Open Journal of Animal Sciences
Vol.07 No.03(2017), Article ID:77824,29 pages
10.4236/ojas.2017.73025
Leishmania in Marsupials―An Overview of Infection Records in the Americas and Australia
João Carlos Araujo Carreira1*, Mônica de Avelar Figueiredo Mafra Magalhães2, Reginaldo Peçanha Brazil3, Alba Valéria Machado da Silva1
1INERU/IOC, FIOCRUZ, Rio de Janeiro, Brazil
2ICICT, FIOCRUZ, Rio de Janeiro, Brazil
3IOC, FIOCRUZ, Rio de Janeiro, Brazil

Copyright © 2017 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: February 28, 2017; Accepted: July 18, 2017; Published: July 21, 2017
ABSTRACT
Marsupials have been the subjects of studies of both experimental and natural infections with different species of Leishmania in the Americas as well as Australia. Over a century has passed since the first description of a mammal being infected with Leishmania and since then several reviews have been published on the systematics of the parasites as well as their hosts. Consequently, an update of this information is needed in order to assure correct identification of the species involved in each case. A comprehensive review was undertaken and included most of the records of marsupials being infected with Leishmania sp. Emphasis was placed on parasitological, eco epidemiological and taxonomic information of both the parasites and mammalian hosts. The clinical profiles of the infections varied from severe, which principally occurred in experimentally infected animals, to asymptomatic, which was typical of natural infections. Marsupials must be considered one of the most important groups of mammalian hosts of Leishmania sp. Important factors, such as their ancient origin, wide geographical distribution and susceptibility to infections by different species of Leishmania, but with considerable resistance to the disease, could indicate that didelphids play a key role in the evolution of Leishmania in the New World.
Keywords:
Leishmania, Marsupials, Wild Hosts, Americas, Australia

1. Introduction
Leishmaniasis is a vector-borne disease caused strictly by intracellular parasitic protozoa of the genus Leishmania. Its geographic distribution covers approximately 88 countries where almost 350 million people live. The majority of these countries are in the tropics and subtropics, and include the USA, the rain forests of Central and South America, Australia and the deserts of western Asia.
The insect vectors of Leishmania comprise a dozen species of sand flies belonging two genera: Phlebotomus in the Old World and Lutzomyia in the New World.
Mammalian hosts of Leishmania are very diverse in comparison to the vectors, with nearly 100 species from more than 10 orders. This diversity plays a key role in the maintenance of these parasites in nature, and is one of the most important factors in determining their dispersion and diversity.
Among the New World mammalian groups that serve as reservoirs of Leishmania spp., marsupials must be considered one of the most important. This infraclass occurs predominantly in the New World and Australasia and represents a monophyletic group derived from the last common ancestor of extant metatherians.
Marsupials are characterized by giving birth to relatively immature litters that, in most cases, complete their development in the mother’s pouch. Approximately 70% of the extant species of marsupials occur in Australia, New Guinea, and adjacent islands, with the remaining diversity occurring in the New World, mainly in South America but also in Central and North America well north of Mexico [1] .
Marsupials in the Americas belong to the Superorder Ameridelphia, which is composed of two families: the Didelphidae (opossums) and the Caenolestidae (shrew opossums). Marsupials in Australasia belong to the Superorder Australidelphia, which contains 17 extant families: Microbiotheriidae, Dasyuridae, Myrmecobiidae, Thylacomyidae, Peramelidae, Notoryctidae, Phascolarctidae, Vombatidae, Phalangeridae, Burramyidae, Tarsipedidae, Petauridae, Pseudocheiridae, Potoroidae, Acrobatidae, Hypsiprymnodontidae and Macropodidae [1] .
The ecology of marsupials is substantially diverse, and is indicative of their remarkable diversification in Australia and the Americas. Marsupials inhabit virtually all of the same niches that placental mammals do, and occur in every major terrestrial habitat. As a group, marsupials also possess diverse diets including carnivory, herbivory and omnivory, and species capable of feeding on a wide range of food sources [2] .
In spite of initially being considered only as secondary reservoir of Leishmania spp., marsupials have come to be accepted as playing a very important role in the epidemiology of leishmaniasis by acting as sylvatic and/or synanthropic hosts. There have been several reports describing natural and experimental infections of marsupials by different species of Leishmania belonging both subgenera L. (Leishmania) and L. (Viannia).
Over a century has passed since the discovery of Leishmania spp. and several revisions have been published on the systematics of the parasite as well as of their hosts. Therefore, periodic updates are very important for assuring correct identification of the species involved.
The aim of the present review is to provide detailed information about the species of marsupials that have documented cases of their infection, naturally or experimentally, with Leishmania spp. Aspects of the biology, systematics and parasites of these species are also addressed.
1.1. Marsupial Hosts of Leishmania in the Americas
As we are going to show, all of the studies involving infections of marsupials with Leishmania in the New World have been with species of the family Didelphidae. These cases included 8 out of 19 extant didelphid genera: Didelphis, Marmosa, Metachirus, Caluromys, Phylander, Monodelphis, Marmosops and Gracilinanus (Figure 1).
