Applied Mathematics
Vol.07 No.14(2016), Article ID:70021,13 pages
10.4236/am.2016.714132
Mathematical Study of Dengue Disease Transmission in Multi-Patch Environment
Ganga Ram Phaijoo, Dil Bahadur Gurung
Department of Natural Sciences (Mathematics), School of Science, Kathmandu University, Dhulikhel, Nepal
Copyright © 2016 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 20 June 2016; accepted 21 August 2016; published 24 August 2016
ABSTRACT
Dengue disease is the most common vector borne infectious disease transmitted to humans by infected adult female Aedes mosquitoes. Over the past several years the disease has been increasing remarkably and it has become a major public health concern. Dengue viruses have increased their geographic range into new human population due to travel of humans from one place to the other. In the present paper, we have proposed a multi patch SIR-SI model to study the host-vector dynamics of dengue disease in different patches including the travel of human population among the patches. We have considered different disease prevalences in different patches and different travel rates of humans. The dimensionless number, basic reproduction number R0 which shows that the disease dies out if R0 < 1 and the disease takes hold if R0 ≥ 1, is calculated. Local and global stability of the disease free equilibrium are analyzed. Simulations are observed considering the two patches only. The results show that controlling the travel of infectious hosts from high disease dominant patch to low disease dominant patch can help in controlling the disease in low disease dominant patch while high disease dominant becomes even more disease dominant. The understanding of the effect of travel of humans on the spatial spread of the disease among the patches can be helpful in improving disease control and prevention measures. In the present study, a patch may represent a city, a village or some biological habitat.
Keywords:
Dengue, Patch, Basic Reproduction Number, Equilibrium Point, Stability

1. Introduction
Dengue disease is regarded as a serious infectious disease. The four serotypes of viruses DEN 1 to DEN 4 are responsible for the disease. It is one of the re-emerging diseases in tropical and subtropical countries. A person infected by one of the four serotypes of dengue viruses will never be infected again by the same serotype, but the person loses immunity to other serotype of viruses and becomes more susceptible in developing dengue hemorrhagic fever [1] . The prevalence of the disease has been increasing dramatically and the disease has become a major public health problem in recent years. According to World Health Organization, dengue has shown 30 fold increase globally over five decades. About 50 - 100 million new infections are estimated to occur annually in more than 100 endemic countries. Almost fifty percent of the world’s population lives in the countries where dengue is endemic [2] .
There have been many mathematical studies to understand the dynamics of infectious diseases. Mathematical models can help in providing guides and suggestions for the control of the disease to the concerned authorities. Kermack and McKendrick introduced an SIR model to study the transmission of infectious diseases [3] which became very popular in the mathematical study of epidemic diseases. Esteva and Vargas proposed an SIR-SI model to study the transmission dynamics of dengue disease considering constant [4] and variable [5] host populations. Since then, different mathematical models have been proposed to study dengue disease transmission. Authors in [6] [7] studied the impact of awareness in the transmission of dengue disease. Pinho et al. [8] used mathematical model for dengue disease transmission with the aim of analyzing and comparing two dengue epidemics that occurred in Brazil. Pongsumpun [9] studied the incubation period of dengue viruses using SEIR model. Edy and Supriatna proposed a two dimensional epidemic model to study the transmission of dengue disease restricting the dynamics for two dimensions for the constant host and vector populations [10] .
Emerging and re-emerging diseases like dengue disease spread very quickly due to the travel of infective human population from one region to the other. They spread the disease in new regions. Different spatial models have been developed to study infectious diseases. Arino and Driessche [11] [12] studied the disease spread in meta-populations and they developed multicity model to study the infectious diseases in different cities. Wang and Mulone [13] ; and Wang and Zhao [14] proposed epidemic models with population dispersal to describe the dynamics of disease spread between n patches and two patches. Hsieh et al. proposed a multi-patch epidemic model to study the impact travel between patches for the spatial spread of influenza [15] .
Lee and Castillo-Chavez [16] formulated the two patch dengue transmission model to explore the role of residence times in dengue transmission dynamics and optimal control strategies assuming that only the human budgets their residence time across the patches. In the present work, we have discussed the multi-patch SIR-SI model to study the transmission dynamics of dengue disease among n-patches. We have investigated the impact of travel rates of humans in the transmission dynamics and control of dengue disease. We have assumed different travel rates and different disease prevalences in different patches.
2. Model Formulation
For the formulation of the model, we divide human population in three classes, susceptible, infective and recovered. Let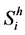 ,
, 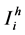 ,
, 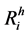 respectively denote the number of susceptible humans, infective humans and recovered humans in patch i. Also, we divide mosquito population in two compartments only, susceptible and infective mosquitoes. Let
respectively denote the number of susceptible humans, infective humans and recovered humans in patch i. Also, we divide mosquito population in two compartments only, susceptible and infective mosquitoes. Let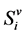 ,
, 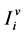 respectively denote the number of susceptible mosquitoes and infective mosquitoes in patch
respectively denote the number of susceptible mosquitoes and infective mosquitoes in patch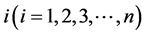 .
.
The SIR-SI Model for 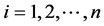 for dengue disease transmission shown in Figure 1, whose parameters are discussed in Table 1, is described by the following system of differential equations
for dengue disease transmission shown in Figure 1, whose parameters are discussed in Table 1, is described by the following system of differential equations
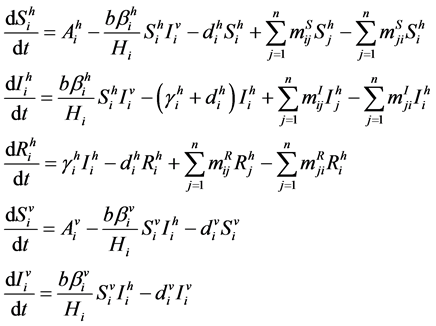 (2.1)
(2.1)

