Open Journal of Forestry
Vol.06 No.05(2016), Article ID:70376,18 pages
10.4236/ojf.2016.65031
Floristic Diversity of Forests of the Northwest Republic of the Congo
Félix Koubouana1, Suspense Averti Ifo2, Jean-Marie Moutsambote1, Romarick Mondzali-Lenguiya1
1ENSAF, BP 69 Université Marien Ngouabi, Brazzaville, Congo
2ENS, BP 69 Université Marien Ngouabi, Brazzaville, Congo

Copyright © 2016 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: July 8, 2016; Accepted: September 3, 2016; Published: September 6, 2016
ABSTRACT
The study of structural and tree flora biodiversity of the forest of the Mbomo-Kellé UFA was done on an area of 88.5 ha (0.014% of the total UFA area). The inventory was carried out in plots of 200 m × 25 m (0.5 ha), arranged in a contiguous way along the opened trails in the forest with an equidistance interval of 2 km. 11,012 trees of DBH ≥20 cm were counted, divided into 107 genera, 31 families and 133 species including 121 completely identified and 12 defined only in terms of genus. Data analysis consisted of calculating relative and weighted frequencies, the Shannon and Weaver diversity indexes, Pielou’s evenness index, and the Jaccard and Sorensen similarity indices. Relative frequencies showed that the most representative family in terms of species is the Fabaceae-Caesal piniodeae with 18.05% of species and the most representative in terms of individuals is the Burseraceae with 19.17% of inventoried trees. The principal components analysis allowed to show inter- and intra- block heterogeneities.
Keywords:
Floral Diversity, Management, Floristic Composition, Floristic Heterogeneity, Mbomo-Kellé UFA, Congo

1. Introduction
Tropical forests are the subject of several studies to better understand the role they could play in sustainable development, climate change, and floristic biodiversity. Tropical forests are characterized by high biodiversity, this diversity is an indicator that allows to appreciate links between the richness and the abundance of individuals; it reflects the degree of heterogeneity or stability of vegetation (Basnet, 1992; Trichon, 1997) . Plant diversity can therefore integrate several parameters such as the vegetation structure and dynamics (Moutsamboté, 1985; Koubouana, 1992; Koubouana, 1993) , In the Republic of Congo, according to the current national definition of the forest, forests cover 69% (CNIAF, 2015) of the territory. Sustainable management of these forests requires a sound knowledge of all the natural forest resource; this knowledge can be relied upon only by field studies of the forest environment. Vegetation studies lead to either conducting physiognomic research of the architectural type, either to identifying a number of representative vegetation parameters, allowing to define it simply, in order to compare it to others vegetations (Lescure et al., 1988) . For the present study, the second approach was used, that of the floristic and structural parameters. Long-term biodiversity conservation depends basically on knowledge of the structure, the species richness and the ecological characteristics of the vegetation.
Many studies have been made in Republic of Congo on improving the knowledge of the plant resource. But this knowledge remains generally piecemeal and predominantly localized in protected areas and logging forest concessions. These previous studies are related to ethnobotanical characteristics and general knowledge of the flora (Cusset, 1980; Adjanohoun et al., 1988; Cusset, 1989; Hecketsweiler, 1990; Hecketsweiler & Mokoko-Ikonga, 1991; Dowsett & Dowsett, 1991; Dowsett, 1991; Doumenge, 1992; Kimpouni et al., 2007; Banzouzi, 2008) ; for the massif of Mayombe (Koubouana & Moutsamboté, 2006) in the massif of Chaillu (Mondzali-Lenguiya, 2008; Moutsamboté, 2012) , for the vegetation of the Northern Congo. As far as concerns us, there are not enough available data on the forest of the central western parts of Republic of Congo. These forests are poorly known in Republic of Congo.
This work contributes to the knowledge of this part of the Congolese forest. Mbomo- Kellé UFA , by its geographical location, could be a transition zone between the flora of southern UFA and the UFA of Northern Congo. This study has as its main objective, to improve the knowledge of the floristic resource of the Mbomo-Kellé UFA, and as specific objectives to: (i) conduct a floristic analysis of the vegetation of the Mbomo-Kellé UFA from three blocks of inventory data; (ii) characterize the UFA’s flora, through the calculation and analysis of diversity indexes (iii) make an analysis of floristic heterogeneity intra- and inter-blocks of inventory from inventory track data.
2. Material and Methods
2.1. Study Site
Experimental plots were installed within the Mbomo-Kéllé forest management unit (UFA) in the Department of Cuvette Ouest between 14'00'' E and 15˚00''E; 00˚27'35.9'' North and 00˚22'03'' South. It is limited to the North by the Republic of Gabon, to the Northeast by the Odzala-Kokoua National Park, to the South by the Tsama-Bamana UFA, to the West and South-West by the Republic of Gabon, and to the East by the Tsama-Bamana UFA (Figure 1). The Mbomo-Kéllé UFA is crossed by the Equator and under the influence of the Congolese equatorial climate. The average annual rainfall is about 1900 mm/year. The average temperature is about 25˚C, the warmest month is March (with 25.9˚C) and the coldest month is May (with 20.26˚C). Annual average

