Journal of Electromagnetic Analysis and Applications
Vol. 3 No. 2 (2011) , Article ID: 4128 , 10 pages DOI:10.4236/jemaa.2011.32012
Static and 50 Hz Electromagnetic Fields Effects on Human Neuronal-Like Cells Vibration Bands in the Mid-Infrared Region
![]()
1Department of Physics, University of Messina, Viale D’Alcontres, Messina, Italy; 2Department of Biochemical Physiological and Nutritional Sciences, University of Messina, Messina, Italy.
Email: e.calabro@yahoo.com
Received January 17th, 2011; revised February 18th, 2011; accepted February 21st, 2011.
Keywords: Static Magnetic Field, 50 Hz electromagnetic field, Neuronal-like cells, FTIR spectroscopy
ABSTRACT
Human neuronal-like cells were exposed to static and 50 Hz electromagnetic fields at the intensities of 2 mT and 1 mT, respectively. The effects of exposure were investigated in the mid-infrared region by means of Fourier self deconvolution spectroscopic analysis. After exposure of 3 hours to static and 50 Hz electromagnetic fields, the vibration bands of CH2 methilene group increased significantly after both exposures, suggesting a relative increase of lipid related to conformational changes in the cell membrane due to electromagnetic fields. In addition, PO2- stretching phosphate bands decreased after both exposures, suggesting that alteration in DNA/RNA can be occurred. In particular, exposure of 3 hours to 50 Hz electromagnetic fields produced significant increases in β-sheet contents in amide I, and around the 1740 cm−1 band assigned to non-hydrogen-bonded ester carbonyl stretching mode, that can be related to unfolding processes of proteins structure and cells death. Further exposure up to 18 hours to static magnetic field produced an increase in β-sheet contents as to α-helix components of amide I region, as well.
1. Introduction
Epidemiological studies carried out up to now evidenced a relationship between an increase of risk of cancer and the exposure to extremely low frequency electromagnetic field (ELF-EMF) [1-3].
Otherwise, a few studies have examined possible effects of exposure to static magnetic field (SMF), particularly in relation to cancer, but the results have been inconclusive [4]. However, the International Commission on Non-Ionizing Radiation Protection (ICNIRP) recommended an exposure limit of 40 mT for the general public [5].
SMFs are generated in electric trains, where measurements performed by references [6,7] reported SMF values up to 15 mT.
Measurements by the National Institute for Occupational Safety and Health (NIOSH) in various industries provided that electro-galvanizing processes and aluminium production can produce SMFs up to 460 µT [8].
Public exposure to small artificial sources of SMF generally occurs from audio speakers components or battery-operated motors. We measured that the small magnets of headphones can produce SMFs up to 10 mT within two centimetres of their magnetic poles.
Furthermore, SMFs are usually used in research and in medical applications, such as magnetic resonance imaging, where scanned patients can be exposed to very strong magnetic fields up to 10 T.
In the vicinity of certain home appliances, the magnetic-field intensities due to ELF-EMFs can be as much as a few hundred microtesla, whereas in some workplaces can reach 10 mT, leading us to investigate the range of exposure to magnetic field around a few millitesla.
Exposure to ELF-EMFs from transmission and distribution lines has been proposed as a possible explanation for the association with childhood leukaemia [9].
In spite of human and animal studies are of greater importance than cellular models for health risk evaluation, they cannot usually be used to investigate mechanistic events underlying the carcinogenesis processes.
In contrast, cellular or in vitro models can be useful for the investigation of many of the numerous molecular aspects of carcinogenesis.
Some studies have shown no genotoxic effects after exposure to ELF-EMFs in several types of mammalian cells [10-12]. In contrast, other studies have shown genotoxicity in cellular models [13-15].
Reference [16] reported increased cell proliferation, changes in cell cycle and increased DNA damage, in HL-60 leukaemia cells exposed to 50 Hz magnetic fields at 0.5-1 mT up to 72 hours.
Reference [17] exposed human lymphocytes to 50 Hz magnetic fields. Lymphocytes were first exposed to the ELF field for 24 h at 80 or 800 µT, and significant increases in the frequency of micronuclei and apoptosis were found.
Reference [18] investigated the effect of 1 h or 24 h exposure of Jurkat cells to a 5 mT, 50 Hz EMF, either alone, or with two genotoxic metabolites. No effect of 1 h exposure to magnetic fields was observed, whereas exposure for 24 h caused increases in micronuclei.
In further studies it was found that the maximum of damage was obtained in fibroblasts after 15-19 h of exposure to ELF-EMFs. After the peak of the damage, the effect declined within the next hours [19].
Effects of exposure of human neuroblastoma cells to a 50 Hz EMF at 1 mT were investigated by [20]. They proved that a 24 h exposure significantly increased cell proliferation (+10%) and a 72 h exposure delayed the retinoic-acid-induced differentiation through increased cell proliferation and decreased expression of the B-myb protein.
Reference [21] studied the effects of ELF-EMF in synovial fibroblasts and neuroblastoma cells. No effects were found in nerve cells, but a decrease in the conductance of gap junction channels under exposure to 20 mA/m2 at 60 Hz and a significant increase in intracellular Ca2+ at current densities of more than 10 mA/m2 were found.
Reference [22] exposed three malignant human cell lines (melanoma, ovarian carcinoma and lymphoma) to a 7 T uniform magnetic field for 64 hours. Exposure reduced the number of viable cells in each cell line by 20% to 40%.
Reference [23] exposed normal human neuronal cell cultures to a strong SMF of 0.2 T for a short period of 15 minutes. A change in morphology incorporation was observed indicating decreased DNA synthesis and consequent inhibition of proliferation. In contrast, non neuronal cells, showed no alteration following exposure.
Reference [24] showed that the exposure of a cellulose membrane to a SMF of 0.24 T produces a significantly enhanced rate of potassium ion transport which did not return to the initial basal level after exchange of the aqueous medium, suggesting that an irreversible change took place on the cellulose membrane.
These results led us to investigate the effects of SMFs and ELF-EMFs on human neuronal-like cells by means of Fourier Transform Infrared (FTIR) Spectroscopy, considering exposure intensity levels lesser than the exposure limits recommended by [5].
The literature cited above provided characteristic parameters of time-exposures and EMF intensities of 3-18 hours and 1-2 mT, respectively, that were used in our study.
