Advances in Microbiology
Vol.4 No.12(2014), Article ID:49774,8 pages
DOI:10.4236/aim.2014.412087
Sweet Potato Starch as a Carbon Source for Growth and Glucoamylase Production from Aspergillus niger
Alfred O. Ubalua
Tissue Culture Unit, Biotechnology Research and Development Center, National Root Crops Research Institute (NRCRI), Umuahia, Nigeria
Email: alfreduba@yahoo.com
Copyright © 2014 by author and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 4 July 2014; revised 1 August 2014; accepted 2 September 2014
A mesophilic strain of Aspergillus niger isolated from cassava effluent samples produced extracellular glucoamylase in submerged culture containing 2% (w/v) soluble or sweet potato starch. On soluble starch medium the maximum glucoamylase activity in the culture filtrate was 9.40 U/mg compared to 8.24 U/mg on sweet potato starch culture filtrate. The mycelial dry weight for both media was 494 and 418 mg respectively. The maximum glucoamylase activity was obtained at a growth temperature of 40˚C and pH 4.5. The implication is that the bioprocess of utilizing sweet potato starch in the culture is attractive due to its relatively cheaper availability in Nigeria, making it even more favorable when economics is considered.
Keywords:Aspergillus niger, Sweet Potato Starch, Submerged Fermentation, Amylolytic Activity, Glucoamylase Prodcution

Aspergillus niger remains one of the most important microorganisms with biotechnological potentials. Since 1919, A. niger has been exploited industrially in the production of citric acid. Its scope of practical importance was further broadened in the last three decades, when the organism was developed as an important transformation host to over-express food enzymes that are well established as technical aids in fruit processing, baking, and in starch and food industries. Amyloglucosidase, pectinase and protease were originally produced by A. niger in surface culture [1] and later in submerged culture [2] . Recently, amyloglucosidase has been exploited for industrial hydrolysis of starch to glucose, maltose, and low molecular weight dextrins. Besides the production of food enzymes, citric acid is also produced almost exclusively by the fermentation of A. niger and A. wentii. Citric acid is the primary acidulant in the food and beverage industries and also in foods such as soft drinks, fruit juices, desserts, jams, jellies, candy and wine [3] .
Medium composition is a prerequisite for successful growth and enzyme production by microorganisms. Medium ingredient such as soluble starch is imported and considerably expensive in sub-Saharan Africa necessitating the search for local alternatives. In Nigeria, agricultural produce like roots and tubers, are abundant; and a valuable percentage are perennially lost due to inadequate storage and processing. The use of starches as raw materials is attractive because of the enormous post-harvest losses associated with our agricultural produce and the ability to obtain high root yields of sweet potato from marginal agricultural soils in our country Nigeria. The possibility that cheap raw materials can increase the yield of amylases has not received much attention. Moreover, the economy and sustainability of many fermentation processes depends to some extent on the cost of carbohydrate raw materials [4] . Complex fermentation media which gave optimum production of amylases have been reported [5] , but not much attempts have been made to develop cheap substrate for optimum production of amylases. The regular use of soluble starch-based fermentation media is not commercially viable for industries in the tropics. For efficient commercial production, a continuous effort is being made to find cheaper substrate sources. Locally available carbon and nitrogenous sources are the decisive factors in the optimum production of enzymes, and these differ very much from substrate to substrate. In this study, the potential of obtaining significant yield of amylase in a mesophilic strain of A. niger isolated from cassava effluent using locally available and cheap sweet potato starch in comparison with the conventional soluble starch was investigated.
2.1. Isolation of Aspergillus niger
Aspergillus niger used in this study was isolated from cassava effluent samples. Samples were collected with clean sealed cellophane bags and analyzed immediately. Each sample was evenly mixed with sterile distilled water in a test tube and serially diluted. A 0.1 ml of the diluent was plated on potato dextrose agar (PDA; Sigma Chemical Co., St. Louis, MO, USA) medium fortified with 0.1 mg/ml of streptomycin, in triplicate. Incubation was at room temperature for 48 h. After 48 h of incubation, all morphologically contrasting colonies were purified by repeated streaking. Purified isolates were also scored for starch hydrolysis as described by Chu [6] . This was done with PDA medium containing 1% soluble starch. Ability to hydrolyse starch was demonstrated by streaking the isolates on the PDA plates and incubation was at room temperature for 48 h before flooding with Grams iodine. In situ enzyme production was observed with the clearing of zone of whitish appearance in the areas surrounding the isolate. Identification was on the basis of the structures bearing spores, stolon, and conidia. The observed structures were compared with the standard descriptions in Barnett and Hunter [7] . The initial selection of the amylase-producing A. niger was based on the formation of clearance zone with iodine solution on the isolation medium containing 1% soluble starch.