Figure 1. Map of limits and geography situation of Mbomo-Kellé UFA.
relative humidity is 84%, with a monthly average maximum relative humidity of 97.33% observed in June and a minimum average monthly relative humidity of 59% observed in September.
The study area is characterized by high hills whose altitude varies between 350 and 680 m, which extend a large area of the border of the Congo and Gabon. This relief extends continuously from the Mbomo-Kellé UFA to the Mbinda area in the Chaillu massif. The river network of the Mbomo-Kellé UFA is organized around two large rivers including the Lebango tributary North of the UFA and the Lekona tributary to the South. All the rivers that are in the UFA are either the Lebango or the Lekona tributaries. The two rivers flow into the Likouala-Mossaka in Itoumbi.
2.2. Data Collection
For a better inventory of field data, Mbombo-Kellé UFA was subdivided into 13 blocks of inventory in which plots were installed to inventory all trees o DBH ≥20 cm. For this work, we used data obtained from three blocks of inventory traversing South UFA (blocks 9, 4 and 8) (Figure 2). The size of each plot was of 200 m × 25 m, i.e. 0.5 ha. A

Figure 2. Map of the vegetation of UFA Mbomo-Kellé.
total of 88.5 ha were inventoried, distributed unevenly in the three blocks with 61.5 ha in block 4, 18 ha in block 8 and 9 ha in block 9. All trees of DBH ≥ 20 cm were identified and their diameter at 1.30 m measured.
2.3. Data Analysis
Floristic diversity
Diversity was determined by using the Shannon Index where pi is the frequency of species I (Ni ⁄N), Ni the number of individuals of species i, and N the number of individuals of all the species.

The Pielou or evenness index
Is the ratio of the Shannon and Weaver index of diversity on the maximum diversity index. It marks the level of diversity achieved by the system compared to the maximum possible (Kimpouni et al., 2013) .

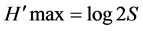
H' max is the logarithm of the total number of species (S):
S = total number of species; NI = number of individuals of a species in the sample;
When the index is low, a phenomenon of dominance is revealed, while when it is high, it marks regular compensation of individuals among species (Kimpouni et al., 2013) .
Principal components analysis
This multivariate analysis is used to discriminate between the blocks and the inventoried trails according to edaphic affinities.
2.4. Similarity
Similarity is determined by the coefficients of similarity. These coefficients are used to measure the association between objects. The similarity of two samples (floristic surveys) is based on the presence or absence of certain species in the two samples (Legendre & Legendre, 1984a) . To study the similarity of our different floristic surveys, we used two binary factors excluding the double zeros; i.e. the coefficient of Sorensen (K) and the coefficient of Jaccard (S).
The SØrensen coefficient provides a two times higher weight to double presence, we can consider the presence as more informative than its absence (Legendre & Legendre, 1984b) .
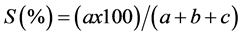
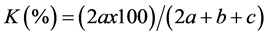
a = number of common presence for both floristic surveys
b = number of presence for the first floristic survey
c = number of presence for the second floristic survey
d = number of species absent in both floristic surveys
According to Legendre & Legendre (1984b) , the SØrensen coefficient is dull compared with the Jaccard coefficient, that is, if the similarity of a pair of objects computed by the Jaccard coefficient is higher than the similarity of another pair of objects, it will be also higher if using the coefficient of Sorensen for the calculation of similarity.
3. Results
Floristic richness
The floristic inventory carried out in blocks4, 8 and 9 of the Mbomo-Kellé UFA allowed to record11012 trees of DBH ≥20 cm, divided into 34 families, 107 genus and 133 species, including 12 species defined only as shown in Table 1.
The calculation of gross and weighted frequencies allowed us to appreciate the wealth specific (in number of species) and weight (in number of trees of DBH ≥20 cm) per family (Figure 2).
It appears in Figure 3 that the Fabaceae-Caesalpinioideae family (18.05%) is the most abundantly represented in number of species in the surveyed zone, followed by the Meliaceae (7.52%), Fabaceae Mimosoideae (6.02%), Eupho-rbiaceae (5.26%), Annonaceae (4.51%) and the \Myristicaceae (3.76%).