Figure 1. Flow chart of the model.

Table 1. Parameters used in the model.
where,
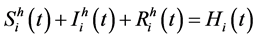 (Total host population in patch i in time t)
(Total host population in patch i in time t)
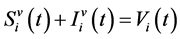 (Total vector population in patch i in time t)
(Total vector population in patch i in time t)
The total host and vector population sizes in all n-patches in time t is
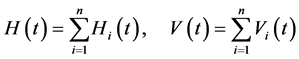
Theorem 1. The system of Equations (2.1) has a unique disease free equilibrium point.
Proof: A disease free equilibrium (DFE) for the system of Equations (2.1) is a steady state solution of the system where 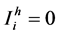 and
and 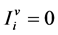 for
for .
.
In disease free situation,

In matrix form,
 (2.2)
(2.2)
where,

Here,  has all off-diagonal entries negative and every column has positive sum. So,
has all off-diagonal entries negative and every column has positive sum. So,  is a non-singular M-matrix. Since all the off diagonal elements are non-zero,
is a non-singular M-matrix. Since all the off diagonal elements are non-zero,  is irreducible [17] . Hence,
is irreducible [17] . Hence,  has a positive inverse and the system of Equations (2.2) has a unique solution. So,
has a positive inverse and the system of Equations (2.2) has a unique solution. So,  is the solution of the system, i.e.,
is the solution of the system, i.e., for
for . Hence, in disease free situation,
. Hence, in disease free situation,  ,
,  ,
,  for all
for all . Also,
. Also,  ,
, .
.
Now, we show that the disease free equilibrium is unique. From the system of Equations (2.1), in disease free situation:
For the host populations only:

i.e.,
 (2.3)
(2.3)
where,
 ,
, 

For vector populations only:

i.e.,
 (2.4)
(2.4)
where,

Here, the matrix C has positive column sums and each non-diagonal element is negative. So, the matrix C is an irreducible and non-singular M-matrix. Again, since C is an irreducible non-singular M-matrix, C must have positive inverse, i.e.,  [17] . Hence, there is a unique solution
[17] . Hence, there is a unique solution .
.
Also, the matrix D is a diagonal matrix with positive diagonal elements. So, there exists  with positive diagonal elements. Hence,
with positive diagonal elements. Hence,  is unique solution of
is unique solution of . The results show that there always exists a unique disease free equilibrium point.
. The results show that there always exists a unique disease free equilibrium point.
3. Basic Reproduction Number
Basic reproduction number  is defined as the expected number of secondary cases produced by a typical infective individual introduced into a completely susceptible population.
is defined as the expected number of secondary cases produced by a typical infective individual introduced into a completely susceptible population.
We use next generation matrix method [18] [19] to find the basic reproduction number. For, we order the infected variables by . Then,
. Then,