2. Materials and Methods
2.1. Cell culture and Treatment
Human neuronal-like cells were cultured in a 1:1 mixture of MEM and Ham’s F12 medium containing 10% (v/v) heat inactivated FBS, L-glutamine (2 mM), sodium pyruvate (1 mM) and maintained at 37˚C in a humidified incubator with 5% CO2 and 95% air.
Sub-confluent cells were washed twice with PBS then incubated in MEM/Ham’s F12 medium containing 10 µM RA (10 mM in DMSO stock solution), 1% FBS, L-glutamine (2 mM), sodium pyruvate (1 mM). The medium was renewed every two days.
After 5 days of 10µM RA exposure, differentiated SHSY5Y cells were exposed to magnetic field.
2.2. Experimental Design
The exposure system consisted of a couple of Helmholtz coils, with pole pieces of round parallel polar faces, to produce a uniform magnetic field at the center of the coils distance.
The coils were located into a incubator in a 5% CO2/ 95% air humidified at the temperature of (37.1 ± 0.1)˚C.
A SMF was generated by the Helmholtz coils couple powered by a DC generator. Uniform intensity fields up to 2 mT were produced, much lesser than the exposure limit for SMF of 40 mT, recommended for public exposure by [5].
Time-varying electromagnetic fields at the frequency of 50 Hz was generated by means of a AC voltage regulating up to 230 volt, which enabled us to provide the magnetic flux density of 1 mT between the polar faces of the coils.
Samples of human neuronal-like cells were placed at the center of the generated field between the coils located into the incubator (Figure 1).
Control samples were placed into another incubator at the same physical conditions, maintained rigorously at the same values of air humidified incubator and temperature during the entire exposure.
The magnetic field intensity was continuously monitored by a magnetic field probe GM07 Gaussmeter of HIRST-Magnetic Instruments Ltd – UK (Figure 2).

Figure 1. The exposure setup used to expose neuronal-like cells to SMF and to 50 Hz EMF.

Figure 2. The magnetic field probe GM07 Gaussmeter (HIRST-Magnetic Instruments Ltd–UK) used for monitoring exposure to SMF and 50 Hz EMF.
2.3. Infrared Spectroscopy
For FTIR analysis after exposure, neuronal-like cells (5 × 105 cells/ml in 25 cm2 culture flask) were washed twice with PBS (pH 7.4), harvested with non-enzymatic cell dissociation solution, and resuspended in 50 µl of PBS. Then, this volume was transferred to a slide and air-dried for readings in the spectrometer Vertex 80 v from Bruker Optics.
Analogue techniques to disaggregate cells from the culture medium were used by [25,26].
The attenuated total reflection (ATR) method was chosen for spectrum collection.
The use of ATR for cells spectra collection in FTIR was enhanced by [27] because it regardless of sample thickness.
References [28,29] showed that the absorption in the spectral region assigned to nucleic acids can change following apoptosis. Reference [30] confirmed that ATR-FTIR spectroscopy can be successful used for analysis and quantification of cells apoptosis.
Furthermore, ATR technique is the desired method to overcome solvent masking since the penetration depth of infrared light is inherently limited to a fraction of the wavelength estimated to be λ/10, permitting rapid secondary structure analysis on small volumes [31].
For each spectrum 64 interferograms were collected and co-added by Fourier transformed employing a HappGenzel apodization function to generate a spectrum with a spectral resolution of 4 cm–1 in the range from 7000 cm–1 to 1000 cm–1.
IR spectra of air were subtracted from acquired spectra at the corresponding temperature to eliminate minor spectral contributions due to residual water vapor. However, a smoothing correction for atmospheric water background was performed.
The IR spectra were baseline corrected by means of automatic baseline scattering correction function, to subtract baselines from spectra, providing spectra with band edges of up to the theoretical baseline.
The spectra were successively area normalized for exposed cells and control samples and vector normalization was used, calculating the average value of the spectrum and subtracting from the spectrum decreasing the midspectrum. The sum of the squares of all values was calculated and the spectrum divided by the square root of this sum.
In addition, interactive baseline rubberband correction was used to minimize the water band contribution to spectra. This method also uses a rubberband which is stretched from one spectrum end to the other, and the band is pressed onto the spectrum from the bottom up with varying intensity. This method performs iteratively, depending on the number of iterations in the algorithm and the baseline as a frequency polygon consisting of n baseline points. The resulting spectrum will be the original spectrum minus the baselines points manually set and a subsequent concave rubberband correction. We used the default value of n = 64 baseline points and a number of 60 iterations.
ATR spectra were smoothed by the Loess algorithm and the deconvolved spectra fitted with Gaussian band profiles. Initial values for the peak heights and widths were estimated from the deconvolved spectra.
In order to enhance fine spectral features, Fourier SelfDeconvolution (FSD) analyses of infrared spectra were performed.
The aim of FSD is to enhance the apparent resolution of a spectrum, or to decrease the line width. Spectral ranges comprising broad and overlapping lines can thus be separated into sharp single lines.
The following parameters were used: Lorentzian shape, bandwidth = 11.05, deconvolution factor = 2, noise reduction factor = 0.5.
3. Results and Discussion
Mid-Infrared spectroscopy using FTIR spectrometers is a consolidated analytical method for the characterization of molecular species in biological systems. Deviations from characteristic vibration spectra of cells or tissues can be due to the presence of abnormalities produced by external stress factors or disease states. Hence this technique was used to investigate the effects of exposure of neuronallike cells to SMFs and ELF-EMFs in the mid-infrared spectral region.