2.2. Profile of Growth and Amylase Induction from Aspergillus niger
250 ml Erlenmeyer flasks were used for the amylase production. Each contained 50 ml of the culture medium of the following composition in g/l: Soluble starch (20), peptone (0.5), K2HPO4 (1), KH2PO4 (0.60), MgSO4∙7H2O (0.10), CaCl2∙2H2O (0.10) (MS). The pH of the medium was adjusted to 7.0 with 1 N H2SO4 and/or 1 N NaOH before sterilization at 121˚C for 15 minutes. Spores of 48 h old cultures of A. niger were harvested by washing with 5 ml of sterile distilled water. The concentration was adjusted to 4 × 107 spores/ml with a haemocytometer and a 0.5 ml of the spore suspension was used to inoculate the medium. Fermentation was performed with mild agitation on a rotary shaker at 200 rpm for 72 h and at room temperature. The following variables were monitored daily: protein content, enzyme activity, specific activity, and starch depletion. For comparative purposes, the soluble starch in the amylase producing medium was substituted with sweet potato starch. Filtration was done with Whatman No. 5 filter paper (diameter 15 cm, and aperture size 2.5 µm) dried to constant weight at 60˚C and the filtrates thus obtained were taken as the crude enzyme and used for growth measurements.
2.3. Extraction of Starch Milk
Mature sweet potato roots were peeled and washed with clean water and allowed to dry at room temperature. They were subsequently disintegrated with a grater. The pulps were placed on a muslin cloth, sieved and flushed with clean water until the starch stream ceases. The crude starch milks were kept in a refrigerator overnight to discourage microbial growth and degradation. The supernatants were decanted leaving the precipitated starch.
2.4. Purification of the Starches
Three (3) liters of clean water equivalent to three times the volume of the crude starch was added and stirred until all the starch was suspended in the water. The suspension was undisturbed until sedimentation ceases. The supernatant was decanted leaving the sediments in a plastic bucket. An equivalent volume of fresh clean water was added again to the volume of the starch sediments and stirred until the starch was suspended in the water. The starch slurry was filtered through a muslin cloth sieve of about 125 micron pore size and flushed with water. The filtrates were allowed to sediment for 20 minutes and the supernatants decanted. This procedure was repeated two times yielding a clean starch cake.
2.5. Quality Control of the Starches
A teaspoonful sample was taken from the top of the clean starch cake sediment and dissolved in a 15 ml of water in a test tube. The tube and its content were centrifuged for 20 minutes at 2000 rpm and a small sample was drawn from the boundary between the liquid and the sediment using a pipette. Observation was done under a microscope (magnification ×100). Absence of particles in the sample under observation confirmed its purity.
2.6. Drying and Storage
The starch cakes were spread on a table and allowed to dry. They were pulverized with a milling machine (Thomas Wiley Mill-model Ed-5, Philadelphia, USA, 1982), stored in bottles and preserved in a refrigerator (Haier Thermocool, model HRF-350, 2001).
2.7. Assay of Amylase Production
Amylase activity was determined in 1 ml containing 0.5 ml of 1% (w/v) soluble starch, 0.1 ml of the enzyme solution and 0.4 ml of 0.1 M phosphate buffer (pH 6). After 5 min at 25˚C, reducing sugar was determined by the method of Nelson [8] . One unit of amylase is defined as the amount of enzyme that releases one µmole of reducing sugar (with glucose as standard) per minute under the assay conditions. Specific activity was expressed as U/mg protein.
2.8. Physicochemical Properties of the Enzyme
2.8.1. Effect of pH on Growth and Amylase Production
Buffered amylase medium was used for the determination of the growth measurements. The pH range considered was 3, 4, 5, 6, and 7. The flasks were incubated at room temperature for 7 days.
2.8.2. Effect of Temperature on Growth and Amylase Production
Aspergillus niger culture was incubated at various temperatures (25˚C - 70˚C) for 7 days. Optimum temperature for growth and amylase activity were also determined.
2.8.3. Analysis of Data
All experiments were carried out in triplicate and data subjected to statistical analysis using Gen Stat. Discovery Edition 3, HP1 1ES UK.
3.1. Characterization of the Fungal Isolate
The growth of the isolate on potato dextrose agar was rapid. The organism was slightly brownish with smooth surface on potato dextrose agar (Plate 1) and with contrasting yellow colour on the reverse side of the culture plate (Plate 2). Conidiophores were upright, simple and terminating in a swelling bearing phialides at the apex with many colours. The organism demonstrated starch hydrolyzing ability by showing area of clear zone along the line of streaking after 36 h incubation and flooding with Grams iodine solution (Plate 3). Based on its growth, morphological appearance and microscopic structures the organism was identified as Aspergillus niger.
