2241. doi:10.1016/j.foreco.2009.04.035 [29] T. D. Perkins, G. T. Adams, S. T. Lawson, P. G. Schab- erg and S. G. McNulty, “Long-Term Nitrogen Fertiliza- tion Increases Winter Injury in Montane Red Spruce (Picea Rubens) Foliage,” Journal of Sustainable Forestry, Vol. 10, No. 1, 2000, pp. 165-172. doi:10.1300/J091v10n01_19 [30] S. Catovsky and F. A. Bazzaz, “Nitrogen Availability Influences Regeneration of Temperate Tree Species in the Understory Seedling Bank,” Ecological Applications, Vol. 12, No. 4, 2002, pp. 1056-1070. doi:10.1890/1051-0761(2002)012[1056:NAIROT]2.0.CO ;2 [31] M. Silberbush and S. A. Barber, “Sensitivity of Simulated Phosphorus Uptake to Parameters Used by a Mechanis- tic-Mathematical Model,” Plant and Soil, Vol. 74, No. 1, 1983, pp. 93-100. doi:10.1007/BF02178744 [32] H. Van Miegroet, D. W. Cole and N. W. Foster, “Nitro- gen Distribution and Cycling,” In: D. W. Johnson and S. Copyright © 2011 SciRes. JEP  Model Estimates of Nutrient Uptake by Red Spruce Respond to Soil Temperature Copyright © 2011 SciRes. JEP 777 E. Lindberg, Eds., Atmospheric Deposition and Nutrient Cycling in Forest Ecosystems of the Integrated Forest Study, Springer-Verlag, Berlin, 1992, pp. 178-196. [33] J. D. Joslin, J. M. Kelly and H. Van Miegroet, “Soil Chemistry and Nutrition of North American Spruce-Fir Stands: Evidence for Recent Change,” Journal of Envi- ronmental Quality, Vol. 21, No. 1, 1992, pp. 12-30. doi:10.2134/jeq1992.00472425002100010002x [34] K. F. Nielsen, R. L. Halstead, A. J. MacLean, R. M. Holmes and S. J. Bourget, “The Influence of Soil Tem- perature on the Growth and Mineral Composition of Oats,” Canadian Journal of Soil Science, Vol. 40, No. 2, 1960, pp. 255-263. doi:10.4141/cjss60-032 [35] C. J. Whitfield and D. E. Smika, “Soil Temperature and Residue Effects on Growth Components and Nutrient Uptake of Four Wheat Varieties,” Agronomy Journal, Vol. 68, 1971, pp. 297-300. doi:10.2134/agronj1971.00021962006300020029x [36] P. Hogberg, L. Hogbom and H. A. Schinkel, “Nitro- gen-Related Root Variables of Trees along an N-Deposition Gradient in Europe,” Tree Physiology, Vol. 18, No. 12, 1998, pp. 823-828. [37] C. G. Del Valle and S. A. Harmon, “Turnip Growth and Mineral Composition as Influenced by Soil Temperature and Two Fertility Levels,” Proceedings American Society for Horticultural Science, Vol. 92, 1968, pp. 578-582. [38] K. F. Nielsen and R. K. Cunningham, “The Effects of Soil Temperature and Form and Level of Nitrogen on Growth and Chemical Composition of Italian Ryegrass,” Soil Science Society of America Proceedings, Vol. 28, 1964, pp. 213-218. doi:10.2136/sssaj1964.03615995002800020026x [39] E. H. DeLucia, S. A. Heckathorn and T. A. Day, “Effects of Soil Temperature on Growth, Biomass Allocation and Resource Acquisition of Andropogon gerardii Vitman,” New Phytologist, Vol. 120, No. 4, 1992, pp. 543-549. doi:10.1111/j.1469-8137.1992.tb01804.x [40] H. Bassirirad, “Kinetics of Nutrient Uptake by Roots: Responses to Global Change,” New Phytologist, Vol. 147, No. 1, 2000, pp. 155-169. doi:10.1046/j.1469-8137.2000.00682.x
|