Open Journal of Immunology
Vol.3 No.4(2013), Article ID:40698,3 pages DOI:10.4236/oji.2013.34028
Isolation of γδT cells from mouse small intestine
![]()
Mechnikov Institute of Vaccine and Sera of RAMS, Moscow, Russia; *Corresponding Author: hell59@mail.ru
Copyright © 2013 Nadezda Snegireva et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights © 2013 are reserved for SCIRP and the owner of the intellectual property Nadezda Snegireva et al. All Copyright © 2013 are guarded by law and by SCIRP as a guardian.
Received 8 October 2013; revised 8 November 2013; accepted 15 November 2013
Keywords: Gamma/Delta T Cells; Small Intestine; Intraepithelial Lymphocytes; Magnetic Separation
ABSTRACT
Gamma/delta T cells (γδТ cells) are among the first lymphoid cells appearing in the ontogenesis. Many of γδТ cells are located in the small intestine/lamina propria of mice and human, where they cooperate with different cells (B-1 cells, in particular) and influence their activity. However, such interactions are studied rather scanty, and the functional role of γδТ cells in the intestine is not yet fully elucidated. To study the interactions of mouse γδТ cells with other cells it is necessary to have purified cell populations. Unfortunately, most approaches used for isolation of human γδТ cells are not suitable for isolation of mouse γδТ cells. The aim of the present study is the modification of the method of mouse intestinal γδТ cell isolation in the quantities sufficient for the in vivo and in vitro experiments.
1. INTRODUCTION
Gamma/delta T cells (γδТ cells) are among the first lymphoid cells appearing in ontogenesis. Many of them reside at the basolateral surfaces of intestinal epithelial cells [1]. From alpha/beta T lymphocytes, appearing a little later, γδТ cells differ by their capacity to recognize unprocessed antigens. It was shown that γδТ cells participated in the processes of oral tolerance [2] and possess immunoregulatory functions [3,4]. However, the interactions of γδТ lymphocytes with other cells are studied rather scanty, and their functional role in the intestine is not yet fully elucidated.
It was suggested that the switch from IgM to IgA in T-independent intestinal B-1 cells might be provided with γδТ cells factors. This suggestion was not experimentally checked, and the question remains open till now. The answer may be obtained by the study of the direct B-1 and γδТ cell interactions in vitro. To perform such experiments purified B-1 and γδТ cells should be used.
It is known that peritoneal B-1 cells may migrate to the gut where many of them become IgA-producers [5]. Thus, peritoneal cavity may be used as a source of B-1 cells. Method of B-1 cell isolation is well developed routine procedure. On the contrary, the isolation of γδТ cells from mouse intestine meets with significant difficulties. We tried to modify and simplify the method of mouse intestinal γδТ cell isolation.
2. MATERIALS AND METHODS
2.1. Mice
Female CBA mice, 16 - 18 g were used. Mice were maintained in the animal house of Mechnikov Institute of Vaccines and Sera of Russian Academy of Medical Science.
2.2. Isolation of γδТ Cells
Intraepithelial lymphocytes (IELs) were isolated from the small intestine; γδТ cells were isolated from IELs. To isolate IELs in some experiments collagenase of Clostridium histolyticum (Gibco) was used.
2.3. Flow Cytometry
The different cell populations were analyzed by cytometry. IELs and γδТ cells were prestained with antiCD16-32 mAb (2.4G2) to block FcγRII/RIII receptors and stained at 4˚C for 30 min with the following fluorochrome conjugated antibodies: FITC-anti-CD3 (17A2), PE-anti-CD19 (1D3), PE-anti-γ/δT (GL3) (all reagents, BD-Pharmingen).
After washing away the excess of reagents stained cells were resuspended in staining medium containing 5 μg/ml propidium iodide (PI) (Sigma). Cells stained with PI (dead cells) were excluded from analysis. Cells were analyzed on Beckman Coulter EPICS XL. Results were treated with the Programmed SYSTEM II (Beckman Coulter).
3. RESULTS AND DISCUSSION
Over the years, a number of strategies have been developed to obtain enriched preparations of intraepithelial lymphocytes (IELs) and γδТ cells. The method described are used mainly for isolation of human intestinal cells [6], although the methods of the isolation of IELs and T lymphocytes from monkey [7], rat [8], pig [9] and mice intestine are also described [10].
The most commonly used approaches involve the removal and washing of intestine, isolation of IELs and purification of T lymphocytes through Percoll density gradients. There are different modifications of these approaches concerning mainly the mode of tissue degradation, the time and the temperature of incubation of intestine segments with EDTA and dithiotreitol (DTT) and the concentrations of collagenase.