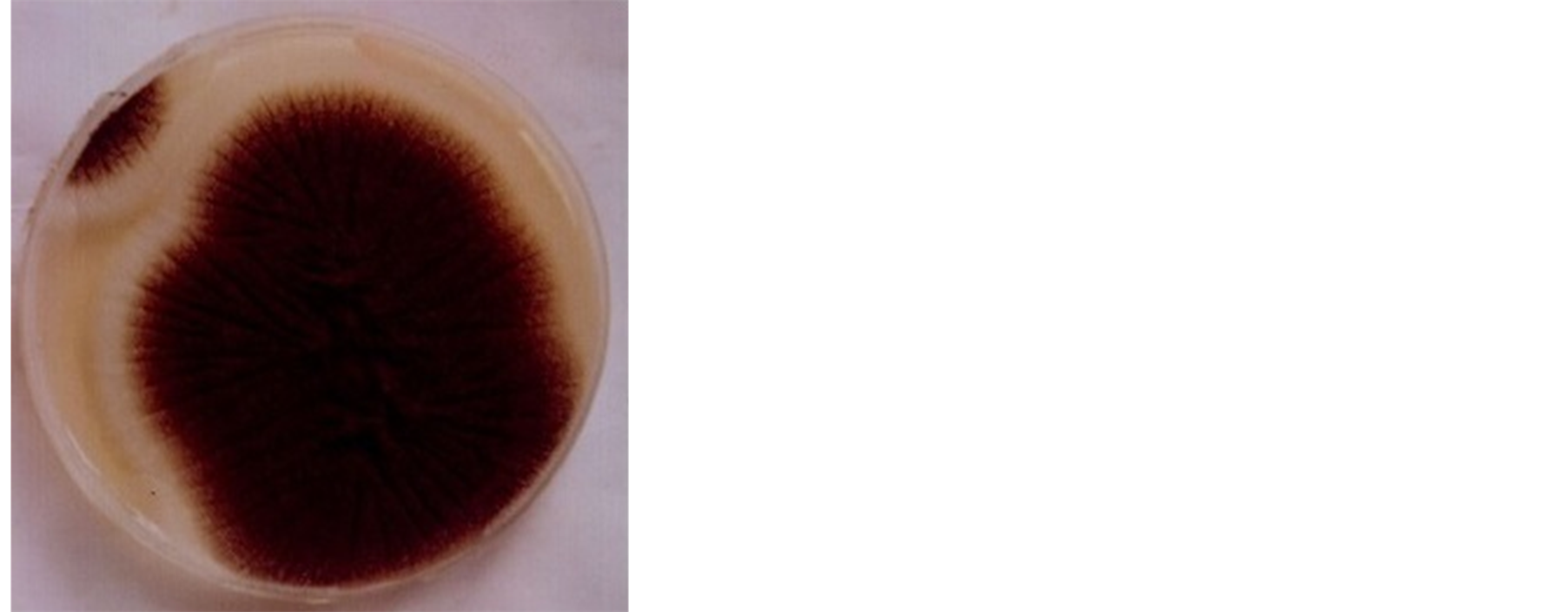
Plate 1. Aspergillus niger.
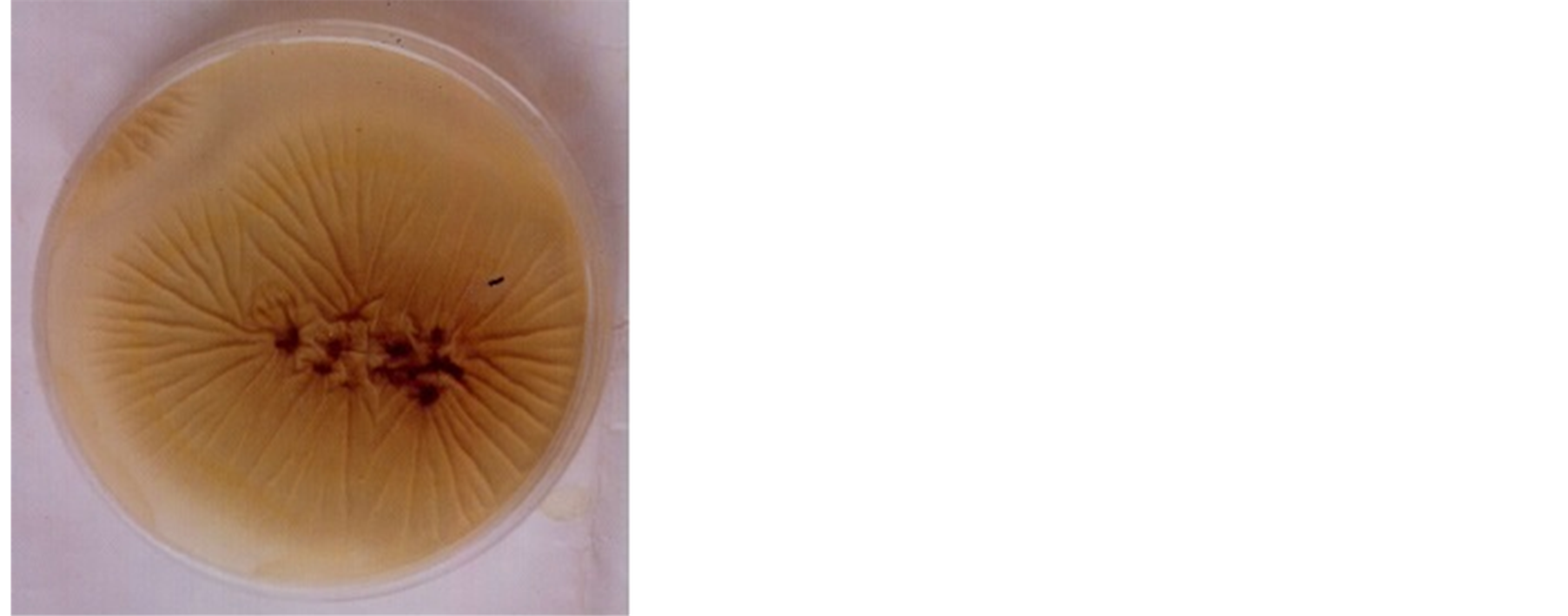
Plate 2. Reverse side of Aspergillus niger.


Plate 3. Aspergillus niger showing whitish area of zone of clearing after incubation and flooding with Grams iodine solution.