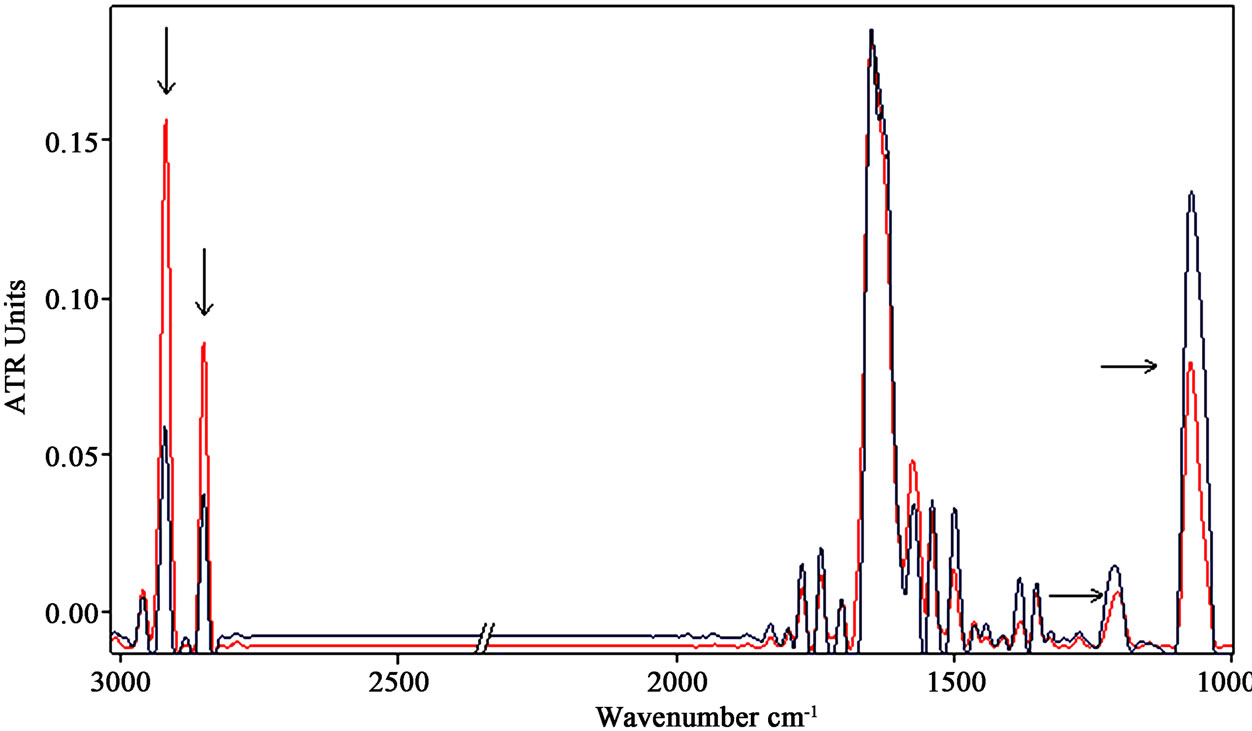
Typical spectra of cells exposed and not exposed to a SMF of 2 mT are shown in Figure 3 (a,b) after exposures of 3 h and 18 h, respectively. Significant changes in the exposed cells spectra were evidenced after 3 h of exposure to SMF in the region from 2800 to 3000 cm−1, where an increase of both stretching vibration bands of CH2 at 2925 cm–1 and 2853 cm–1 occurred after exposure (Figure 3 (a)).The bands at 2925 cm-1 and 2853 cm–1 are due to asymmetric asCH2 and symmetric sCH2 stretching of methylene group, respectively, relative to the presence of lipids or proteins [32,33].
Bending vibration bands of CH2 group at lower energies were observed to increase, as well.
Applying a statistical analysis (t-test) to the integrated area of sCH2 stretching vibration bands of different spectra, we found that their increase was significantly different in comparison to controls (p < 0.001).
The observed increase of CH2 content after exposure of 3 h to a SMF at 2 mT can be performed by an increase of the lipid content in the cellular environment.
The molecular mechanism underlying the CH2 increase may be related to a slight decrease in cellular volume following extensive growth conditions [34,35].
In fact, a relative increase in the surface area of the cellular membrane can produce an augmentation of the phospholipids fraction in the biomass, corresponding to a major enrichment in CH2 relative to CH3 groups.
Further changes were observed around 1230 and 1080 cm–1 vibration bands after exposure, indicated by arrows in Figure 3 (a).
They can be assigned to the phosphate absorption bands corresponding to the asymmetric stretching phosphate mode asPO2- and symmetric stretching phosphate mode sPO2- of phosphodiester bonds in nucleic acids, respectively [32,33].
The ratio between the integrated area of asymmetric and symmetric stretching PO2- bands of exposed samples and that relative to control samples decreased significantly (P < 0.01), suggesting that some alteration in DNA/RNA can be occurred after exposure to SMF.
Longer exposure up to 18 h to SMF provided lesser increase of CH2 stretching vibration bands, as can be observed in (Figure 3 (b)).
 (a)
(a)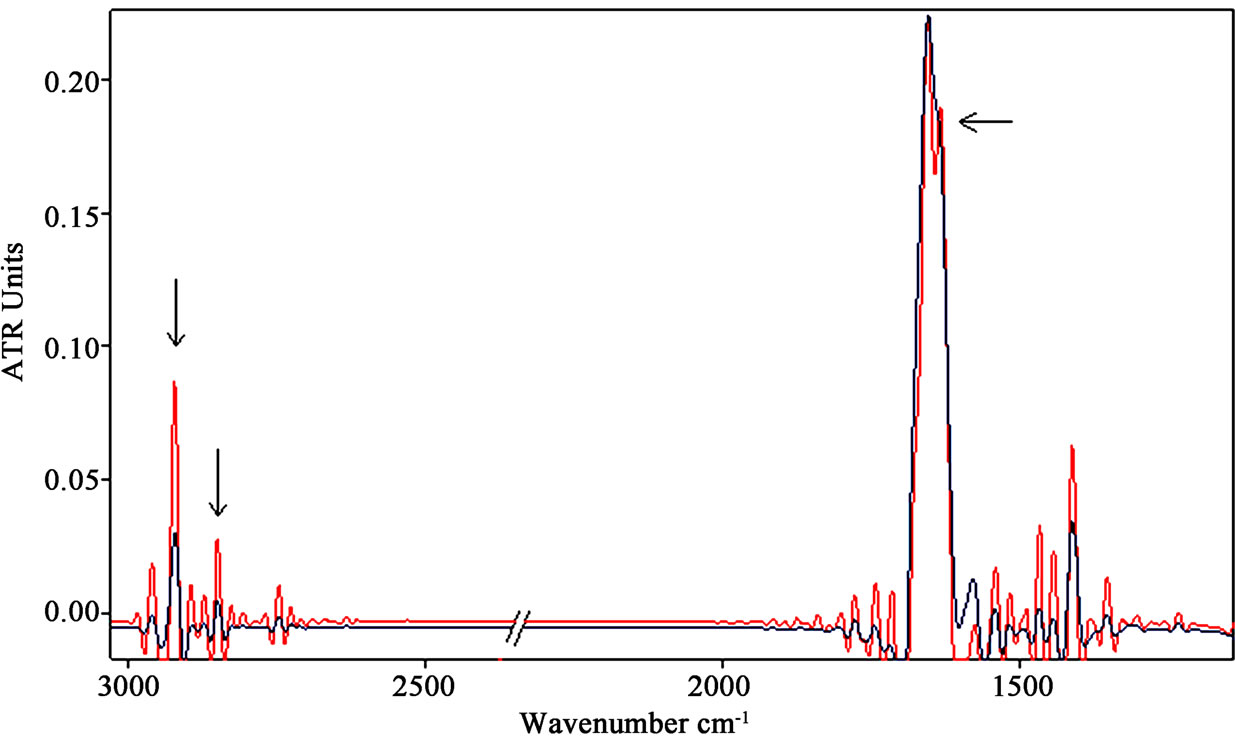 (b)
(b)
Figure 3. (a) Representative infrared spectra of neuronal-like cells samples after exposure of 3 h to a SMF at the intensity of 2 mT (the spectrum of the exposed sample is represented by the red line). The increases of CH2 stretching vibration bands are indicated by arrows. The decreases of PO2 bands are indicated, as well. (b) The increase of the β-sheet content at 1635 cm–1 in amide I region was observed after 18 h of exposure.