We have found that low viability of mouse IELs depended mainly on the high sensibility of intestinal mouse cells to the temperature and, especially, to the collagenase treatment. In addition, relatively many IELs (and γδТ cells, respectively) were lost during centrifugation in Percoll gradients. Therefore we tried to develop the method of IEL isolation without collagenase treatment and without cell purification through Percoll gradient. Below the protocol used in our experiments is written in details.
Mouse small intestine was washed by RPMI 1640 medium with 8% FCS on ice, everted and cut into several 2 - 3 cm segments to avoid winding. (Note! The intestine should not be minced or cut into very small pieces, otherwise the yield of cells would be greater but their viability would be less). The segments were placed into a 50-ml conical tube containing RPMI 1640 medium with 8% FCS, 1 mM EDTA, 1 mM DTT (Sigma), and 250 μl of penicillin + streptomycin (1/100) (Pen Strep, Gibco). The tube was shaken horizontally on an orbital shaker for 30 - 60 min at 37˚C. The tissue suspension was passed through 4 layers of gauze to remove mucus and centrifuged for 6 min. at 400 g. The yield of IELs CD3+ T cells and γδТ cells after different time of isolation is present in the Figure 1. It may be seen that the increase of the time of tissue incubation from 30 to 60 min. permits to isolate from the intestine of 3 mice about 38 - 40 × 106 cells instead of 1 - 2 × 106 after 30 min. incubation.
To purify γδТ cells commercial ТCR γ/δT cell Isolation Kit (Miltenyi Biotec) was used. The yield of CD3+ T cells in 2 experiments was equal to 27 and 20 mln, and the purity of CD3+ T cells was about 63% - 67%. The yield of γδТ cells was 1.0 - 1.2 × 106 (from
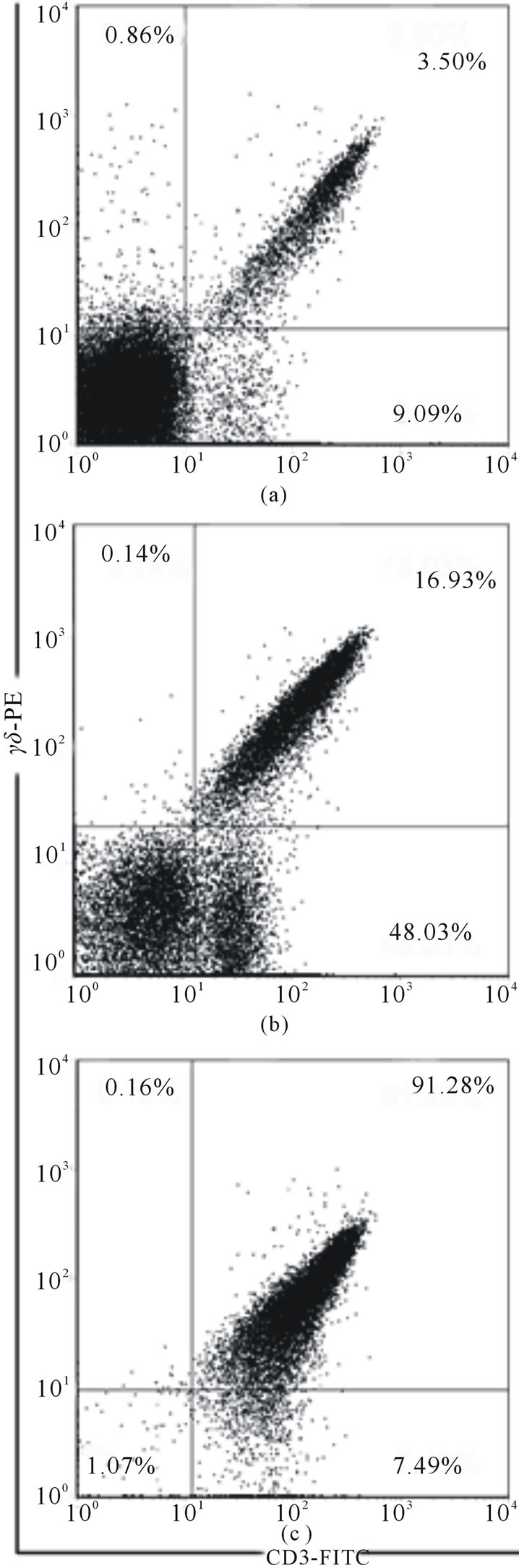
Figure 1. The percentage of γδТ cells at different stages: (a) after 30 min of incubation; (b) after 60 min of incubation; (c) after magnetic cell sorting.
intestines of 3 mice). After one-step purification the purity of γδТ cells was about 50%, but after two step purification it increased to 90% - 92%. The viability of cells was not less of 85%. These data do not differ from those recently published [11,12], where IELs were purified by centrifugation through Percoll gradient, and γδТ cells were isolated by immunomagnetic method. We believe that modification described here may be used for the isolation of γδТ cells for in vivo and in vitro studies.