3.2. Amylase Production by Aspergillus niger
Growth progression and pattern of A. niger, amylase production is as shown in Figure 1. Complex media are of essence for the optimum production of amylase by A. niger. Induction of extracellular glucoamylase was highest in growth medium containing 2% (w/v) soluble starch, followed closely by sweet potato starch. A maximum amylase activity of 8.24 U/mg was produced on sweet potato growth medium at the end of stationary phase compared to 9.40 U/mg on soluble starch medium (Figure 1). The observed maximum activities coincided with 48 h of incubation (Figure 1) which corresponded to the organisms early stationary phase. Sani et al. [9] and Uguru et al. [10] observed highest amylase activity in Aspergillus sp. grown on starches from cassava peel and yam peel at the late and early stationary phase respectively. At these maximum amylase activity productivity stages, about 58% of the sweet potato starch was utilized in the growth medium compared to 71% of the soluble starch implying that soluble starch in the medium was easily metabolized by the organism. However, the ability of sweet potato starch medium to induce a competitive optimal amylase titre of 8.24 U/mg compared to 9.40 U/mg on soluble starch medium is remarkable and advantageous in the sub-Saharan Africa. Therefore, since sweet potato starch is cheap and readily available compared to soluble starch, it could serve as a possible alternative for the cost-effective production of amylases when used as culture medium ingredient especially in sub-Saharan Africa.
3.3. Substrate Specificity of Amylase Production by Aspergillus niger
To determine the effect of medium composition on the production of amylase, A. niger was grown in mineral salts media devoid of carbon source (sweet potato and soluble starch). Growth of A. niger and amylase production were assessed at the end of the fourth day. Mycelial dry weight produced in the medium without carbon source was 58 mg (50 ml). But when the organism was grown in soluble or sweet potato starch alone, growth increased appreciably to 273 and 249 mg respectively (Figure 2). The corresponding amylase activity was 0.13 and 0.08 U/mg respectively. The implication is that carbon source is indispensable for optimum production of
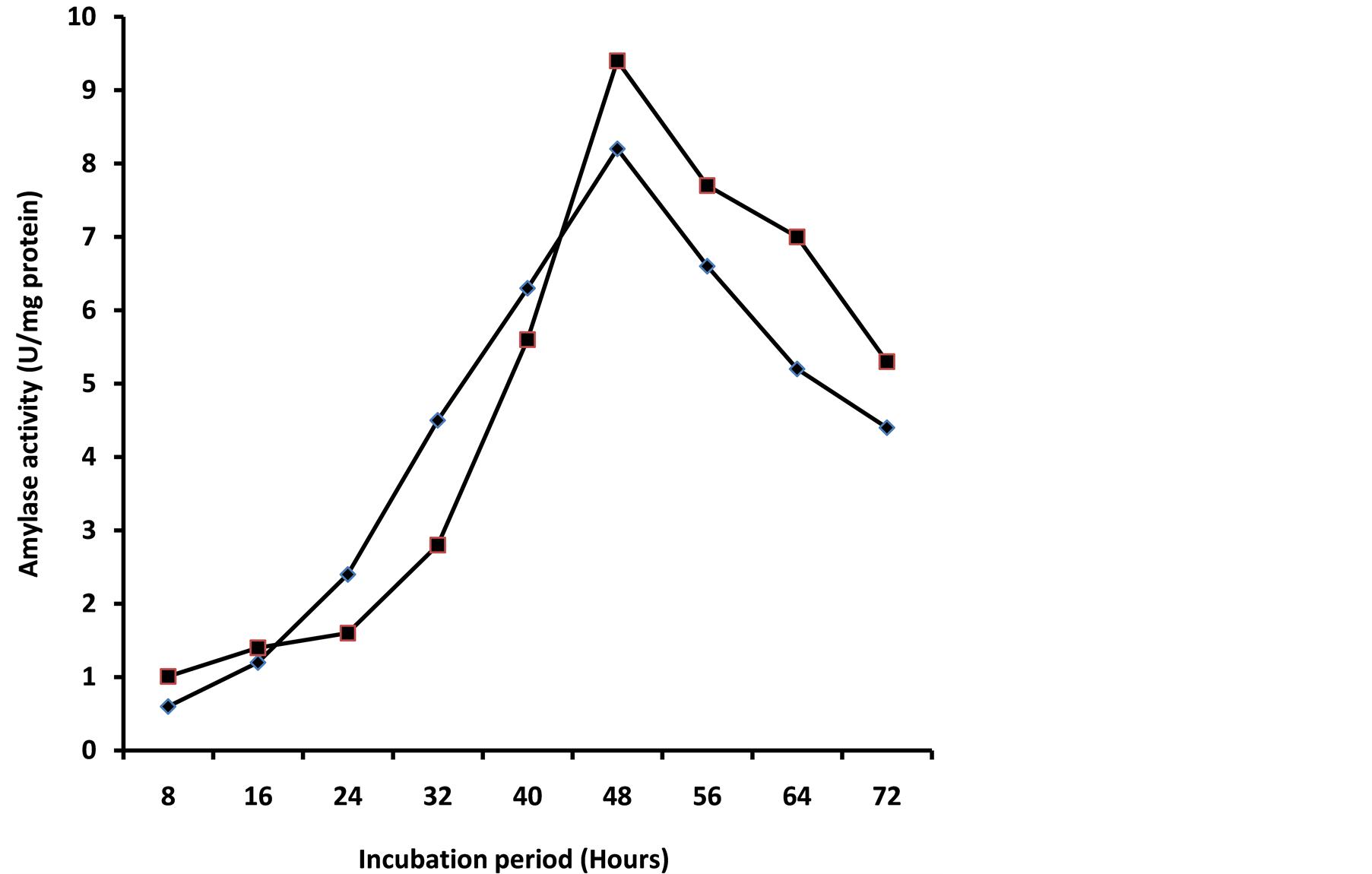
Figure 1. Growth progression and glucoamylase production from a mesophilic strain of A. niger in mineral salts medium supplemented with sweet potato or soluble starch. Amylase activity in soluble starch medium (♦); amylase activity in sweet potato starch medium (■).