Table 1. Floristic composition of the study area.

Figure 3. Floristic richness and weight of different inventoried families.
If we consider the quantitative aspect, it is the Burseraceae family (19.01%) which is the most represented, followed by the Fabaceae-Caesalpinioideae (15.96%) (16.09), Myristicaceae (13.18%), Annonaceae (9.49%), Euphorbiaceae (8.32%), and Fabaceae Mimosoideae (7.32%).
The quantitative abundance of Burseraceae, Myristicaceae and Annonaceae is explained by the strong representation respectively of species Santiria trimera (14.57%), Scyphocephaluim mannii (7.55%) and Greenwayodendron suaveolens (4.30%).
The Shannon-Weaver and Pielou evenness indexes were calculated in each block considering the number of individuals per species and the total number of individuals of each block. The results showed that the index of Shannon and Weaver varies between 5 and 6 bits, and evenness between 0.4 and 0.6 (Figure 4).

Figure 4. Values for the Shannon and Weaver index and the Pielou evenness index.
These indexes did not vary significantly between inventory blocks. It should be noted that through both the relative frequencies that index calculated, neither of these settings show inter and intra-bloc heterogeneity.
Heterogeneity between blocks
Inter-block heterogeneity was studied from a principal component analysis made on a presence-absence basis, by observing encountered species and as variable or column represented by the different inventory blocks Table 2 shows the contribution of the variables on the factor axes.
This table shows that the essential information on the distribution of the variables in the correlation circle (Figure 5) is explained by F1 and F2 axes.
Figure 5 shows that all the variables, i.e. all the blocks, are arranged in the positive side of the F1 axis. This could explain the association of the inventory blocks with a same forest type. This result is confirmed by the calculated similarity index values (Figure 6).
Indeed, the values of the Jaccard and Sorensen indexes are greater than 60%, indicating a similarity between inventory blocks. However, this similarity cannot hide the local variations of flora. So we note that on the F2 axis, there are two groups: the Group made of block 4, and group 2 made by blocks 8 and 9. Block 4 is opposed to block 9. Indeed, block 4 was located in a mostly surface area or barely marshy, while in block 9, many trails cross swampy areas or follow the banks of the rivers where characteristic wetland species are present, such as Sarcocephalus pobeguinii Pobeg., Coelocaryon botryoides Vermoesen, Xylopia staudtii Engl. & Diels and Alstonia congensis Engl. Many trails of block 8 cross fallow areas, where species include Dacryodes edulis (G. Don.) H.J. Lam, and many shoots of Musanga cecropioides R. Br. The factorial axis F2 discriminates against our blocks according to not only the local substrate variability, but also according to the degree of human activity in the forest.

Figure 5. Distribution of variables (blocks) in the correlation circle.

Figure 6. Jaccard and Sorensen similarity Indexes.
Table 2. Contribution of variables (in %) to the total inertia.
Intra-block heterogeneity
Theoretically, inter-block heterogeneity would arise from intra-block heterogeneity. To test this hypothesis, we performed a principal component analysis (PCA) on the basis of an array of presence-absence of species on the distance of each block. Obviously, it is interesting to a PCA in the blocks where there were more than two past inventoried i.e. blocks 4 and 8.
Heterogeneity in block 4
With regard to block 4, Table 3 indicating the contribution of the variables represented by the trails in the different factorial axes, shows that the essential information is represented in the F1 - F2 (Figure 7) plan below.
This figure shows considering axis F1, all the trails are the positive side of the axis indicating that they belong to a single block of inventory. However on the F2 axis, two groups of Trails are distinguished: the trails 5- 6 Group and the trails 2-3-4 Group. Indeed, the F2 axis discriminates between trails depending on the nature of the substrate. The distance of the first group have not gone into a swamp of hydromorphic soils while those in the second group cut the marshy areas. This is explained by the presence of species such as Sarcocephalus pobeguinii. Pobeg, Xylopia staudtii Engl. & Diels, Alstonia congensis Engl., Hallea stipulosa (DC.) Leroy. These results are confirmed by the values of the index of similarity (Figure 8).
It appears on this figure that the values of the index of similarity are low between two different trails of different groups whilst the values are high for trails of the same group.