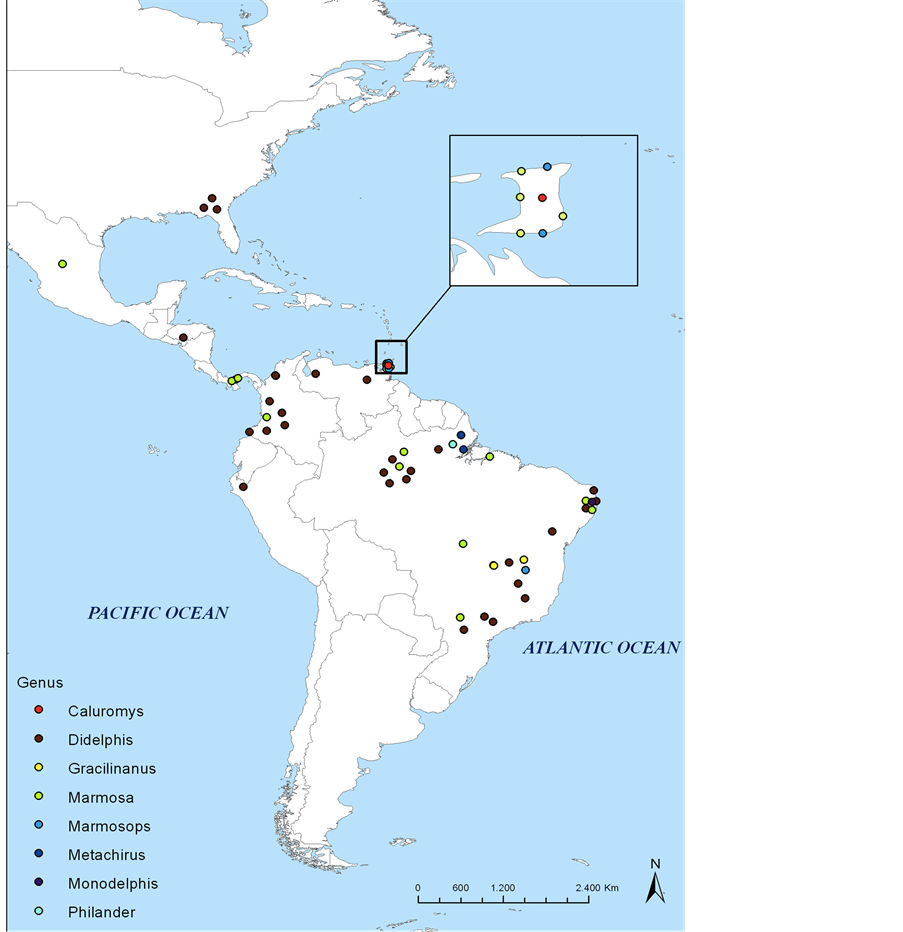
Figure 1. Geographical distribution of records of infection with Leishmania spp. in different didelphid genera in the Americas from 1962 to 2015. All data were mapped through ArchView software using geographical coordinates provided by the authors or estimated based on reference points provided in the studies and utilizing ArchView and Google Earth.
1.2. Didelphidae
The family Didelphidae is certainly one of the main groups of mammals in the life cycle of Leishmania because of the importance of many of its species as reservoir hosts. These species are among the best-studied mammalian hosts of Leishmania with a great number of records of infections. Furthermore, Didelphis is the only genus of New World marsupials that have already been experimentally infected with the parasite.
After Didelphis, the genus Marmosa must be considered second in importance with 18 records of infections [15 in M. (Marmosa) and 3 in M. (Micoreus)]; followed by Metachirus, Gracilinanus and Caluromys, with 2 records each, and Phylander, Monodelphis and Marmosops, with one each.
2. The Genus Didelphis (Opossums)
Opossums possess several important features that facilitate their role as reservoir hosts of Leishmania spp., including an ancient origin; a wide geographic distribution, occurring from the USA to Argentina; and inhabiting various habitats where Leishmania are endemic (Figure 1). Unlike other sylvatic mammalian hosts, species of Didelphis characteristically adapt easily to anthropic habitats, being able to take part in both sylvatic and peri-domestic parasite cycles, and can even serve as a link between them.
Initially the genus Didelphis contained three species [3] : D. virginiana occurring in North and Central America; Didelphis marsupialis living predominantly in South America, but sympatric with the former in Central America; and D. albiventris described from higher altitudes in South America. Subsequently, three new species were recognized, D. aurita, D. pernigra and D. imperfect [3] [4] [5] , bringing the total number of species of Didelphis to six.
2.1. Didelphis and Leishmania
Among the six extant species of the genus Didelphis, infection with Leishmania has been previously documented for four: D. marsupialis, D. albiventris, D. aurita and D. virginiana. In the present study, we report for the first time the infection of D. pernigra in Peru. This species was previously considered as D. albiventris, but was recognized as a distinct species with the revision of the genus [6] .
2.2. Didelphis in Brazil
2.2.1. D. marsupialis (Common Opossum)
All records of Leishmania spp. infection of D. marsupialis in Brazil are natural infections, and most were in the states of Amazonas and Para in the Amazon Region (Table 1).
In Amazonas, Arias and Naiff [7] [8] emphasized the importance of D. marsupialis as a reservoir host of the L. guyanensis in disturbed primary forest because of the high levels of infection they observed, varying from 20% to 71%. 4% depending on the area studied. The same studies also found considerable infection

Table 1. List of records of marsupials infected with Leishmania spp. in Brazil. Data provided include the states where the studies were carried out, the mammalian host species, the parasite species, the type of infection and the diagnostic methods and references.
Natural infection (N), Histology (H), Culture (C), Imprint (I), Molecular Biology (MB), Xenodiagnosis (X), Isoenzyme (Iso), Indirect Immunofluorescence Assay (IFA), Dipstick Test (K-39), Direct Agglutination Test (DAT), Smear (S), Monoclonal Antibody (MA), Enzyme-linked Immunosorbent Assay (ELISA), not presented (NP), (a) Originally referred to by the authors as Micoureus demerarae.
of D. marsupialis with L. amazonensis, ranging from 4.7% to 25%.
In the state of Para, Lainson et al. [9] observed a lower level of infection than reported in Amazonas, on the order of 4.7% for both parasite species.
In addition to the records of Leishmania infection of D. marsupialis in the Amazon Region, there is one report of a prevalence of 21.6% among the D. marsupialis in the state of Minas Gerais [10] . However, it is quite probable that the species involved was D. aurita and not D. marsupialis, because the latter does not occur in the state of Minas Gerais.