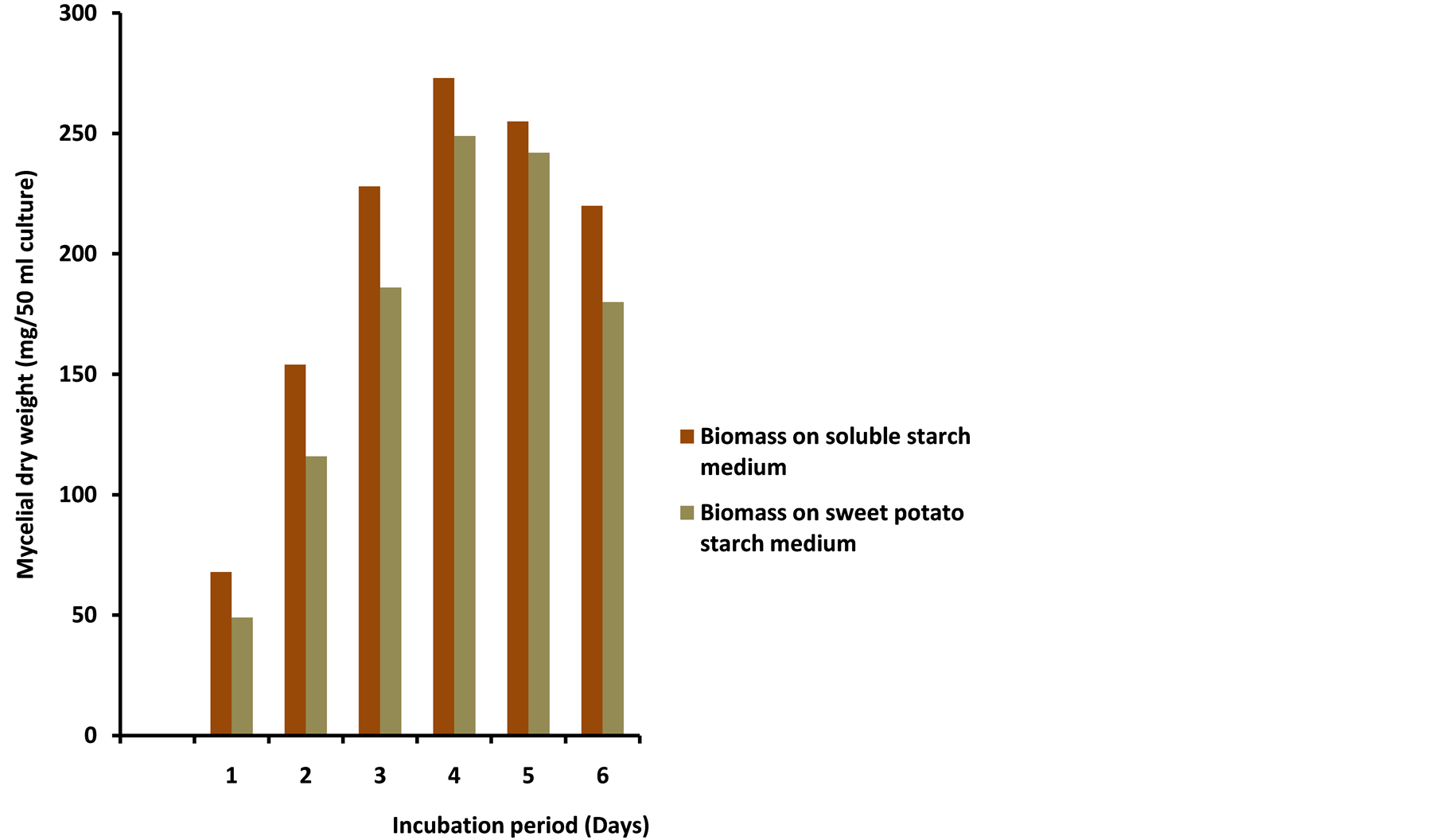
Figure 2. Effects of medium composition on growth and glucoamylase production by a mesophilic strain of A. niger on growth medium containing soluble starch and sweet potato starch.
the enzyme. Similarly, when soluble or sweet potato starch was added to the complete amylase growth medium, there was upward increase in biomass production to 494 and 418 mg. These values coincided with an increase in amylase production of 92 and 66 U/mg for soluble and sweet potato starch media respectively. The significance of this experiment is that for optimum growth and induction of amylase, mineral salts, nitrogen source and carbon source are of utmost importance. Although there is not much variation in the observed biomass produced in both media, amylase synthesis was higher in the soluble starch MS medium compared to sweet potato starch MS medium. It is therefore concluded that the ability of sweet potato starch MS medium to promote a comparable optimum biomass production and glucoamylase activity of 66 U/mg compared to 92 U/mg on soluble starch MS medium is remarkable in the quest to develop affordable local alternatives for industrial applications in the sub-Saharan Africa.