Figure 7. Distribution of variables (trails) in the circle of correlation in block 4.

Figure 8. Index of similarity of Jaccard and Sorensen’s for block 4.
Table 3. Contribution of the variables (in %) to the total inertia.
Heterogeneity in block 8
Figure 9 shows the Trail distribution in block 8.
The point made in block 4 is valid for block 8. Indeed, all trails are arranged in the positive side of the F1 axis, indicating their belonging to a single block. In the F2 axis, trails 4 and 7 are floristically close because they are situated on the positive side of the F2 axis. Trail 6, located on the negative side of the F2 axis, contains species that are characteristic of marshy areas.
4. Discussion
By comparing the results obtained in the study’s zone and those obtained in the UFA of the North of the Republic of Congo, in particular the UFA Pokola and Missa, it appear to be more higher than those of the other UFA of the North of Congo. The forest of the UFA Mbomo-Kellé is floristiquement richer than of the UFA Missa.
The six most representative families in terms of species in the forest of the Mbomo-Kellé UFA are those of: Fabaceae-Caesalpinioideae with 18.05%, Meliaceae 8.27%, Euphorbiaceae 6.02%, Fabaceae-Mimosoideae 6.02%, Annonaceae 4.51% and 4.51% Sapotaceae. These results are similar with those of Mondzali-Lenguiya (2008) observed in the forest of the Missa UFA where three families have a high percentage of representation (Table 4).