4. CONCLUSION
The method of γδТ cell isolation from mouse small intestine without collagenase treatment and without centrifugation through Percoll gradients is described. The procedure permits obtaining viable γδТ cells of 90% purity.
REFERENCES
- Yoshikai, Y. (1999) The interaction of intestinal epithelial cells and intraepithelial lymphocytes in host defense. Immunologic Research, 20, 219-235. http://dx.doi.org/10.1007/BF02790405
- Hänninen, A. and Harrison, L.C. (2000) Gamma delta T cells as mediators of mucosal tolerance: The autoimmune diabetes model. Immunologic Research, 173, 109-119. http://dx.doi.org/10.1034/j.1600-065X.2000.917303.x
- Yang, Y., Tung, J.W., Ghosn, E.E., Herzenberg, L.A. and Herzenberg, L.A. (2007) Division and differentiation of natural antibody-producing cells in mouse spleen. Proceedings of the National Academy of Sciences of the United States of America, 104, 4542-4546. http://dx.doi.org/10.1073/pnas.0700001104
- Kühl, A.A., Pawlowski, N.N., Grollich, K., Blessenohl, M., Westermann, J., Zeitz, M., Loddenkemper, C. and Hoffmann, J.C. (2009) Human peripheral gamma/delta T cells possess regulatory potential. Immunology, 128, 580- 588. http://dx.doi.org/10.1111/j.1365-2567.2009.03162.x
- Kroese, F.G., Butcher, E. S., Stall, A.M., Lalor, P.A., Adams, S. and Herzenberg L.A. (1989) Many of the IgA producing plasma cells in the murine gut are derived from self-replenishing precurcors in the peritoneal cavity. International Immunology, 1, 75-84. http://dx.doi.org/10.1093/intimm/1.1.75
- Bhagat G., Naiyer, A.J., Shah, J.G., Harper, J., Jabri B., Wang, T.C., Green, P.H. and Manavalan, J.S. (2008) Small intestinal CD8+TCRgammadelta+NKG2A+ intraepithelial lymphocytes have attributes of regulatory cells in patients with celiac disease. The Journal of Clinical Investigation, 118, 281-293. http://dx.doi.org/10.1172/JCI30989
- James, S.P. and Graeff, A.S. (1985) Spontaneous and lymphokine-induced cytotoxic activity of monkey intestinal mucosal lymphocytes. Cellular Immunology, 93, 387-397.http://dx.doi.org/10.1016/0008-8749(85)90143-1
- Lyscom, N. and Brueton, M.J. (1982) Intraepithelial, lamina propria and Peyer’s patch lymphocytes of the rat small intestine: Isolation and characterization in terms of immunoglobulin markers and receptors for monoclonal antibodies. Immunology, 45, 775-783. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1555414/
- Rothkötter, H.J., Kirchhoff, T. and Pabst, R. (1998) Lymphoid and non-lymphoid cells in the epithelium and lamina propria of intestinal mucosa of pigs. Gut, 35, 1582-1589. http://dx.doi.org/10.1136/gut.35.11.1582
- Mosley, R.L. and Klein, J.R. (1992) A rapid method for isolating murine intestine intraepithelial lymphocytes with high yield and purity. Journal of Immunological Method, 156, 19-26. http://www.ncbi.nlm.nih.gov/pubmed/1331242 http://dx.doi.org/10.1016/0022-1759(92)90006-F
- Montuflar-Solis, D. and Klein, J.R. (2006) An improved method for isolating intraepithelial lymphocytes (IELs) from the murine small intestine with consistently high purity. Journal of Immunological Method, 308, 251-254. http://dx.doi.org/10.1016/j.jim.2005.10.008
- Ye, Y.F., Yue, M., Xi, J., Chen, S.H. and Li, Y.M. (2010) Isolation of Murine Small Intestinal Intraepithelial gdT. Immunological Investigations, 39, 661-673. http://dx.doi.org/10.3109/08820131003753026