3.4. Physicochemical Properties
The pH of a growth medium plays an important role in enzyme secretion as they are sensitive to the concentration of hydrogen ions present. There exist a close relationship between amylase synthesis and the utilization of carbon source. pH variations during fermentation may give information about the amylase production, such as the start and the end of the enzyme production period and could also affect product stability in the medium [11] . In this study, the optimum pH for the activity of amylase was observed to be 4.5. The growth of A. niger on 2% (w/v) sweet potato or soluble starch containing medium at various pHs considered produced variable amylase activities. At pH 3.0, amylase activity of 1.4 ± 0.2 U/mg was recorded compared to pH 4.5 that recorded maximum activity of 9.1 ± 0.2 U/mg at the end of the fourth day of incubation on soluble starch medium (Figure 3). Growth on sweet potato starch medium compared favourably by producing an activity of 8.2 ± 0.4 U/mg at the end of the fourth day of incubation. Beyond the fourth day of the experimental period, activity decreased progressively. Increasing acidity and alkalinity was not favourable for the amylase production and activity for both media. Earlier studies have revealed that fungi required slightly acidic pH and that bacteria requires neutral pH for optimum growth [12] thus confirming the observed pH optimum of 4.5 obtained in this study for a mesophilic strain of Aspergillus niger. Several authors have also reported that fungi of Aspergillus sp. such as A. oryzae, A. ficuum and A. niger were observed to give significant yields of amylase at a pH range of 5.0 - 6.0 in submerged fermentation [13] [14] .
Temperature is critical and the influence of temperature on amylase production is related to the growth of the organism. Aspergillus niger grown on 2% (w/v) sweet potato starch medium produced an amylase activity of 8.1 U/mg at a temperature of 40˚C compared to 45˚C (6.2 ± 0.05 U/mg) at the end of the fourth day of incubation (Figure 4). Low levels of amylase activity were produced at a temperature of 25˚C (1.2 ± 0.001 U/mg) and 35˚C (4.1 ± 0.003 U/mg). But at 60˚C and 70˚C the activity dramatically dropped to 1.10 ± 0.001 U/mg and 0.8 ± 0.05 U/mg. Soluble starch medium produced a slight higher amylase activity of 9.2 U/mg at 40˚C and 1.30 ±
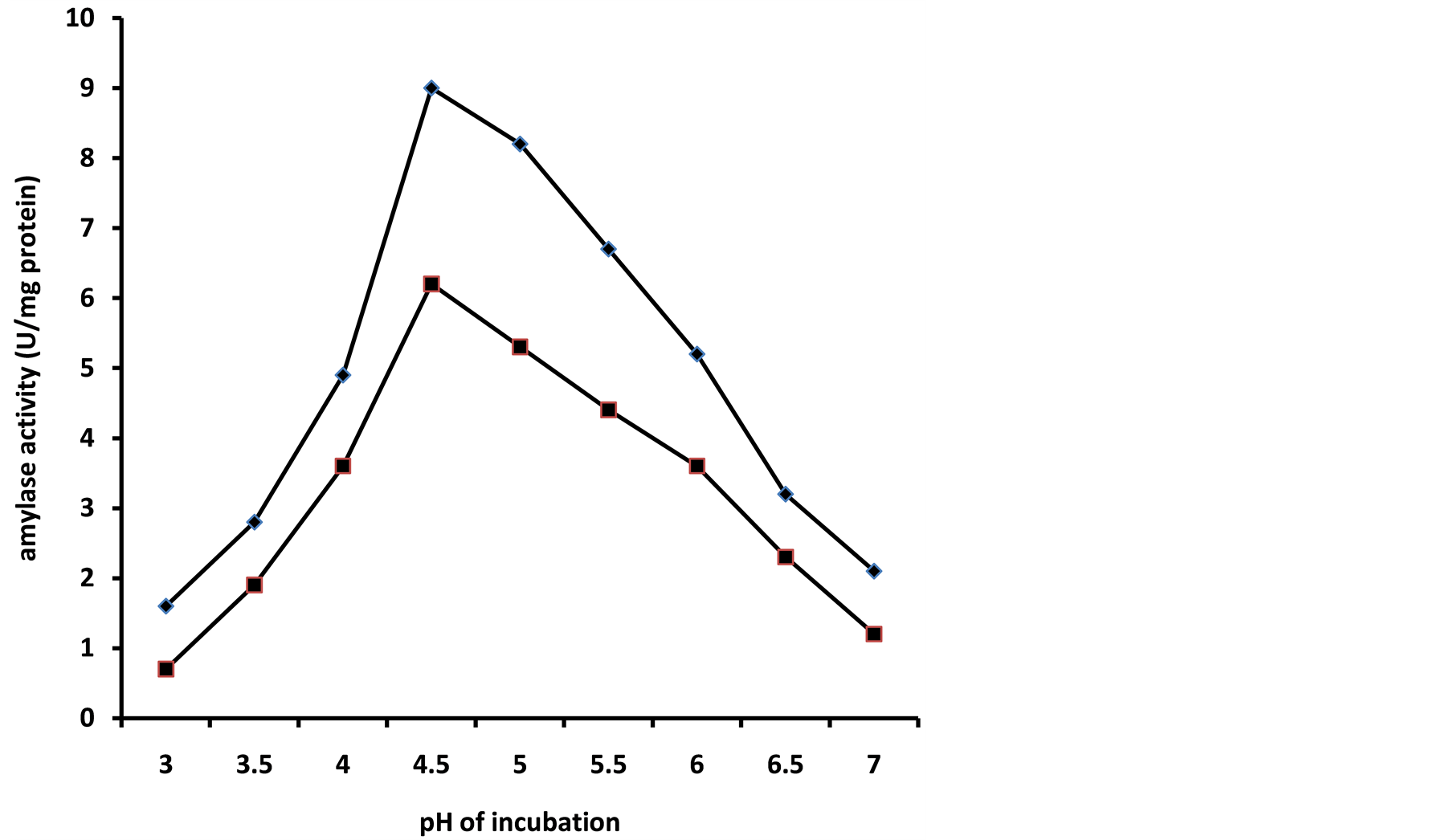
Figure 3. Effect of pH of growth medium on amylase synthesis by a mesophilic strain of A. niger grown on soybean meal and peptone. Amylase activity on soybean meal (♦) and amylase activity on peptone (■).