Figure 9. Distribution of variables (trails) in the circle of correlation in block 8.
Table 4. Comparison of the six richest families of the Mbomo-Kellé UFA and the Missa UFA.
Fabaceae-Caesalpinioideae species richness appears to be a commonality of Congolese forests because Kimpouni et al. (2007) also note a gross frequency of 18.05% in the forest near Tobin on the Congolese coast.
The forest species density studied varies between 0.01 tree∙ha−1 to 18.12 tree∙ha−1 and the overall density of the forest is 124.43 tree∙ha−1. This result is similar to that obtained in the forest of the Ngombe UFA by IFO company (2007). Indeed, the logging concession company IFO found a density of 124.13 tree∙ha−1 for the Ngombe UFA. However this result is slightly lower than the density in the forest species of the Missa UFA which is of 189.001 tree∙ha−1 calculated in relation to the data of Mondzali-Lenguiya (2008) .
Quantitatively, the forest of the Mbomo-Kellé UFA showed a preponderance of individuals from the family Burseraceae, Fabaceae-Caesalpinoideae, Euphorbiaceae and Myristicaceae. These results are similar to those observed in the Boubissi UFE in the Mayombe (located in South of Republic of Congo) by reflecting its character of transition between the forests of the South and North Republic of Congo.
Weighted spectrum shows that in addition to the Fabaceae-Caesalpinioideae, the families of the Burseraceae (19.17%), of the Myristicaceae (13.29%) and of Euphorbiaceae (9%) are the most represented in the number of trees regardless of the inventoried block. These high values are dependent on the abundance of the species respectively Santiria trimera, Scyphocephalium mannii and Plagiostyles africana as we found Kimpouni et al. (2007) in the forests of the Bambama UFA (in the South-Congo) thereby reinforcing the transition forest characteristic of the Mbombo-Kellé UFA, between the forests of the South and North Congo forests.
The floristic variability or the floristic heterogeneity inter-block was studied from the factorial analysis of correspondence. The results showed on the one hand that the sampled blocks of inventory belong to the same forest type and on the other hand which there are floristic variabilities due to the variation of the substrate, like had underlined it (Basnet, 1992; Gartlan et al., 1986) . The membership of the same forest type is confirmed by the values of indices of similarity. This heterogeneity inter-block is also observed in the tailboards of the same block. The variability of the edaphic factors (composition of the grounds, grounds dewatered and hydromorphic grounds) explains the richness of the forest of the UFA Mbomo-Kellé.
The floristic variability or the floristic heterogeneity between-block was studied starting from the factorial analysis of correspondence. The results showed on the one hand that the sampled blocks of inventory belong to the same forest type and in the other hand which there are floristic variabilities due to the variation of the substrate, like had underlined it (Basnet, 1992; Gartlan et al., 1986) . The various blocks belong to the same type of plot highlighted by the indices of similarities. This heterogeneity inter-block is also observed in the tailboards of the same block. The variability of the edaphic factors (composition of the grounds, grounds dewatered and hydromorphic grounds) could explained the richness of the forest of the UFA Mbomo-Kellé.
5. Conclusion
The floristic inventory of the Mbomo-Kéllé UFA forest allowed us to appreciate its floristic composition. Qualitatively, all inventoried blocks were rich in Fabaceae- Caesalpinoideae, Meliaceae, Myristicaceae, Euphorbiaceae, Fabaceae Mimosoideae, Burseraceae and Annonaceae. Shannon-Weather calculated index values are high, around 6-bit indicating a recovery of the same importance for most of the inventoried species. However, the Pielou evenness index was relatively low in block 4 indicating a low abundance of some species; these include the Santiria trimera (Oliv.) Aubrév. Santiria trimera (Burseraceae), Plagiostyles africana (Mull. Arg.) Prain Plagiostyles africana (Euphorbiaceae), Scyphocephalium mannii (Beth.) Warb. Scyphocephalium mannii (Myristicaceae). The results of the principal components (PCA) analysis revealed floristic heterogeneities within blocks. Regarding the inter-block variance, the results of the PCA have shown that three blocks belong to a same ecosystem. What was confirmed by the indices of similarity of Jaccard and Sorensen. The PCA also indicated the floristic differences between blocks due to the local variability of the substrate. As regards inter-block heterogeneity, the factorial axes of the PCA have discriminated between Trails of blocks 4 and 8 depending on the nature of the substrate. Some past crossed swampy areas of hydromorphic soils. Scyphocephalium mannii is present in the forests of Southern Congo in the Chaillu massif and is reported in the forests of Northern Congo in the Mbomo-Kellé UFA in Odzala National Park and in parts of the Western Sangha. This floristic fact shows that the Mbomo-Kéllé UFA is a transition zone between the South-Congo forests and the forests of Northern Congo. This observation is confirmed by the similarity of relief between the Chaillu massif and the Mbomo-Kellé UFA.
Structurally, the density of trees in the surveyed area is 124.42 trees/ha. Weight frequency showed a preponderance of trees from the families Burseraceae, Myristicaceae, and Euphorbiaceae.