2.2.2. D. albiventris (White-Eared Opossum)
Didelphis albiventris must be considered the second most important reserve host species of the genus Didelphis, with eleven records of infection with L. infantum, L. braziliensis and L. amazonensis. Ten of these cases were in Brazil where this species is the most cited in the genus, including the state of Bahia with L. infantum, L. braziliensis and L. amazonensis; the state of Pernambuco with L. braziliensis and L. (Viannia); the state of Rio Grande do Norte with L. infantum; the state of Minas Gerais with L. braziliensis; the state of São Paulo with Leishmania spp.; and the federal district of Brasilia with Leishmania spp (Table 1).
In the state of Bahia there have been three studies. The first [11] described a case of natural infection of one opossum with L. infantum in the city of Jacobina. This report was highlighted as the first record for the continent of a non-canid wild animal being naturally infected with the agent of visceral leishmaniasis. The same study also demonstrated that D. albiventris was the most abundant wild mammal around the houses in the area and that Lutzomyia longipalpis, the principal vector of visceral leishmaniasis in the New World, was observed promptly feeding on the animals.
In the second study in Bahia, also in Jacobina, Sherlock et al. [12] investigated commensal and sylvatic reservoirs of Leishmania spp. Didelphis albiventris was the most frequently caught mammal with 44% of the total, among which were also a bat, seven different rodent species, armadillos and canids. Of the 84 individuals of D. albiventris studied, two (2.4%) were infected with L. infantum, one (1.2%) with L. amazonensis and one (1.2%) with L. braziliensis. The authors reasoned that the low rate of infection indicates that D. albiventris should not be considered the primary and most important reservoir of visceral leishmaniasis in Jacobina. However, it is important to point out that despite the apparent low prevalence, other important epidemiological indicators, such as predominance in domestic and peri-domestic habitats, attractiveness to the insect vector and concomitancy with human cases of the disease, could counter such low percentages of infection.
The third study in Bahia is probably related to the two previously mentioned studies. In this case, Sherlock [13] described the natural infection of D. albiventris with L. infantum, however, it is not clear which species was actually studied because it was referred to as the “black ear opossum Didelphis albiventris”, but this species has characteristic white ears.
In the state of Pernambuco there have been two studies of infection of D. albiventris. In the first, Brandão-Filho et al. [14] found (5\37) 13.5% of D. albiventris to be infected with L. (Viannia), with 13.3% in the forest and 3.1% in plantations. They suggested that besides rodents, opossums were among the animals most likely to function as reservoirs for L. (V.) braziliensis in Northeastern Brazil.
The second study was carried out by Lima et al. [15] in the municipality of São Vicente Férrer. They found rates of infection of D. albiventris to be 37.5% for L. (Viannia) and 6.3% for L. infantum, with one animal being co-infected by both species.
In the state of Rio Grande do Norte there has been only one record of an individual D. albiventris being naturally infected with L. infantum. The study surveyed small wild mammals from a Biological Reserve located in the municipality of Natal and domestic animals from an urban area [16] . The results showed that of 20 marsupials, including 16 Didelphis albiventris and 4 Monodelphis domestica, only one D. albiventris (5%) tested serologically positive for L. infantum.
In the state of São Paulo there have been two records of infection of D. albiventris with Leishmania spp. The first was from a study carried out by Quintal et al. [17] in the extreme western part of the state in a remnant of Atlantic Forest. They found that 1.6% of D. albiventris tested positive to qPCR. The second case was from a study in the city of Bauru in the central region of the state where Santiago et al. [18] observed a high rate of infection among opossums (91.6%), with 112 being D. albiventris and 10 being D. aurita. However, these species are sympatric in the area and the percentage was attributed to both of them together.
The last record of infection of D. albiventris with Leishmania in Brazil occurred in an endemic area for American tegumentary leishmaniasis, located in the northern part of the state of Minas Gerais. Here Quaresma et al. [19] studied domestic and synanthropic wild hosts of L. braziliensis in an Indigenous Territory. Opossums were the second most frequent animal captured representing 19.6% of the total, 28.6% of which were infected with L. braziliensis. The rate of infection of the peri-domicile areas and the trails between the houses were comparable with 20% and 21.4%, respectively.
2.2.3. D. aurita (Black-Eared Opossum)
All the reports of Leishmania spp. infection of D. aurita occurred in the Brazilian states of São Paulo and Rio de Janeiro. The parasite species involved were L. mexicana, L. forattinii and L. infantum (Table 1).
In the state of São Paulo there have been four publications relating to the infection of D. aurita by Leishmania spp, L. mexicana and L. forattinii. The first was Yoshida et al. [20] , who in a research note reported the first case of natural infection of a marsupial with Leishmania in the state. The authors considered the probable importance of marsupials as hosts of Leishmania, in addition to rodents. In the second paper, Yoshida et al. [21] characterized the Leishmania species that was isolated from the D. aurita of their previous note as Leishmania mexicana, and in the third paper, Yoshida et al. [22] described Leishmania (L.) foratinii as a new parasite species infecting opossums and rodents captured in primary Atlantic Forest. The fourth record for São Paulo was mentioned above under the account for D. albiventris, where Santiago et al. [18] observed that 91.6% of the marsupials they tested were naturally infected; however, this was attributed to the two species D. albiventris and D. aurita, combined.
In the state of Rio de Janeiro, there has been one report of infection of D. aurita with Leishmania. Carreira et al. [23] observed a prevalence of 16.6% among D. aurita in a single endemic area for visceral leishmaniasis in Atlantic Forest. Macroscopic examination revealed that one opossum exhibited symptoms compatible with spleen hypertrophy whereas three others possessed amastigotes in the spleen. In addition, one of the opossums possessed parasites in a submandibular lymph node.
2.3. Didelphis in Colombia
D. marsupialis
There have been six reports of Leishmania spp. infection of D. marsupialis in Colombia. One of these reports was an experimental infection with L. infantum, whereas the others were natural infections with the following parasite species: L. infantum (three), L. braziliensis (one) and L. (Viannia) (one) (Table 2).