Nevertheless, the spectral analysis after 18 h of exposure revealed a shift of the peaks of amide I and II regions and an increase of the β-sheet contents at 1635 cm–1 (indicated by arrow in Figure 3(b)) comparing exposed and not exposed spectra.
The amide I band centered at about 1645 cm−1 (which arises from the amide C═O stretching vibration and N-H bending mode) and the amide II band, centered around 1545 cm−1 (arising from vibration modes of C–N–H bending and C–N stretching due to peptide groups in all proteins) provide main informations about the overall protein secondary structure in the cells [36].
The amide I and II regions after exposure of 18 h to SMF were zoomed in and represented in Figure 5(a), where the relative increase of the β-sheet contents at 1635 cm-1 with respect to the α-helix component, significantly different in comparison to control (p < 0.05), was enhanced and indicated by arrow.
This result indicated a change in the overall protein conformational state within the cell, that could be due to a denaturation of proteins during apoptosis or, alternatively, to different distribution of proteins and unfolding process and formation of aggregates [37].
Exposure of 3 h of neuronal-like cells to a 50 Hz EMF at the magnetic flux density of 1 mT was carried out.
A representative spectrum in the mid-infrared region is represented in Figure 4.
It appears that the intensity of both stretching vibration bands of CH2 at 2925 cm−1 and 2853 cm−1 increased strongly and significantly for exposed samples (p < 0.001).
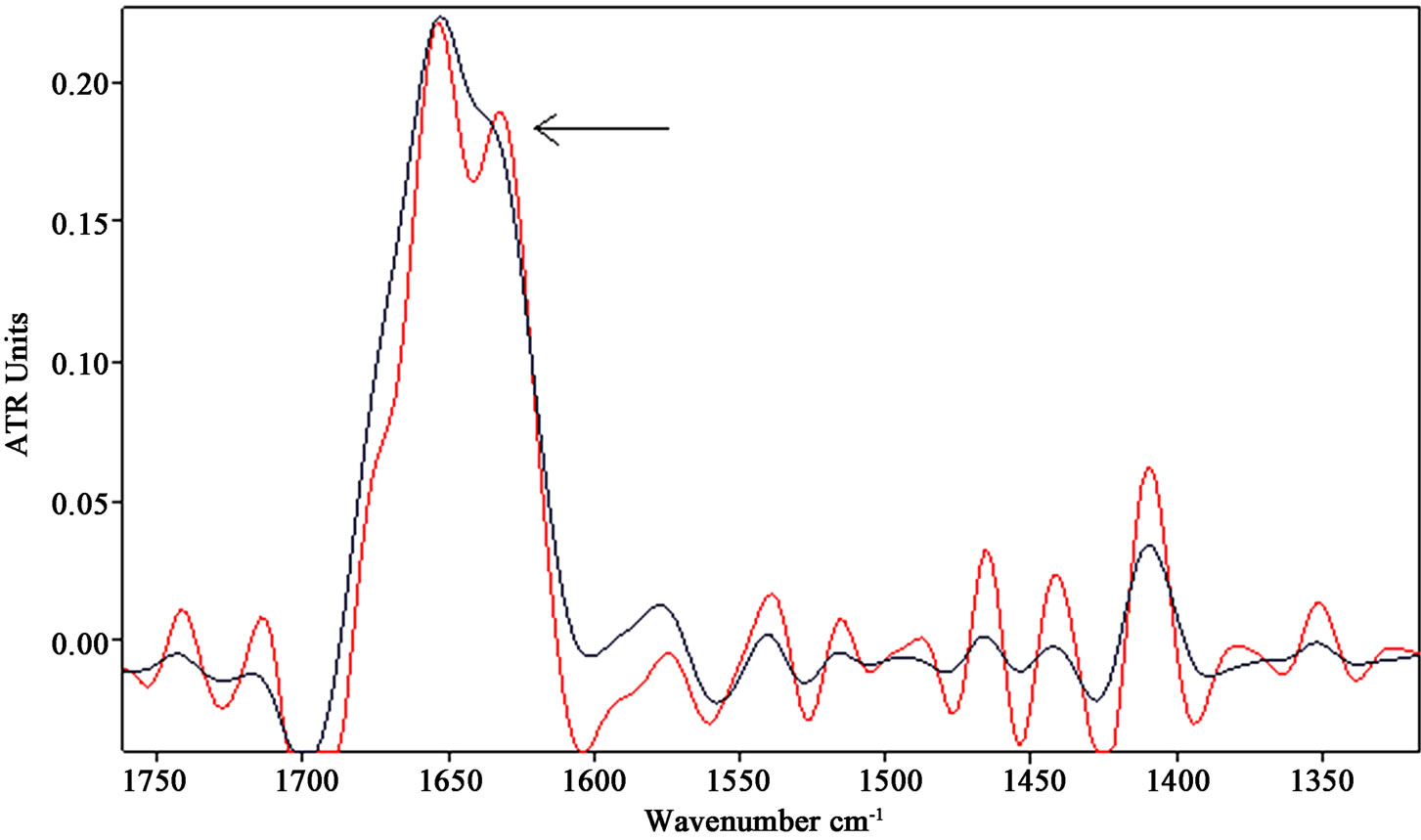
The analysis of exposed spectra in the amide I region revealed a loss of the α-helical and short segment connecting α-helix segments content and an increase of the β-sheet component at 1635 cm−1 relative to the α-helix, as can be clearly observed in Figure 5(b), where the amide I and II regions were zoomed in.