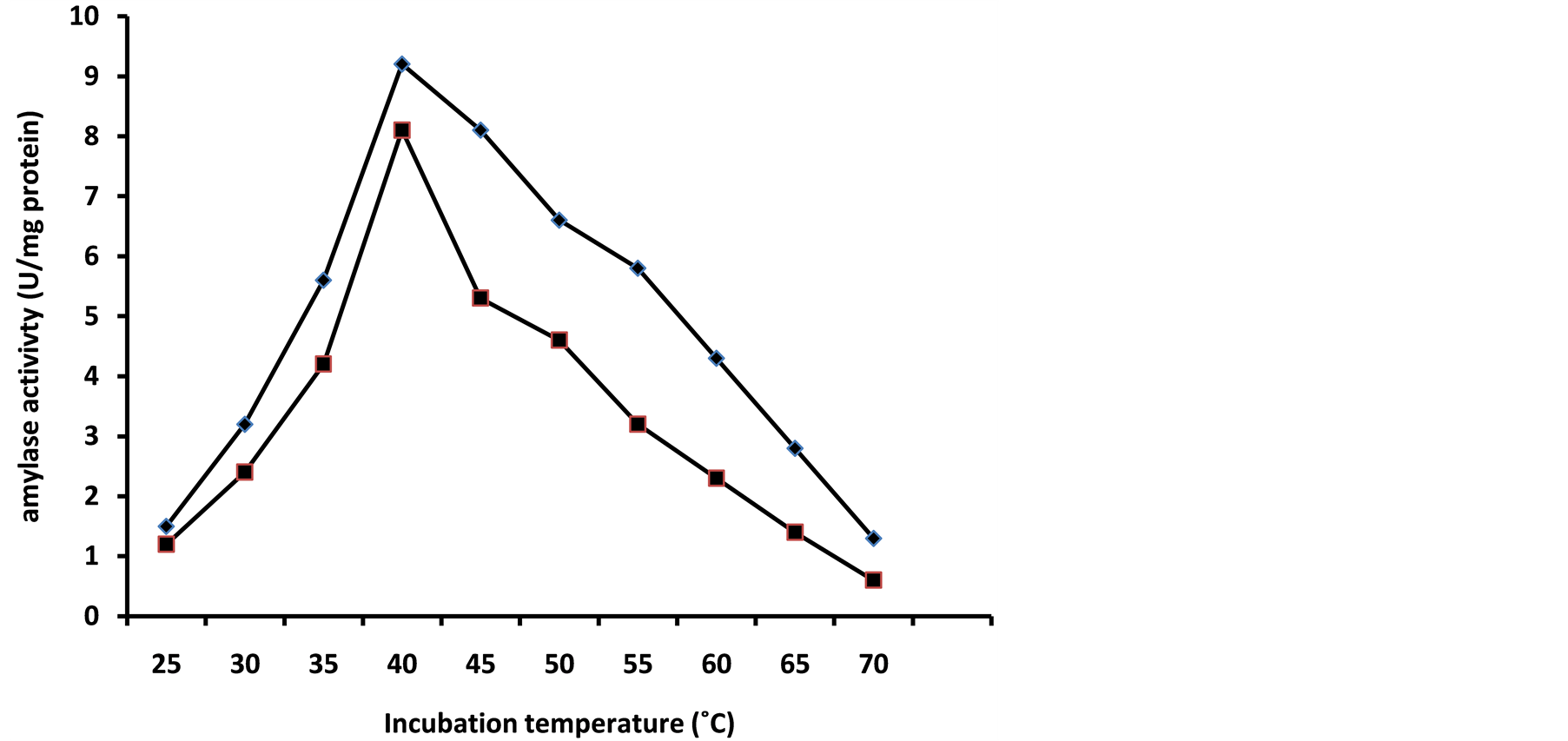
Figure 4. Effect of incubation temperature on growth and glucoamylase synthesis by a mesophilic strain of A. niger grown on sweet potato or soluble starch. Amylase activity on soluble starch medium (■); amylase activity on sweet potato starch medium (♦)
0.002 U/mg at 70˚C (Figure 4). The expression of low levels of activity at 35˚C and 75˚C could be attributed to poor growth of A. niger at these two temperature extremes. The observed optimum temperature of amylase production in this study is within the range of the findings of many authors in the literature [11] [15] -[17] .