Table 2. List of records of marsupials infected with Leishmania spp. in South America other than from Brazil. Data provided include countries where the studies were carried out, the mammalian host species, the parasite species, the type of infection, the diagnostic methods and references.
Natural infection (N), Experimental infection (E), Histology (H), Culture (C), Imprint (I), Molecular Biology (MB), Xenodiagnosis (X), Isoenzyme (Iso), Smear (S), not presented (NP), (a) Originally referred to by the author as Marmosa fuscata, (b) Originally referred to by the author as Marmosa mitis, (c) Originally referred to by the authors as Didelphis albiventris.
In the case of the experimental infection, Travi et al. [24] inoculated an adult female and four recently weaned young opossums with amastigotes or promastigotes. The parasite burden was estimated by culture, testing for parasite DNA in skin samples, histopathological analysis and xenodiagnoses. The authors utilized two routes of inoculation, and observed the same effectiveness when inoculated through the dermis as when inoculated with a large number of parasites directly into the blood stream. Most animals presented subclinical infection, with a discrete inflammation of the liver, lymph nodes and spleen. An exception was one animal that showed evident signs of disease, including abundant cell parasitism, general spleen necrosis and fatty degeneration in the liver. The xenodiagnoses found sand flies to become infected even after feeding on subclinically infected opossums, which reinforces their potential as reservoirs, as had previously been observed in dogs by Molina et al. [25] .
In conclusion, these authors highlighted the role of D. marsupialis as a wild reservoir host of L. infantum in Colombia, and explained that it was a function of several factors: 1) among all the animals trapped in an endemic area (data not shown), D. marsupialis was the most attractive to Lutzomyia evansi, the principal vector species of visceral leishmaniasis in Colombia; 2) it presented significant rates of natural infection [26] [27] ; and 3) this species represented, in terms of biomass, the most abundant small mammal in a focus of visceral leishmaniasis in northern Colombia [28] .
Among the three records of natural infections of D. marsupialis with L. infantum in Colombia, Corredor et al. [29] studied a population in El Callejon, a rural area highly endemic for L. infantum and where originally existed a tropical dry forest. The authors stressed the importance of D. marsupialis in the domestic cycle of the parasite because of the high percentage (around 32%) of infection observed.
Another study took place in the municipality of San Andreas de Sotavent in northwestern Colombia inside of an ecological zone composed of tropical dry forest. Here Travi et al. [27] recorded a prevalence of infection of 22.7% and they pointed out the probable role of D. marsupialis in both sylvatic and domestic cycles of the parasite.
In the third report, Travi et al. [30] studied the dynamics of L. infantum infection in small mammals of undisturbed and degraded tropical dry forests in northern Colombia. Among the mammals, D. marsupialis had one of the highest prevalences of infection with 14.3% and 9.5% in the undisturbed and degraded forests, respectively. The authors concluded that although the infection rate was similar at both sites, its significant increase in relative abundance in the degraded forest facilitated the establishment of the parasite in this habitat.
In the record describing the natural infection of D. marsupialis with L. braziliensis, Alexander et al. [31] utilized PCR and DNA hybridization to detect the potential presence of the parasite in tissue samples from wild mammals trapped in coffee plantations at La Guaira and Jinguales, Colombia. Among the five opossums tested, only one presented a very faint positive result with a specific probe for L. braziliensis.
In the last record associating D. marsupialis with Leishmania infections in Colombia, Ocampo et al. [32] studied environmental factors associated with American cutaneous leishmaniasis in an Andean focus and in Agua Bonita. They found 33.3% of the animals to be infected with Leishmania (Viannia), as detected by PCR-Southern blot, and the importance of opossums as a reservoir host of cutaneous leishmaniasis in disturbed habitats was reinforced.
2.4. Didelphis in Venezuela
D. marsupialis
There have been just two studies that have recorded the infection of D. marsupialis with Leishmania spp. The first comprised both experimental and natural infections, and described D. marsupialis as a primary peri-urban reservoir of Leishmania spp in the city of Trujillo, an endemic area of tegumentary leishmaniasis. In the experimental infections, 50% of the animals became infected after inoculation with L. braziliensis, and through the xenodiagnoses with Lutzomyia townsendi, an anthropophilic sandfly species very common in the area, the percentage of infection was 5.5% [33] . In the natural infections the parasite species observed were L. braziliensis and L. garnhami, both with the same prevalence of 2.5%.
In the second study in Venezuela, Zulueta et al. [34] studied aspects of the epidemiology of visceral leishmaniasis in an endemic focus in eastern Venezuela. They observed that one out 14 D. marsupialis was infected (7.1%) with L. donovani species complex based on PCR (currently L. infantum).
2.5. Didelphis in Honduras
D. marsupialis
There have been two reports describing experimental infections of D. marsupialis with L. infantum in Honduras, which represent the first work to utilize marsupials in studies of experimental infection with Leishmania, in the New World (Table 3).
In the first case, Lainson and Strangways-Dixon [35] reported that one out four (25%) of the D. marsupialis infected through intraperitoneal and intradermal inoculations developed visceral infection. In 1964, the same authors, in a
Table 3. List of records of marsupials infected with Leishmania spp. in Central America Data provided include the countries where the studies were carried out, the mammalian host species, the parasite species, the type of infection, the diagnostic methods and references.
Natural infection (N), Experimental infection (E), Histology (H), Culture (C), Imprint (I), Smear (S), Not determined (ND).
second article [36] , reported that among a dozen mammal species inoculated, detectable infections were produced only in two: D. marsupialis (25%) and a species of rodent, Sigmodon hispidus (100%). Despite the high prevalence observed also among the opossums in this study, the authors only highlighted the rodents as important hosts.