Applying a statistical analysis (t-test) to different exposed and control samples, the β-sheet/α-helix ratio of the integrated areas (exposed/control) of their vibration bands provided an average value of 1.29/1.13, which demonstrated a significant increase of the β-sheet content with respect to the α-helix in the proteins secondary structure (p < 0.05).
Another relevant change observed in the mid-infrared region after the exposure to ELF-EMF is represented by the increase in intensity of the band around 1740 cm-1, indicated by arrows in Figure 4 and Figure 5(b). This band can be assigned to non-hydrogen-bonded ester carbonyl C═O stretching mode within phospholipids, whose increase gives further evidence of a dying cell state as suggested by [38,39].
The integrated area of 1740 cm–1 band of exposed samples increased significantly with respect to control after exposure (p < 0.001).
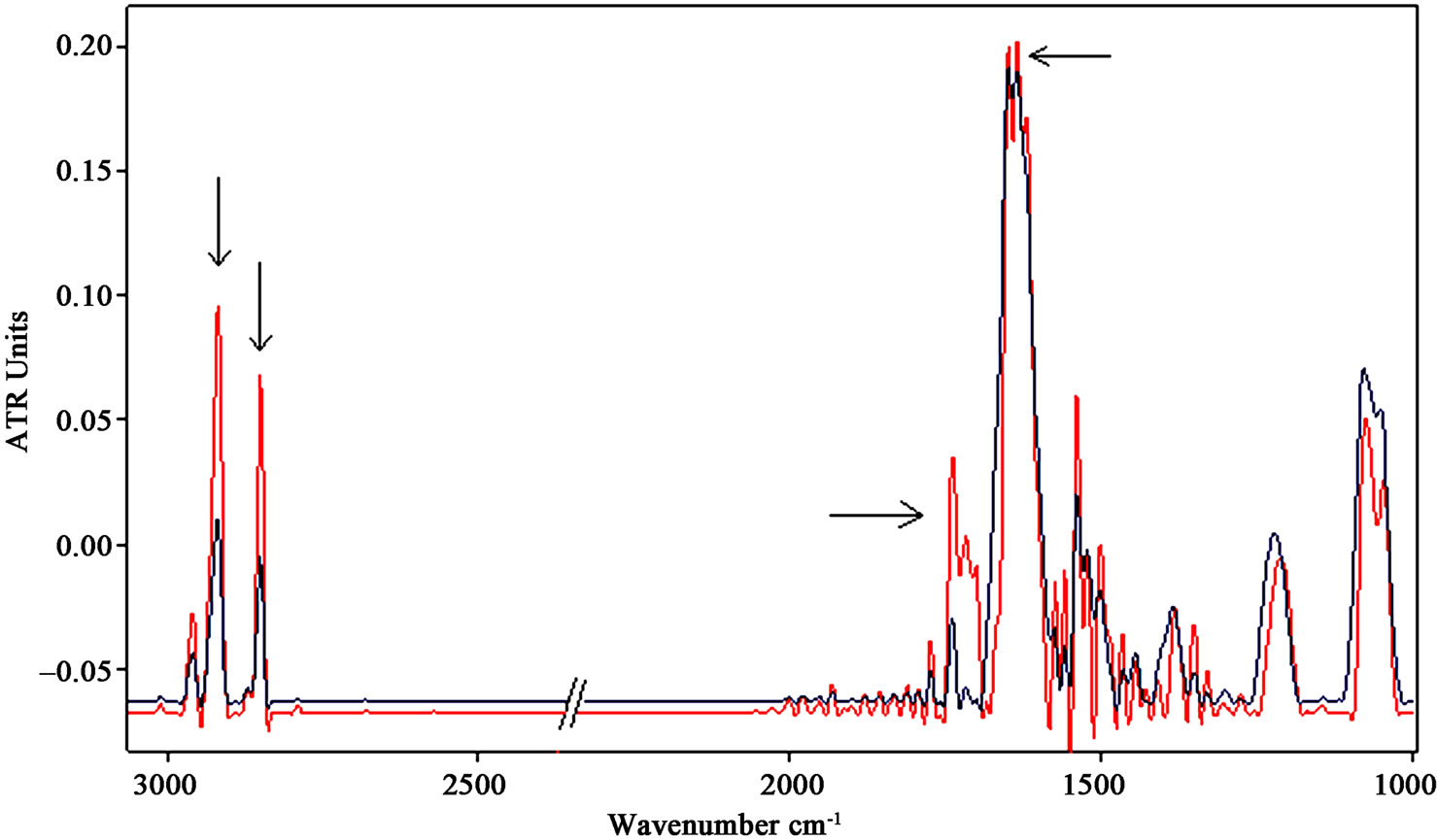
Figure 4. Representative infrared spectra of neuronal-like cells samples after exposure of 3 h to 50 Hz EMF at the intensity of 1 mT (the spectrum of the exposed sample is represented by the red line). The increases of CH2 stretching vibration bands are indicated by arrows. The increases of the β-sheet content in amide I and of the non-hydrogen-bonded ester carbonyl C═O stretching band around 1740 cm–1 indicate a relevant change in the overall protein conformational states within the cell.