Cite this paper
Koubouana, F., Ifo, S. A., Moutsambote, J.-M., & Mondzali- Lenguiya, R. (2016) Floristic Diversity of Forests of the Northwest Republic of the Congo. Open Journal of Forestry, 6, 386- 403. http://dx.doi.org/10.4236/ojf.2016.65031
References
- 1. Adjanohoun, E. J., Ahyi, A. M. R., Ake Asi, L., Baniakina, J., Chibon, P., Cusset, G., Doulou, V., Enzanza, A., Eyeme, J., Goudote, E., Keita, E., Mbemba, C., Mollet, J., Moutsamboté, J. M., Mpati, J., & Sita, P. (1988). Contribution aux études ethnobotaniques et floristiques en République populaire du Congo : médecine traditionnelle et pharmacopée (605 p). Paris: ACCT. [Paper reference 1]
- 2. Banzouzi, F. (2008). Inventaire d’aménagement de l’UFE Boubissi: Végétation et flore du Bloc B (46 p). Rapp. de fin de formation, IDR, Brazzaville: Univ. Marien Ngouabi. [Paper reference 1]
- 3. Basnet, K. (1992). Effect of Topography on the Pattern of Trees in Tabanuco (Dacryodes Excels) Dominated Rain Forest or Puerto-Rico. Biotropica, 24, 31-42. [Paper reference 3]
- 4. CNIAF (2015) Carte de changement de couverture forestière en République du Congo pour la période 2010-2012. [Paper reference 1]
- 5. Cusset, G. (1980). Etude phytoécologique préliminaire de la forêt Yombé (78 p). Université paris VI et Eq. Biol. For. Trop. Dimonika. [Paper reference 1]
- 6. Cusset, G. (1989). La flore et la végétation du Mayombe congolais, état des connaissances. In J. Sénéchal et al. (Eds.), Revue de connaissances sur le Mayombe (pp. 103-136). Paris: Unesco. [Paper reference 1]
- 7. Doumenge, C. (1992). La réserve de Conkouati, Congo: le secteur sud-ouest. UICN, Chevron, Gland, Suisse, 213 p. [Paper reference 1]
- 8. Dowsett, R. J., & Dowsett-Lemaire, F. (1991). Flore et faune du Bassin du Kouilou (Congo) et leur exploitation. Tauraco Research Report (CONOCO), 4, 340 p. [Paper reference 1]
- 9. Dowsett-Lemaire, F. (1991). The Vegetation of the Kouilou basin in Congo. In R. J. Dowsett, & F. Dowsett-Lemaire (Eds.), Flore et faune du Bassin du Kouilou (Congo) et leur exploitation. Tauraco Research Report, 4, 17-51. [Paper reference 1]
- 10. Gartlan, J. S., Newbery, M., Thomas, D. W., & Waterman, P. G. (1986). The Influence of Topography and Soil Phosphorus on the Vegetation of Korup Forest Reserve, Cameroun. Vegetation, 65, 131-148.
http://dx.doi.org/10.1007/BF00044814 [Paper reference 2] - 11. Hecketsweiler, P., & Mokoko-Ikonga, J. (1991). La réserve de Conkouati, Congo: Le secteur sud-est (323 p). Gland: UICN. [Paper reference 1]
- 12. Hecketsweiler, P. (1990). La conservation des écosystèmes forestiers du Congo. Gland: UICN. [Paper reference 1]
- 13. Kimpouni, V., Loumeto, J., & Mizingou, J. (2007). Caractérisation de l’okoumé (Aucoumea klaineana) dans la forêt du littoral congolais. DGRST, Actes des premières journées nationales sur la recherche forestière au Congo, Brazzaville, 26-29 juin 2006. [Paper reference 3]
- 14. Kimpouni, V., Mbou, P., Gakosso, G., & Motom, M. (2013). Diversité floristique du sous-bois et regeneration naturelle de la forêt de la Patte d’Oie de Brazzaville, Congo. International Journal of Biological and Chemical Sciences, 7, 1255-1270.
http://dx.doi.org/10.4314/ijbcs.v7i3.31 [Paper reference 2] - 15. Koubouana, F., & Moutsambote, J.-M. (2006). Etude préliminaire de la végétation des UFE Tsinguidi et Bambama (42 p). Brazzaville: Rapp. D’étude. [Paper reference 1]
- 16. Koubouana, F. (1992). Etude comparée de plusieurs types d’analyses spatiales sur les forêts semi-décidues de la Vallée du Niari (CONGO). Bulletin d’écologie, 23, 163-174. [Paper reference 1]
- 17. Koubouana, F. (1993). Les forêts de la Vallée du Niari (Ongo): Etudes floristique et structurales (265 p). Thèse doct, Paris: University of Paris VI. [Paper reference 1]
- 18. Legendre, L., & Legendre, P. (1984a). Ecologie Numérique, tome 1: Le traitement multiple des données écologiques (2rd éd., 260 p). Paris: Masson. [Paper reference 1]
- 19. Legendre, L., & Legendre P. (1984b). Ecologie Numérique, tome 2: La structure des données écologiques (2rd éd., 335 p). Paris: Masson. [Paper reference 2]
- 20. Lescure, J. P., & Boulet, R. (1985). Relationship beetwen Soil and Vegetation in a Tropical Rain Forest in Franch Guiana. Biotropica, 17, 155-160.
http://dx.doi.org/10.2307/2388508 [Paper reference 1] - 21. Mondzali-Lenguiya, R. (2008). Inventaire d’aménagement de l’UFA Missa: aspect floristique des zones 1,2,3 and 4 (43 p). Rapport de stage de fin de formation en vue de l’obtention du diplome d’ingénieur des travaux de développement rural. Brazzaville: technique forestière, Institut de Développement Rural (IDR), Université Marien Ngouabi. [Paper reference 3]
- 22. Moutsamboté, J.-M. (1985). Dynamique de reconstitution de la forêt Yombe (Dimonika, R.P. du Congo) (301 p). Bordeau: These 3 cycle University of Bordeaux III. [Paper reference 1]
- 23. Moutsamboté, J.-M. (2012). Etude écologique, phytogéographique et phytosociologique du Congo septentrional (Plateaux, Cuvettes, Likouala et Sangha) (632 p). Thèse de Doctorat d’Etat, Brazzaville: University of Marien Ngouabi. [Paper reference 1]
- 24. Trichon, V. (1997). Hétérogénéité spatiale des forêts d’une forêt tropicale humide de Sumatra: Effet de la topographie sur la structure floristique. Annals of Forest Science, 54, 431-446.
http://dx.doi.org/10.1051/forest:19970502 [Paper reference 1]