2.6. Didelphis in Peru
D. pernigra (Formerly D. albiventris)
The only supposed record of Leishmania infections in D. albiventris outside of Brazil occurred in Peru, where domestic and wild mammals were found naturally infected with L. (V.) peruviana in the Peruvian Andes (Table 2). Opossums exhibited the highest prevalence (2.8%) among the wild animals, even though a variety of rodent genera that have been traditionally considered as wild reservoirs, such as Phyllotis, Akodon and Oryzomys were included [37] .
It is important to point out that although the authors described correctly the opossum species as D. albiventris, three years later a revision of this genus recognized the Andean White-eared Opossum, D. pernigra, as a new species.
2.7. Didelphis in the USA
2.7.1. D. marsupialis
There is only one report concerning Leishmania sp. infection of D. marsupialis in the USA. Hanson et al. [38] inoculated individuals of D. marsupialis intracardially with an African strain of Leishmania donovani, and reported that those animals developed fulminating visceral leishmaniasis and concluded that they would be useful in the study of anti-leishmanial drugs (Table 4).
2.7.2. D. virginiana (North American Opossum)
There are two records of experimental infections of D. virginiana with an L. donovani African strain (Khartoum) (Table 4). White et al. [39] compared the patterns of infection with L. donovani among opossums, armadillos and ferrets and concluded that the opossum was the most susceptible because of the larger number of parasites in both liver and spleen. These same authors [40] evaluated the effect of two chemotherapeutic drugs on 17 individuals of D. virginiana that had been previously inoculated with L. donovani. They concluded that factors
Table 4. List of records of marsupials infected with Leishmania spp. in North America. The data indicate the countries where the studies were carried out, both the host and parasite species, the type of infection, the diagnostic methods and references.
Natural infection (N), Experimental infection (E), Histology (H), Culture (C), Imprint (I), Molecular Biology (MB), not presented (NP).
related to the pattern of infection, such as rapid acute death and abnormalities of hemostasis, could limit the use of this species as an animal model for testing anti-leishmanial drugs.
3. The Genus Marmosa and Other Mouse Opossums
Before addressing Leishmania infection of mouse opossums, it is necessary to clarify that these animals comprise a great number of different taxonomic groups with numerous morphological similarities and broad concurrence in their geographical distribution (Figure 1).
Formerly, the term “mouse opossum” was mostly associated with the genus Marmosa, nevertheless this genus has been the subject of multiple revisions and several new genera have come to be considered mouse opossums as well [41] .
In 1933, in one of the first revisions of the genus Marmosa, Tate [42] subdivided the genus into five groups, Cinerea, Murina, Noctivaga, Microtarsus and Elegans. Each of these groups was then subdivided into sections varying in number from two to four. For example, the Cinerea group contained two sections, Marmosa Cinerea and M. Regina, while the Murina group had four sections: M. Murina, M. Mitis, M. Mexicana and M. Canesces.
Later, Gardner and Creighton [43] , re-described the groups and sections proposed by Tates, assigning the sections Cinerea, Murina, Noctivaga and Elegans, to the respective genera Micoureus [1] (Lesson, 1842), Marmosa (Gray, 1821), Marmosops (Matschie, 1916) and Thylamys (Gray, 1843). In addition, most of the species belonging the Microtarsus group were placed in a new genus, Gracilinanus.
More recently, phylogenetic research based on molecular data [44] [45] [46] determined that Marmosa was polyphyletic, while Marmosops, Micoureus, and Thylamys, as previously established by Gardner and Creighton [43] , were all monophyletic. A new genus, however, was defined for “Marmosa” canescens by Voss and Jansa [47] , and some new genera were later described for two clades previously suppressed by synonymy within Gracilinanus [48] [49] . Finally, after the last revision of genus Marmosa, Micoureus, which was previously considered a separate genus, was moved into Marmosa as a subgenus [50] .
Currently, the term “mouse opossum” can be associated with five genera, as follows: Marmosa (Linnaeus’s Mouse Opossum), Marmosops (Slender Mouse Opossums), Thylamys (Fat-tailed Mouse Opossums), Tlacuatzin (Grayish Mouse Opossum) and Gracilinanus (Gracile Mouse Opossum). Nonetheless, in some instances, both Gracilinanus and Marmosops have been excluded from consideration as “mouse opossums” and instead are identified as just the Gracile Opossum and the Slender Opossum.
The genus Marmosa specifically comprises a collection of black-masked and long-tailed pouchless marsupials. They are solitary, nocturnal and live in a wide range of tropical and subtropical habitats from Mexico to Bolivia. It has been considered the most abundant genus of the family Didelphidae, being composed by 19 currently recognized species belonging to five subgenera as follows: 1) M. Marmosa (Gray, 1821) represented by the species macrotarsus, murina, tyleriana, and waterhousei; 2) M. Micoureus (Lesson, 1842) with the species alstoni, constantiae, demerarae, paraguayanus, phaea and regina; 3) M. Stegomarmosa (Pine, 1972) with andersoni and lepida; 4) M. Eomarmosa [51] with rubra; and 5) M. Exulomarmosa [51] with the species isthmica, mexicana, robinsoni, simonsi, xerophila, and zeledoni.
3.1. Marmosa and Leishmania
Five species of Marmosa have already been reported to have been naturally infected by different species of Leishmania: 1) M. (Exulomarmosa) robinsoni, infected with Leishmania spp and L. mexicana; 2) M. (Marmosa) murina, with Leishmania spp and L. amazonensis; 3) M. (Micoreus) demerarae; and 4) M. (Micoreus) paraguayanus both infected with L. braziliensis and L. amazonensis; and 5) M. (Exulomarmosa) mexicana with L. mexicana.