 (a)
(a)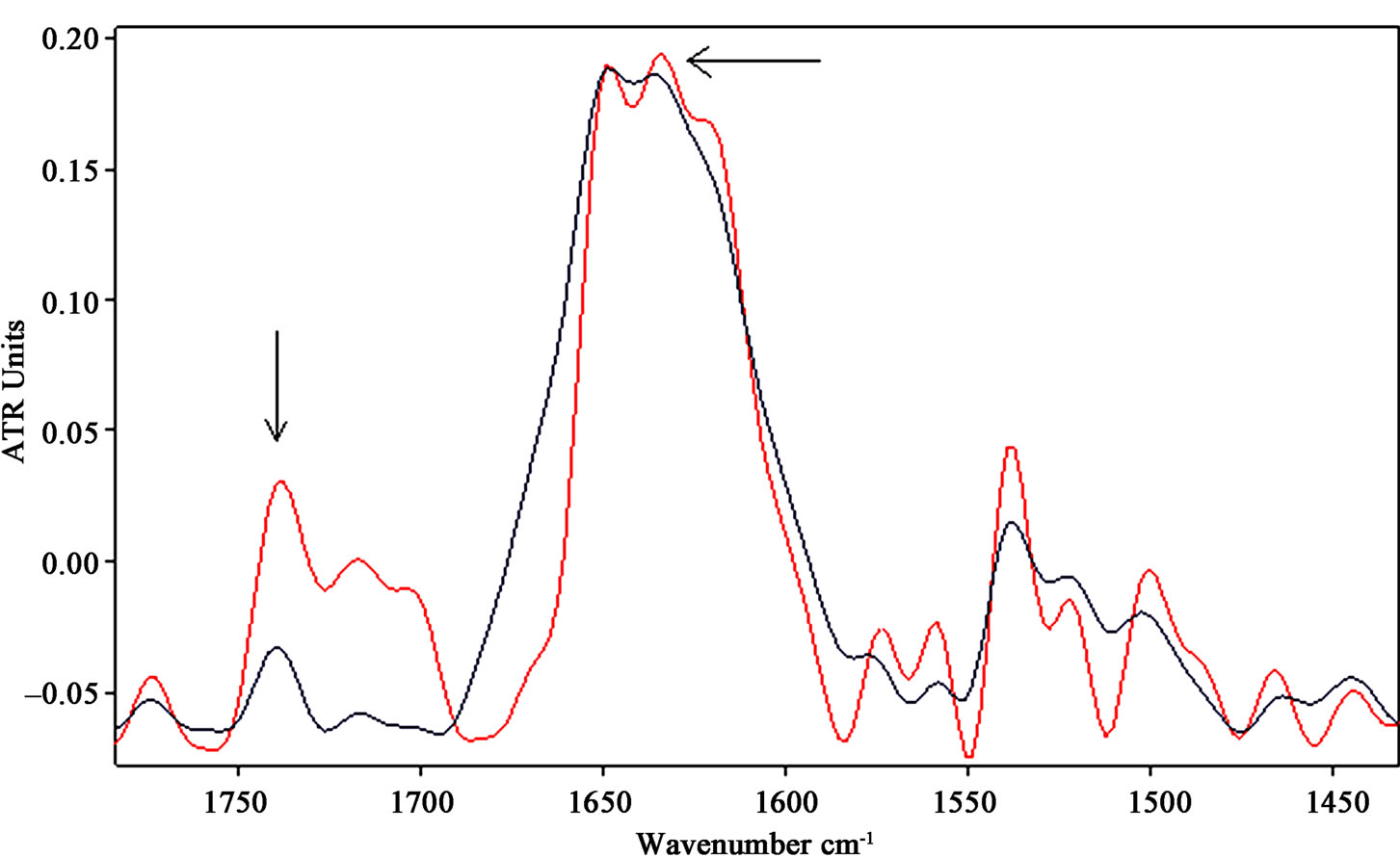 (b)
(b)
Figure 5. (a) Representative FSD spectra in the amide I and II regions of neuronal-like cells samples after exposure to SMF. The relative increase of the β-sheet content as to the α-helix component in amide I is enhanced and indicated by arrow. (b) Representative FSD spectra in the amide I and II regions of neuronal-like cells samples after exposure to 50 Hz EMF. Besides the increase of the β-sheet content in amide I, the increase around 1740 cm-1 (associated with the non-hydrogen-bonded ester carbonyl C═O stretching band within phospholipids) was evidenced.
This circumstance can be performed as the C═O ester carbonyl groups of lipids in the cell are becoming predominantly non-hydrogen bonded, which would be in agreement with an eventual oxidative damage having occurred [40,41].
Previous FTIR spectroscopic analysis of dying cells has shown that two main characteristic spectral signatures can be assumed as indicative of death [42,43]: the increase of the amount of β-sheet content in the secondary structures around 1630 cm−1 (that produces the shift down of the amide I and II peak’s centroid) and the appearance of a peak around 1740 cm−1, which can indicate a change in the overall protein conformational states within the cell. Both effects occurred after exposure to ELF-EMFs.
4. Conclusions
The effects of static magnetic field and 50 Hz electromagnetic field on neuronal-like cells were investigated in the mid-infrared spectral region by means of FTIR spectroscopy in Fourier self-deconvolution.
Exposure of 3 hours to a uniform magnetic field at the intensity of 2 mT produced a significant increase in intensity of CH2 methylene group vibration bands, which may be associated with an increase of lipid content related to an increase of the surface of the cellular membrane. In addition, a decrease of the PO2- phosphate bands was observed, suggesting that alteration in DNA/ RNA can be occurred after exposure. Further exposure up to 18 hours produced an increase of the β-sheet/α-helix ratio in amide I region due to unfolding processes of proteins.
Exposure of 3 hours to a 50 Hz electromagnetic field at the magnetic flux density of 1 mT provided that signifycant increases of the amount of β-sheet content and of the band around 1740 cm−1 (assigned to non-hydrogenbonded ester carbonyl stretching mode) occurred after exposure, which may be considered as the main characteristic spectral signatures assumed as indicative of cells death.
However, further research is needed to understand the relationship of metabolism and cellular functions with exposure to electromagnetic magnetic fields.
REFERENCES
- WHO (World Health Organization), “Extremely Low Frequency (ELF) Fields,” Environmental Health Criteria, Geneva, Vol. 35, 1984.
- WHO (World Health Organization), “Magnetic Fields,” Environmental Health Criteria, Geneva, Vol. 69, 1987.
- WHO (World Health Organization), “Electromagnetic Fields (300 Hz to 300 GHz),” Environmental Health Criteria, Geneva, Vol. 137, 1993.
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans, “Non-Ionizing Radiation, Part 1: Static and Extremely Low-Frequency (ELF) Electric and Magnetic Fields,” IARC (Monographs on the Evaluation of Carcinogenic Risks to Humans), Lyon, 2002, Vol. 80, pp. 1-395.