where,

Here, the matrix  has column sums,
has column sums,  and all off diagonal elements are negative. So, the matrix
and all off diagonal elements are negative. So, the matrix  is an irreducible non-negative M-matrix. Hence,
is an irreducible non-negative M-matrix. Hence,  exists and is positive, i.e.,
exists and is positive, i.e., .
.
Also,  is a diagonal matrix with positive entries. So, nonnegative
is a diagonal matrix with positive entries. So, nonnegative  exists. The basic reproduction number
exists. The basic reproduction number  for the system (2.1) is the spectral radius of
for the system (2.1) is the spectral radius of .
.
In fact,

Theorem 2. If R0 < 1, then the disease free equilibrium is locally asymptotically stable and unstable if R0 > 1.
Proof: Let  and
and  be the matrices of partial derivatives evaluated at the disease free equilibrium. The Jacobian matrix for the linearization of the system about the disease free equilibrium is obtained as the block structure
be the matrices of partial derivatives evaluated at the disease free equilibrium. The Jacobian matrix for the linearization of the system about the disease free equilibrium is obtained as the block structure

Matrix J is triangular. So, the eigenvalues of J are those of the partition matrices  and
and . Also,
. Also,

Matrices C and D (matrices defined in Theorem 1) are non-singular M matrices. So, spectral abscissa,  [17] and eigenvalues of the matrix
[17] and eigenvalues of the matrix  have negative real parts.
have negative real parts.
Hence, the matrix J will have eigenvalues all with negative real parts if the matrix  has all eigenvalues with negative real parts. Also, F is non-negative matrix and V is non-singular M-matrix. So, eigenvalues of
has all eigenvalues with negative real parts. Also, F is non-negative matrix and V is non-singular M-matrix. So, eigenvalues of  will have negative real parts if and only if
will have negative real parts if and only if  [19] . i.e., disease free equilibrium is locally asymptotically stable if and only if the basic reproduction number
[19] . i.e., disease free equilibrium is locally asymptotically stable if and only if the basic reproduction number .
.
If , then
, then . It shows that at least one eigenvalue lies in right half plane. So, the disease free equilibrium is unstable if
. It shows that at least one eigenvalue lies in right half plane. So, the disease free equilibrium is unstable if .
.
Theorem 3. If R0 < 1, then the disease free equilibrium is globally asymptotically stable and unstable if R0 > 1.
Proof: Since,  and
and , we have from the system of Equations (2.1),
, we have from the system of Equations (2.1),
 (3.1)
(3.1)
Consider the linear system
 (3.2)
(3.2)
The system of Equations (3.2) can be written as
 (3.3)
(3.3)
where,  Here, F is a non-negative matrix and V is a non-negative M- matrix. So,
Here, F is a non-negative matrix and V is a non-negative M- matrix. So,

i.e., Eigenvalues of  lie on left half plane if
lie on left half plane if . Hence, each positive solution of (3.3) satisfies
. Hence, each positive solution of (3.3) satisfies
 (3.4)
(3.4)
i.e.,  for all
for all 
Since all the variables in the system of Equations (2.1) are non-negative, the use of Comparison theorem [20] [21] leads to
 for all
for all  (3.5)
(3.5)
From the system of Equations (2.1), we have
 (3.6)
(3.6)
i.e.,  (In matrix form)
(In matrix form)
Here,  (matrix defined in Theorem 1) is non-singular M-matrix. So, all eigenvalues of
(matrix defined in Theorem 1) is non-singular M-matrix. So, all eigenvalues of  lie in the left half plane. Hence,
lie in the left half plane. Hence,
 .
.
Again, as ,
,
 (3.7)
(3.7)
In matrix form
 (3.8)
(3.8)
 (3.9)
(3.9)
Here, matrices C and D are non-singular M-matrices, all their eigenvalues lie in left half plane. Therefore, if  and
and  be the homogeneous solutions of Equation (3.8) and Equation (3.9), then
be the homogeneous solutions of Equation (3.8) and Equation (3.9), then
 and
and 
Matrix C is an irreducible, non-singular M-matrix. So, the matrix C has positive inverse.  is a particular solution and
is a particular solution and  is the general solution of Equation (3.8). Also, Matrix D is diagonal matrix with positive diagonal elements. So, D has an inverse with positive diagonal elements. Hence,
is the general solution of Equation (3.8). Also, Matrix D is diagonal matrix with positive diagonal elements. So, D has an inverse with positive diagonal elements. Hence,  is a particular solution and
is a particular solution and  is the general solution of Equation (3.9). And,
is the general solution of Equation (3.9). And,