3.2. Marmosa in Brazil
3.2.1. Marmosa (M.) murina (Murine Mouse Opossum)
All studies related to infections of Marmosa murina with Leishmania sp. occurred in central and northern Brazil (Table 1). The three papers from the central region include surveys performed in the state of Mato Grosso. Lainson and Shaw [52] found 7.7% of M. murina to be naturally infected with an unidentified species of cutaneous Leishmania sp., however, their criterion for selecting animals for testing was simply the presence of skin lesions. In a subsequent paper, the same authors re-described and expanded on the information related to their first report [53] , and in a third paper, with additional coauthors, they isolated a stock of Leishmania from M. murina, that they characterized as L. amazonensis through enzymatic profile [54] . In the Northern Region of Brazil, there has been only one record of an individual M. murina being infected with an unspecified strain of Leishmania [52] .
3.2.2. Marmosa (M.) demerarae (Long-Furred Woolly Mouse Opossum)
In the only record of infection of M. demerarae with Leishmania sp. in Brazil, Arias et al. [8] observed 100% (2/2) infection with L. amazonensis in an area of virgin forest, but with considerable human activities, in the state of Amazonas (Table 1).
3.2.3. Marmosa (M.) paraguayanus (Tate’s Woolly Mouse Opossum)
The two records of Leishmania spp. infection of M. paraguayanus in Brazil were presented by Quintal et al. [17] (Table 1). These researchers studied six areas surrounding the Atlantic Forest in the extreme western part of the state of São Paulo, and found a prevalence of 11.6% for Leishmania spp by qPCR. In addition, from three tissue samples taken for parasite species identification, two were positive for L. amazonensis and one for L. braziliensis. Given the high parasitic load they found in the skin samples, these authors acknowledged the probable role of this marsupial as a potential reservoir host of Leishmania spp.
3.2.4. Marmosa spp
Concluding the accounts involving the genus Marmosa in Brazil, there are two more studies in which the authors did not identify the species; however, given that the studies were performed in the state of Pernambuco, and considering the geographic range of potential species, it is likely these were individuals of either M. murina or M. demerarae.
In the first paper, Brandão-Filho et al. [14] , studied wild and synanthropic hosts in an endemic area of cutaneous leishmaniasis (Table 1). With the use of PCR, they observed infections with L. braziliensis that varied in prevalence from 16.7% to 25%, depending on the year when the samples were collected. Additionally, prevalence varied according to location of capture, with the rates being 4.1%, 1.6% and 1.2% for animals caught in forest, animal sheds and plantations, respectively.
The second record was presented by Lima et al. [15] , who studied small mammals as hosts of Leishmania spp in a highly endemic area for zoonotic leishmaniasis. Twenty five percent (1/4) of Marmosa spp tested positive for Leishmania spp through PCR and none of the animals presented clinical signs suggestive of leishmaniasis such as skin lesions (Table 1).
3.3. Marmosa in Trinidad and Tobago
Marmosa (E.) robinsoni (Robinson’s Mouse Opossum)
Two manuscripts addressed natural infections of Marmosa mitis with Leishmania spp. in Trinidad and Tobago, although currently this species is considered a synonym of M. robinsoni, and so this last name must be considered.
The first manuscript was a preliminary report in which the authors report the isolation (through inoculation of a hamster) of a parasite morphologically corresponding to Leishmania from one specimen of Marmosa mitis [55] (Table 2). The second study, entitled “Enzootic rodent leishmaniasis in Trinidad, West Indies” was carried out by the same author [56] in six areas covering the northern part of the country, where they found 16.7% of Marmosa mitis to be naturally infected with Leishmania spp. Nevertheless, these results probably underestimated the real prevalence of infection because only animals that presented skin lesions were included in the study.
3.4. Marmosa in Panama
Marmosa robinsoni
In Panama, there have been three studies recording infections of M. robinsoni with Leishmania spp., two with Leishmania spp and one with L. mexicana (Table 3). In the first, Herrer et al. [57] described a focus of enzootic cutaneous leishmaniasis in a forest area in the eastern part of the country where they recorded a prevalence of 2.8% infection with a Leishmania sp., species they described as to behave like L. mexicana in experimentally infected hamsters. Telford et al. [58] studied the ecological factors relating to the mammalian hosts in the same area, but in this case trapping effort was concentrated in a secondary forest, and consequently the prevalence was 1.9% and M. robinsoni were considered incidental hosts. Herrer et al. [59] studied the reservoir hosts of cutaneous leishmaniasis in a forest habitat and observed that 1.6% of M. robinsoni were infected with L. mexicana.
3.5. Marmosa in Colombia
Marmosa demerarae
In Colombia there is one record where Alexander et al. [31] observed an infection prevalence of 66.7% (two out of three) for L. braziliensis among M. demerarae caught in coffee plantations in the western part of the country between the Andes and the coast (Table 2).
3.6. Marmosa in Mexico
M. (Marmosa) mexicana (Mexican Mouse Opossum)
In Mexico, Van Wynsberghe et al. [60] observed that in addition to high rates of infection (50% and 100% during 1997-1998 and 2001-2004, respectively), most animals were asymptomatic in certain areas of an important focus of cutaneous leishmaniasis in the state of Campeche (Table 4).
4. The Genus Marmosops (Slender Opossums)
As originally described, the genus Marmosops [61] was comprised of 11 species, with M. incanus [62] being assigned as the type species. Later, Tate [42] treated Marmosops as a junior synonym of Marmosa, but Gardner and Creighton [43] recognized Marmosops as a distinct taxon, and elevated all of Tate’s species groups of Marmosa to generic rank.
Currently, the genus Marmosops contains the following 14 species: M. bishopi, M. cracens, M. dorothea, M. fuscatus, M. handleyi, M. impavidus, M. incanus, M. invictus, M. juninensis, M. neblina, M. noctivagus, M. parvidens, M. paulensis and M. pinheiroi. However, some authors consider M. creightoni to be an additional species.