- ICNIRP (International Commission on Non-Ionizing Radiation Protection), “Guidelines for Limiting Exposure to Time-Varying Electric, Magnetic, and Electromagnetic Fields (up to 300 GHz),” Health Physics, Vol. 74, No. 4, 1998, pp. 494-522.
- P. Chadwick and F. Lowes, “Magnetic Fields on British Trains,” Annals of Occupational Hygiene, Vol. 42, No. 5, 1998, pp. 331-335.
- F. M. Dietrich and W. L. Jacobs, “Survey and Assessment of Electric and Magnetic Field (EMF) Public Exposure in the Transportation Environment,” US Department of Transportation, Federal Railroad Administration, 1999 (Report No. PB 99-130908).
- NIOSH (National Institute for Occupational Safety and Health), National Institute of Environmental Health Sciences and US Department of Energy, “Questions and answers—EMF in the Workplace. Electric and Magnetic Fields Associated with the Use of Electric Power,” EMF RAPID Program, Washinton DC, 1996.
- S. Milham and E. M. Ossiander, “Historical Evidence that Residential Electrification Caused the Emergence of the Childhood Leukemia Peak,” Medical Hypotheses, Vol. 56, No. 3, 2001, pp. 290-295. doi:10.1054/mehy.2000.1138
- J. A. Reese, R. F. Jostes and M. E. Frazier, “Exposure of Mammalian Cells to 60-Hz Magnetic or Electric Fields: Analysis for DNA Singles-Strand Breaks,” Bioelectromagnetics, Vol. 9, No. 3, 1988, pp. 237-247. doi:10.1002/bem.2250090305
- J. C. Murphy, D. A. Kaden, J. Warren and A. Sivak, “Power Frequency Electric and Magnetic Fields: A Review of Genetic Toxicology,” Mutation Research, Vol. 296, No. 3, 1993, pp. 221-240.
- D. W. Fairbairn and K. L. O'Neill, “The Effects of Electromagnetic Fields Exposure on the Formation of DNA Sinlge Strand Breaks in Human Cells,” Cellular and Molecular Biology, Vol. 40, No. 4, 1994, pp. 561-567.
- I. Nordenson, K. M. Hansson, M. Sandstrom and M. O. Mattson, “Effect of Low-Frequency Magnetic Fields on the Chromosomal Level in Human Amniotic Cells,” In: B. Norden and C. Ramel, Eds., Interaction Mechanisms of Low-Level Electromagnetic Fields in Living Systems—Resonant Phenomena, Oxford University Press, Oxford, 1992, pp. 240-250.
- M. Simko, R. Kriehuber and S. Lange, “Micronucleus Formation in Human Amnion Cells after Exposure to 50 Hz Magnetic fields Applied Horizontally and Vertically,” Mutation Research, Vol. 418, No. 2-3, 1998, pp. 101-111.
- G. R. Ding, K. Wake, M. Taki and J. Miyakoshi, “Increase in Hypoxanthine-Guanine Phosphoribosyl Transferase Gene Mutations by Exposure to Electric Field,” Life Sciences, Vol. 68, 2001, pp. 1041-1046.
 doi:10.1016/S0024-3205(00)01007-9
doi:10.1016/S0024-3205(00)01007-9 - F. I. Wolf, A. Torsello, B. Tedesco, S. Fasanella, A. Boninsegna, M. D’Ascenzo, C. Grassi, G. B. Azzena and A. Cittadini, “50-Hz Extremely Low Frequency Electromagnetic Fields Enhance Cell Proliferation and DNA Damage: Possible Involvement of a Redox Mechanism,” Biochimica et Biophysica Acta, Vol. 1743, No. 1-2, 2005, pp. 120-129. doi:10.1016/j.bbamcr.2004.09.005
- G. R. Verheyen, G. Pauwels, L. Verschaeve and G. Schoeters, “Effect of Coexposure to 50 Hz Magnetic Fields and an Aneugen on Human Lymphocytes, Determined by the Cytokinesis Block Micronucleus assay,” Bioelectromagnetics, Vol. 24, No. 3, 2003, pp. 160-164. doi:10.1002/bem.10100
- R. Pasquini, M. Villarini, G. S. Scassellati, C. Fatigoni and M. Moretti, “Micronucleus Induction in Cells Co-Exposed in Vitro to 50 Hz Magnetic Field and Benzene, 1,4-Benzenediol (Hydroquinone) or 1,2,4-Benzenetriol,” Toxicology in Vitro, Vol. 17, No. 5-6, 2003, pp. 581-586. doi:10.1016/S0887-2333(03)00137-1
- S. Ivancsits, A. Pilger, E. Diem, A. Schaffer and H. W. Rüdiger, “Vanadate Induces DNA Strand Breaks in Cultured Human Fibroblasts at Doses Relevant to Occupational Exposure Mutation Research, Vol. 519, No. 1-2, 2002, pp. 25-35.