for all 
Thus, as , we obtain the equilibrium point
, we obtain the equilibrium point . Hence, the disease free equilibrium is globally asymptotically stable if
. Hence, the disease free equilibrium is globally asymptotically stable if . If
. If  [19] , Theorem 2 admits that the disease free equilibrium point is unstable.
[19] , Theorem 2 admits that the disease free equilibrium point is unstable.
4. For n = 2 (Considering Two Patches Only)
We have

and ,
, 
Basic Reproduction Number

where,

 (Basic reproduction number, Patch 1)
(Basic reproduction number, Patch 1)
 (Basic reproduction number, Patch 2)
(Basic reproduction number, Patch 2)
5. Numerical Results and Discussions
We considered the case of two patches and computed basic reproduction number  for the numerical results. The parameter values chosen for the simulation are:
for the numerical results. The parameter values chosen for the simulation are: ,
,  [22] ,
[22] ,  ,
,  ,
, [8] ,
[8] ,  ,
,  ,
,  [8] ,
[8] ,  ,
,  [9] ,
[9] ,  [22] ,
[22] ,  [9] . With
[9] . With  and
and . Thus, patch 1 is a high disease dominant patch and patch 2 is a low disease dominant patch.
. Thus, patch 1 is a high disease dominant patch and patch 2 is a low disease dominant patch.
Figure 2 shows the dynamics of susceptible hosts of patch 1 and patch 2. Patch 1 is a high disease dominant patch, so the susceptible host population in the patch decreases most rapidly over the time. Patch 2 is a low disease dominant patch. So, the number of susceptible hosts in patch 2 increases initially due to the travel of susceptible hosts from patch 1. Afterwards, due to the interaction of susceptible hosts with infectious mosquitoes and due to the natural death of some humans, the susceptible host population starts decreasing.
When the susceptible hosts come in contact with infectious mosquitoes, hosts get infected. So, the population size of infected hosts increases (Figure 3). Eventually, the infected host population decreases to zero due to their recovery from the disease and due to the natural death of some humans.
Changes in basic reproduction number  with the changes in travel rates are illustrated in Figure 4. It is observed that basic reproduction number
with the changes in travel rates are illustrated in Figure 4. It is observed that basic reproduction number  decreases with the increasing values of
decreases with the increasing values of . Also, the number increases when the values of travel rate
. Also, the number increases when the values of travel rate  are increased. The figure shows that, the burden of disease reduces when the travel rates of hosts from high disease dominant patch to low disease dominant patch are high. The burden of disease increases when the travel rates of hosts from low disease dominant patch to the high disease
are increased. The figure shows that, the burden of disease reduces when the travel rates of hosts from high disease dominant patch to low disease dominant patch are high. The burden of disease increases when the travel rates of hosts from low disease dominant patch to the high disease

Figure 2. Dynamics of susceptible host population.

Figure 3. Dynamics of infected host population.
dominant patch are high. Thus, we should increase the travel rates of hosts from high disease dominant patch to the low disease dominant patch to bring the disease under control.
Basic reproduction number  of two patches is determined by the basic reproduction numbers,
of two patches is determined by the basic reproduction numbers,  of patch 1 and
of patch 1 and  of patch 2. Figure 5 shows that
of patch 2. Figure 5 shows that  increases together with
increases together with  and
and . If basic reproduction numbers of the both patches are high, then the basic reproduction number
. If basic reproduction numbers of the both patches are high, then the basic reproduction number  gets higher. Thus, if any one of the two patches is more disease dominant and there is mobility between the two patches, then this can cause the whole system to be more endemic.
gets higher. Thus, if any one of the two patches is more disease dominant and there is mobility between the two patches, then this can cause the whole system to be more endemic.
6. Discussion of Travel Restrictions
In this section, the dynamics of the host population is observed with the restriction of the travel of symptomatic hosts from one patch to the other patch.
Restricting the travel of symptomatic hosts from low disease dominant patch to high disease dominant patch

Figure 4. Basic reproduction number against  and
and .
.