4.1. Marmosops and Leishmania
There are three records that refer to species of Marmosops being naturally infected with Leishmania spp., one in Brazil and two in Trinidad. The species involved were M. incanus and M. fuscatus, and were infected with L. guyanensis and Leishmania spp, respectively (Figure 1).
4.2. Marmosops in Brazil
Marmosops incanus (Grey Slender Opossum)
Quaresma et al. [19] observed a prevalence of 50% (1/2) infection with L. guyanensis among M. incanus in one area in the state of Minas Gerais State where other animals, including rodents and canids, were infected with other Leishmania species, including L. braziliensis and L. infantum (Table 1).
4.3. Marmosops in Trinidad
In Trinidad, the opossum species cited by the authors as Marmosa fuscata, is now recognized as a new genus and species. Therefore Marmosa fuscata is currently considered as a synonym of Marmosops fuscatus.
Marmosops fuscatus (Dusky Slender Opossum)
The first record of a M. fuscatus being infected with with Leishmania in Trinidad and Tobago was in 1969 as part of a preliminary report about natural infection with Leishmania sp [55] (Table 2). Subsequently, in 1974, the same author published a study entitled “Enzootic Rodent Leishmaniasis in Trinidad, West Indies”, in which an overall infection prevalence of 11.1% was reported; although a considerably higher prevalence of infection was reported for a single area of Evergreen Seasonal Marsh Forest (33.3%), these animals were considered as only secondary hosts.
5. The Genus Metachirus (Brown Four-Eyed Opossum) and Leishmania
The genus Metachirus contains only a single currently recognized species, Metachirus nudicaudatus, which is also the largest of the pouchless didelphids found in forest habitats. In spite of the recognition of just one species, five subspecies have been identified and it has been suggested that one or more of these subspecies may in fact be genuine species [52] . There are only two records of M. nudicaudatus being naturally infected with Leishmania in Brazil (Figure 1).
In the first case, Miles et al. [54] used enzymatic characterization to identify a strain of Leishmania isolated from M. nudicaudatus as L. amazonensis (Table 1). In the second case, Lainson et al. [9] observed considerable prevalence (66.7%) of L. amazonensis infections in an area highly endemic for cutaneous leishmaniasis (Table 1).
6. The Genus Caluromys (Woolly Opossum) and Leishmania
Currently the genus Caluromys is composed of three species: C. philander, C. derbianus and C. lanatus. Of these, the firsts two have been reported with natural infections of Leishmania sp. in Trinidad and Panama (Figure 1).
6.1. Caluromys philander (Bare-Tailled Woolly Opossum) in Trinidad
In Trinidad, Tikasingh [56] observed a total prevalence of 4.3% for cutaneous leishmaniasis in six areas of a variety of different habitats in the northern part of the country (Table 2). This study was the first to record the presence of amastigotes in a sore on an individual of this species.
6.2. C. derbianus (Central American Woolly Opossums) in Panama
In Panama, Johnson and Hertig [63] studied the behavior of Leishmania in Panamanian sandfly species fed on naturally infected mammals.
In phlebotomines that had previously fed on one Caluromys (probably C. derbianus), they observed a number of flagellates similar to some already observed in cultures of Leishmania isolated from sandflies from the same area. However, it was not clear which parasite species were involved in this case because, the authors stated, trypanosomes are frequently correlated with infections in Caluromys (Table 3).
An interesting observation was of sandflies feeding on young animals that were still in the marsupium.
7. The Genus Gracilinanus (Gracile Mouse Opossums) and Leishmania
Gracilinanus was separated from the genus Marmosa in 1989 and currently six species are recognized: G. aceramarcae, G. agilis, G. dryas, G. emiliae, G. marica and G. microtarsus.
There are two records of species of Gracilinanus being infected with Leishmania spp., both in Brazil and both with the species G. agilis (Agile Gracile Mouse Opossum) (Figure 1).
The first report was in the state of Minas Gerais in southeastern Brazil where Quaresma et al. [19] studied wild, synanthropic and domestic hosts in an endemic area of cutaneous leishmaniasis (Table 1). Of the total animals caught, G. agilis represented 4.1%, of which 75% (3/4) were infected with L. braziliensis. It has been suggested that the presence of a significant number of rodents and marsupials infected with L. (V.) braziliensis could be related to a high rate of transmission and that these animals could be important in the maintenance of the parasite in the area.
The second record was in the federal district of Brasilia during a study in two areas, the Brasília National Park (BNP) and the Contagem Biological Reserve (CBR), which are federal conservation units of different types of habitats including grasslands, cerrado and gallery forests [64] (Table 1). For the two study areas, 19% (3 out 16) of the G. agilis tested positive for Leishmania spp through PCR and sequencing. Although the authors concluded that there were no significant differences in infection rates among the mammal species surveyed, the study highlighted the role of G. agilis, in addition to two rodent species, as hosts of Leishmania spp [64] .
8. The Genus Monodelphis (Short-Tailed Opossums) and Leishmania
Currently the genus Monodelphis is comprised of 20 species: M. adusta, M. americana, M. brevicaudata, M. dimidiata, M. domestica, M. emiliae, M. glirina, M. iheringi, M. kunsi, M. maraxina, M. osgoodi, M. palliolata, M. rubida. M. scalops, M. sorex, M. theresa, M. umbristriata, M. unistriata, M. reigi and M. ronaldi [65] [66] [67] [68] [69] .
Until now there has been only one record of an interaction between a species of Monodelphis and Leishmania sp., which was a natural infection of a M. domestica (Gray Short-tailed Opossum) in Brazil (Figure 1). In the study, Lima et al. [15] found the natural infection prevalence for M. domestica to be 25% (1/4) with Leishmania (Viannia) spp., in a highly endemic area for zoonotic leishmaniasis in the state of Pernambuco in northeastern Brazil (Table 1).