- M. C. Pirozzoli, C. Marino, G. A. Lovisolo, C. Laconi, L. Mosiello and A. Negroni, “Effects of 50 Hz Electromagnetic Field Exposure on Apoptosis and Differentiation in a Neuroblastoma Cell Line,” Bioelectromagnetics, Vol. 24, No. 7, 2003, pp. 510-516. doi:10.1002/bem.10130
- A. A. Marino, O. V. Kolomytkin and C. Frilot, “Extracellular Currents Alter Gap Junction Intercellular Communication in Synovial Fibroblasts,” Bioelectromagnetics, Vol. 24, No. 3, 2003, pp. 199-205. doi:10.1002/bem.10085
- R. R. Raylman, A. C. Clavo and R. L. Wahl, “Exposure to Strong Static Magnetic Field Slows the Growth of Human Cancer Cells in Vitro,” Bioelectromagnetics, Vol. 17, 1996, pp. 358-363. doi:10.1002/(SICI)1521-186X(1996)17:5<358::AID-BEM2>3.0.CO;2-2
- S. Pacini, G. B. Vannelli, T. Barni, M. Ruggiero, I. Sardi, P. Pacini and M. Gulisano, “Effect of 0.2 T Static Magnetic Field on Human Neurons: Remodeling and Inhibition of Signal Transduction without Genome Instability,” Neuroscience Letters, Vol. 267, No. 3, 1999, pp. 185-188. doi:10.1016/S0304-3940(99)00362-6
- R. Ohata, N. Tomita and Y. Ikada, “Effect of a Static Magnetic Field on Ion Transport in a Cellulose Membrane,” Journal of Colloid and Interface Science, Vol. 270, No. 2, 2004, pp. 413-416. doi:10.1016/j.jcis.2003.09.035
- A. Hammiche, M. J. German, R. Hewitt, H. M. Pollock and F. L. Martin, “Monitoring Cell Cycle Distributions in MCF-7 Cells Using Near-Field Photothermal Microspectroscopy,” Biophysical Journal, Vol. 88, 2005, pp. 3699- 3706. doi:10.1529/biophysj.104.053926
- H. Y. N. Holman, R. Goth-Goldstein, E. A. Blakely, K. Bjornstad, M. C. Martin and W. R. McKinney, “Individual Human Cell Responses to Low Doses of Chemicals Studied by Synchrotron Infrared Spectromicroscopy,” In: A. Mahadevan-Jansen and G. J. Puppels Ed., Biomedical Spectroscopy: Vibrational Spectroscopy and Other Novel Techniques, Proceedings of SPIE, Vol. 3918, 2000, pp. 57-63. doi:10.1117/12.384959
- K. L. Chan and S. G. Kazarian, “New Opportunities in Micro-and Macro-Attenuated Total Reflection Infrared Spectroscopic Imaging: Spatial Resolution and Sampling Versatility,” Applied Spectroscopy, Vol. 57, No. 4, 2003, pp. 381-389. doi:10.1366/00037020360625907
- J. Zhou, Z. Wang, S. Sun, M. Liu and H. Zhang, “A Rapid Method for Detecting Conformational Changes during Differentiation and Apoptosis of HL60 Cells by FourierTransform Infrared Spectroscopy,” Biotechnology and Applied Biochemistry, Vol. 33, No. 2, 2001, pp. 127-132. doi:10.1042/BA20000074
- K. Z. Liu, L. Jia, S. M. Kelsey, A. C. Newland and H. H. Mantsch, “Quantitative Determination of Apoptosis on Leukemia Cells by Infrared Spectroscopy,” Apoptosis, Vol. 6, No. 4, 2001, pp. 269-278. doi:10.1023/A:1011383408381
- F. Gasparri and M. Muzio, “Monitoring of Apoptosis of HL60 Cells by Fourier-Transform Infrared Spectroscopy,” Biochemical Journal, Vol. 369, 2003, pp. 239-248. doi:10.1042/BJ20021021
- B. M. Smith, and S. Franzen, “Single-Pass Attenuated Total Reflection Fourier Transform Infrared Spectroscopy for the Analysis of Proteins in H2O Solution,” Analytical Chemistry, Vol. 74, No. 16, 2002, pp. 4076-4080. doi:10.1021/ac020103v
- B. Rigas, S. Morgello, I. S. Goldman and P. T. Wong, “Human Colorectal Cancers Display Abnormal FourierTransform Infrared Spectra,” Proceedings of the National Academy of Sciences of the United States of America, New York, Vol. 87, No. 20, 1990, pp. 8140-8144. doi:10.1073/pnas.87.20.8140
- F. S. Parker, “Applications of Infrared Spectroscopy in Biochemistry, Biology, and Medicine,” Plenum Press, New York, 1971.
- C. L. McDowell and E. T. Papoutsakis, “Decreasing Extracellular pH Increases CD13 Receptor Surface Content and Alters the Metabolism of HL60 Cells Cultured in Stirred Tank Bioreactors,” Biotechnology Progress, Vol. 14, No. 4, 1998, pp. 567-572. doi:10.1021/bp980050w
- C. L. McDowell and E. T. Papoutsakis, “Serum Increases the CD13 Receptor Expression, Reduces the Transduction of Fluid-Mechanical Forces, and Alters the Metabolism of HL60 Cells Cultured in Agitated Bioreactors,” Biotechnology and Bioengineering, Vol. 60, No. 2, 1998, pp. 259-268. doi:10.1002/(SICI)1097-0290(19981020)60:2<259::AID-BIT13>3.0.CO;2-H
- W. K. Surewicz and H. H. Mantsch, “New Insight into Protein Secondary Structure from Resolution-Enhanced Infrared Spectra,” Biochimica et Biophysica Acta, Vol. 952, No. 2, 1988, pp. 115-130. doi:10.1016/0167-4838(88)90107-0
- R. Bauer, R. Carrotta, C. Rischel and L. Ogendal, “Characterization and Isolation of Intermediates in β-Lactoglobulin Heat Aggregation at High pH,” Biophysical Journal, Vol. 79, No. 2, 2000, pp. 1030-1038. doi:10.1016/S0006-3495(00)76357-0
- H. Fabian, D. Chapman and H. H. Mantsch, “In Infrared Spectroscopy of Biomolecules,” Wiley-Liss, New York, 1996.
- M. Jackson and H. H. Mantsch, “Biomedical Infrared Spectroscopy,” In: H. H. Mantsch and D. Chapman, Eds., Infrared Spectroscopy of Biomolecules, Wiley-Liss, New York, 1996, pp. 311-340.
- R. Mittler, In: R. A. Lockshin, Z. Zakeri and J. L. Tilly, Eds., When Cells Die: A Comprehensive Evaluation of Apoptosis and Programmed Cell Death, Wiley-Liss, New York, 1998, pp. 147-174.
- R. Birge, E. Fajardo and B. Hempstead, In: R. A. Lockshin, Z. Zakeri, J. L. Tilly, Eds., When Cells Die: A Comprehensive Evaluation of Apoptosis and Programmed Cell Death, Wiley-Liss, New York, 1998, pp. 347-384.
- N. Jamin, P. Dumas, J. Moncuit, W. H. Fridman, J. L. Teillaud, G. L. Carr and G. P. Williams, “Highly Resolved Chemical Imaging of Living Cells by Using Synchrotron Infrared Microspectrometry,” Proceedings of the National Academy of Sciences of the United States of America 95, Vol. 95, 1998, pp. 4837-4840.
- H. Y. N. Holman, M. C. Martin, E. A. Blakely, K. Bjornstad and W. R. McKinney, “IR Spectroscopic Characteristics of Cell Cycle and Cell Death Probed by Synchrotron Radiation Based Fourier Transform IR Spectromicroscopy,” Biopolymers, Vol. 57, No. 6, 2000, pp. 329-333. doi:10.1002/1097-0282(2000)57:6<329::AID-BIP20>3.0.CO;2-2