Figure 5. Basic reproduction number  against
against  and
and .
.
( and keeping other parameters constant), Figure 6 shows that the burden of disease can be reduced in patch 1 but patch 2 becomes even more disease dominant (Figure 7).
and keeping other parameters constant), Figure 6 shows that the burden of disease can be reduced in patch 1 but patch 2 becomes even more disease dominant (Figure 7).
Similarly, when  and all other parameters are same i.e., on restricting the travel of symptomatic travelers from high disease dominant patch to low disease dominant patch, we find that basic reproduction number of patch 1 increases (Figure 8) and basic reproduction number of patch 2 decreases (Figure 9). Thus, the disease in low disease dominant patch can be controlled by restricting the travel of symptomatic hosts from high disease dominant patch to low disease dominant patch.
and all other parameters are same i.e., on restricting the travel of symptomatic travelers from high disease dominant patch to low disease dominant patch, we find that basic reproduction number of patch 1 increases (Figure 8) and basic reproduction number of patch 2 decreases (Figure 9). Thus, the disease in low disease dominant patch can be controlled by restricting the travel of symptomatic hosts from high disease dominant patch to low disease dominant patch.
Dynamics of infected host populations are observed in Figure 10 and Figure 11 with travel restrictions. When infected hosts of patch 1 are restricted to travel (Figure 10) more hosts in patch 1 (very few hosts in patch 2) are observed infected of the disease when compared with the case that infected hosts of patch 2 are restricted (Figure 11) to travel. The graphical results (Figure 10, Figure 11) suggest that the disease spread in patch 2 can be brought under control by restricting the travel of infected hosts from patch 1 to patch 2.

Figure 6. Basic reproduction number of patch 1 against  with
with .
.

Figure 7. Basic reproduction number of patch 2 against  with
with .
.

Figure 8. Basic reproduction number of patch 1 against  with
with .
.

Figure 9. Basic reproduction number of patch 2 against  with
with .
.
7. Conclusions
In the present work, we have studied the effect of travel of humans on the transmission dynamics of dengue disease. We discussed the disease transmission dynamics between n-patches by subdividing vector population in susceptible and infectious class and host population in susceptible, infectious and recovered class.
We defined the multi-patch basic reproduction number  by taking each patch together. Basic reproduction number
by taking each patch together. Basic reproduction number  of patch 1 and
of patch 1 and  of patch 2 are calculated. The results show that the disease dies out if
of patch 2 are calculated. The results show that the disease dies out if  and invades the population if
and invades the population if . Theorem 2 and Theorem 3 show that the disease free equilibrium is locally and globally asymptotically stable if
. Theorem 2 and Theorem 3 show that the disease free equilibrium is locally and globally asymptotically stable if  and unstable if
and unstable if .
.
Travel of human from one place to another place affects the whole dynamics of the dengue disease transmission. We have shown that traveling of infected human changes the less disease dominant patch to high disease

Figure 10. Dynamics of infected host population with .
.