The study demonstrated that locally available sweet potato starch could promote good yield of glucoamylase from A. niger. Although soluble starch medium yielded higher titres of the enzyme, there is a possibility of increasing amylase yield through medium optimization and provision of optimal growth conditions. Moreover, the strategy of the enzymatic hydrolysis of starch by the locally isolated novel enzyme opens up very interesting perspectives for industrial exploitation especially in the sub-Saharan Africa in view of the relative high cost of the conventional soluble starch. Another important point to be noted is that the bioprocess utilizing sweet potato starch is attractive due to its relatively cheaper availability throughout the year in Nigeria.
- Frost, G.M. and Moss, D.A. (1987) Production of Enzymes by Fermentation. In: Rehm, H.J., Reed, G., Eds., Biotechnology, Vol. 7a, VCH, Weinheim, 65-102.
- Kluyver, A.J. and Perquin, L.H.C. (1932) Zur Methodik der Schimmel-stoffwechseluntersuchung. Biochem Z, 266, 68-81.
- Schuster, E., Dunn-Coleman, N., Frisvad, J.C. and van Dijck, P.W.M. (2002) On the Safety of Aspergillus niger—A Review. Applied Microbiology and Biotechnology, 59, 426-435. http://dx.doi.org/10.1007/s00253-002-1032-6
- Castellonos, O.F., Sinitsyn, A.P. and Vlasenko, E.Y. (1995) Comparative Evaluation of Hydrolytic Efficiency toward Microcrystalline Cellulose of Penicillium and Trichoderma Cellulases. Bioresource Technology, 52, 119-124. http://dx.doi.org/10.1016/0960-8524(95)00011-3
- Fogarty, W.M. and Kelly, C.T. (1980) Amylase, Amyloglucosidase and Related Glucanases. In: Rose, A.H., Ed., Microbial Enzymes and Bioconversions, Vol. 5, Academic Press, London, 770-778.
- Chu, W.-H. (2007) Optimization of Extracellular Alkaline Protease Production from Ciacco, Fabricacao de amido e sua utilizacao. Secretaria da Industria, Comercio, Ciencia e Tecnologia, Sao Paulo.
- Barnett, H.L. and Hunter, B.B. (1972) Illustrated Genera of Imperfect Fungi. 3rd Edition, Burgess Publishing Co., Minneapolis, 241 p.
- Nelson, S. (1944) A Photometric Adaptation of the Somogyi Method for the Determination of Glucose. The Journal of Biological Chemistry, 153, 375-380.
- Sani, A., Awe, F.A. and Akinyanju, J.A. (1992) Amylase Synthesis in Aspergillus flavus and Aspergillus niger on Cassava Peel. Journal of Industrial Microbiology Biotechnology, 10, 55-59. http://dx.doi.org/10.1007/BF01583634
- Uguru, G.C., Akinyanju, J.A. and Sani, A. (1997) The Use of Yam Peel for Growth of Locally Isolated Aspergillus niger and Amylase Production. Enzyme and Microbial Technology, 21, 48-51. http://dx.doi.org/10.1016/S0141-0229(96)00225-6
- Gupta, R., Gigras, P., Mohapatra, H., Goswami, K.V. and Chauhan, B. (2003) Microbial α-Amylases: A Biotechnological Perspective. Process Biochemistry, 10, 1-18.
- Fogarty, W.M. (1983) Microbial Amylases. In: Fogarty, W.M., Ed., Microbial Enzyme and Biotech, Applied Science Publishers, London, 1-90
- Carlsen, M., Spohr, A.B., Nielsen, J. and Villadsen. J. (1996) Morphology and Physiology of an α-Amylase Producing Strain of Aspergillus oryzae during Batch Cultivations. Biotechnology and Bioengineering, 49, 266-276. http://dx.doi.org/10.1002/(SICI)1097-0290(19960205)49:3%3C266::AID-BIT4%3E3.0.CO;2-I
- Djekrif-Dakhmouche, S., Gheribi-Aoulmi, Z., Merihi, Z. and Bennamoun, L. (2005) Application of a Statistical Design to the Optimization of Culture Medium for α-Amylase Production by Aspergilus niger ATCC 16404 Grown on Orange Waste Powder. Journal of Food Engineering, 73, 190-197. http://dx.doi.org/10.1016/j.jfoodeng.2005.01.021
- Oguntimein, G.B. (1993) Growth and Amylase Production by Bacillus licheniformi Isolated from Cassava Processing Waste. Nigerian Food Journal, 11, 60-67.
- Cordeira, C.A.M., Martins, M.L.L. and Luciano, A.B. (2002) Production and Properties of α-Amylase from Thermophilic Bacillus sp. Brazilian Journal of Microbiology, 33.
- Gomes, E., de Souza, S.R., Grandi, R.P. and Da Silva, R. (2005) Production of Thermostable Glucoamylase by a Newly Isolated Aspergillus flavus A1.1 and Thermomyces lanuginosus A 13.37. Brazilian Journal of Microbiology, 36, 75-82.