Figure 11. Dynamics of infected host population with .
.
dominant patch. Also, restricting the travel of infected hosts helps in controlling the disease. Basic reproduction number is seen higher when there is higher travel rate from low disease dominant patch to the high disease dominant patch. The basic reproduction number is seen lowered when there is higher travel rate from high dominant disease patch to the low disease dominant patch. Thus, we can control the disease in low disease dominant patch by restricting the travel of infected hosts from high disease dominant patch.
Cite this paper
Ganga Ram Phaijoo,Dil Bahadur Gurung, (2016) Mathematical Study of Dengue Disease Transmission in Multi-Patch Environment. Applied Mathematics,07,1521-1533. doi: 10.4236/am.2016.714132
References
- 1. Gubler, D. (1998) Dengue and Dengue Hemorrhagic Fever. Clinical Microbiology Reviews, 3, 480-496.
- 2. World Health Organization (2012) Global Strategy for Dengue Prevention and Control 2012-2020, World Health Organization, Geneva.
- 3. Kermack, W.O. and McKendrick, A.G. (1927) A Contribution to the Mathematical Theory of Epidemics. Proceedings of the Royal Society of London, 115, 700-721.
http://dx.doi.org/10.1098/rspa.1927.0118 - 4. Esteva, L. and Vargas, C. (1998) Analysis of a Dengue Disease Transmission Model. Mathematical Biosciences, 150, 131-151.
http://dx.doi.org/10.1016/S0025-5564(98)10003-2 - 5. Esteva, L. and Vargas, C. (1999) A Model for Dengue Disease with Variable Human Population. Journal of Mathematical Biology, 38, 220-240.
http://dx.doi.org/10.1007/s002850050147 - 6. Gakkhar, S. and Chavda, N.C. (2013) Impact of Awareness on the Spread of Dengue Infection in Human Population. Applied Mathematics, 4, 142-147.
http://dx.doi.org/10.4236/am.2013.48A020 - 7. Phaijoo, G.R. and Gurung, D.B. (2015) Mathematical Study of Dengue Disease with and without Awareness in Host Population. International Journal of Advanced Engineering Research and Applications (IJAERA), 1, 239-245.
- 8. Pinho, S.T.R., Ferreira, C.P., Esteva, L., Barreto, F.R., Morato e Silva, V.C. and Teixeira, M.G.L. (2010) Modelling the Dynamics of Dengue Real Epidemics. Philosophical Transactions of the Royal Society A, 368, 5679-5693.
http://dx.doi.org/10.1098/rsta.2010.0278 - 9. Pongsumpun, P. (2008) Mathematical Model of Dengue Disease with the Incubation Period of Virus. World Academy of Science, Engineering and Technology, 44, 328-332.
- 10. Soewono, E. and Supriatna, A.K. (2001) A Two-Dimensional Model for the Transmission of Dengue Fever Disease. Bulletin of the Malaysian Mathematical Sciences Society, 24, 49-57.
- 11. Arino, J. and Van den Driessche, P. (2006) Disease Spread in Metapopulations. Field Institute Communications, 48, 1-12.
- 12. Arino, J. and van den Driessche, P. (2003) A Multicity Epidemic Model. Mathematical Population Studies, 10, 175-193.
http://dx.doi.org/10.1080/08898480306720 - 13. Wang, W. and Mulone, G. (2003) Threshold of Disease Transmission in a Patch Environment. Journal of Mathematical Analysis and Applications, 285, 321-335.
http://dx.doi.org/10.1016/S0022-247X(03)00428-1 - 14. Wang, W. and Zhao, X.Q. (2007) An Epidemic Model in a Patchy Environment. Mathematical Biosciences, 112, 97-112.
- 15. Hsieh, Y.H., van den Driessche, P. and Wang, L. (2007) Impact of Travel between Patches for Spatial Spread of Disease. Bulletin of Mathematical Biology, 69, 1355-1375.
http://dx.doi.org/10.1007/s11538-006-9169-6 - 16. Lee, S. and Castillo-Chavez, C. (2015) The Role of Residence Times in Two-Patch Dengue Transmission Dynamics and Optimal Strategies. Journal of Theoretical Biology, 374, 152-164.
http://dx.doi.org/10.1016/j.jtbi.2015.03.005 - 17. Berman, A. and Plemmons, R.J. (1979) Non-Negative Matrices in Mathematical Sciences. Academic Press, New York.
- 18. Diekmann, O., Heesterbeek, J.A.P. and Metz, J.A.J. (1990) On the Definition and Computation of the Basic Reproduction Ratio RO in Models for Infectious Diseases in Heterogeneous Populations. Journal of Mathematical Biology, 28, 365-382.
http://dx.doi.org/10.1007/BF00178324 - 19. van den Driessche, P. and Watmough, J. (2007) Reproduction Numbers and Sub-Threshold Endemic Equilibria for Compartmental Models of Disease Transmission. Mathematical Biosciences, 180, 29-48.
http://dx.doi.org/10.1016/S0025-5564(02)00108-6 - 20. Lakshmikantham, V., Leela, S. and Martynyuk, A.A. (1998) Stability Analysis of Non-Linear Systems. Dekker, Florida.
- 21. Smith, H.L. and Waltman, P. (1995) The Theory of Chemostat. Cambridge University Press, New York.
http://dx.doi.org/10.1017/CBO9780511530043 - 22. Central Bureau of Statistics (2014) Statistical Pocket Book of Nepal. Government of Nepal, Ramshahpath, Kathmandu.