9. The Genus Philander (Four-Eyed Opossums) and Leishmania
Originally, the genus Philander was assumed to contain only a single species, P. opossum, with the following seven subspecies: P. opossum andersoni, P. o. azaricus, P. o. canus, P. o. grisescens, P. o. melanurus, P. o. opossum, and P. o. quica until P. mcilhennyi. Currently seven species, commonly known as Four-eyed Opossums, are recognized for the genus: P. andersoni, P. deltae, P. frenatus, P. mcilhennyi, P. mondolfii, P. olrogi and P. opossum [67] [70] .
There is only one record of infection of a species of Philander with Leishmania sp., which involved P. opossum in Brazil (Figure 1).
Philander opossum (Gray Four-Eyed Opossum) in Brazil
In the only study to have recorded an infection of P. opossum with Leishmania, Lainson et al. [9] observed a prevalence of natural infection of 18% (2/11) with L. amazonensis, in an area highly endemic for cutaneous leishmaniasis in the state of Para. In both infected individuals, the parasite was isolated from the skin (Table 1).
10. Marsupial Hosts of Leishmania in Australia
Although traditionally Australia has been considered to be free of Leishmania, in 2004 the first cases of natural infection were diagnosed in captive Macropus rufus. The first studies involving experimental infection of marsupials with Leishmania occurred in Australia just after World War II in 1945 and 1948.
There have been four studies referencing the infection of marsupials with Leishmania in Australia; two were experimentally infected animals and two were natural infections (Figure 2). Together these studies included three marsupial families: Phalangeridae, Pseudocheiridae and Macropodidae, represented by the genera, Trichosurus, Pseudocheirus and Macropus, respectively.
11. The Genus Macropus
The genus Macropus has 13 extant species which are placed into 3 subgenera: Notamacropus, Osphranter and Macropus. These taxa include several types of terrestrial wallabies, wallaroos and kangaroos, as follows:
Subgenus Notamacropus: Macropus agilis (Agile Wallaby), Macropus dorsalis (Black-striped Wallaby), Macropus eugenii (Tammar Wallaby), Macropus irma (Western Brush Wallaby), Macropus parma (Parma Wallaby), Macropus parryi (Whiptail Wallaby), Macropus rufogriseus (Red-necked Wallaby).
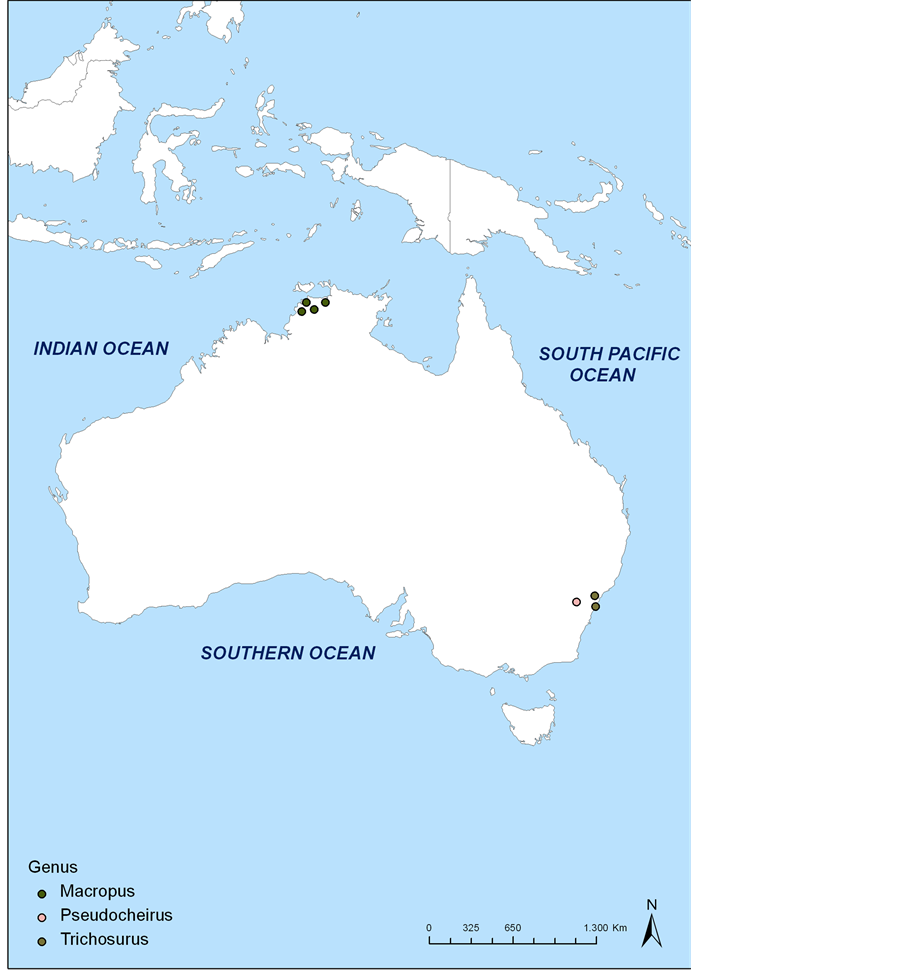
Figure 2. Geographical distribution of records of infection with Leishmania spp. in Trichosurus, Pseudocheirus and Macropus in Australia. All data were mapped through ArchView software using geographical coordinates provided by the authors or estimated based on reference points provided in the studies and utilizing ArchView and Google earth. For experimental infections, the location indicated where the animals were necropsied.
Subgenus Osphranter: Macropus antilopinus (Antilopine Kangaroo), Macropus bernardus (Black Wallaroo), Macropus robustus (Common Wallaroo), Macropus rufus (Red Kangaroo).
Subgenus Macropus: Macropus fuliginosus (Western Grey Kangaroo), Macropus giganteus (Eastern Grey Kangaroo).
Concerning records of Leishmania spp. infection in Australian marsupials, the genus Macropus is the most important with four species that have already been found naturally infected: M. (O.) rufus, M. (O.) robustus, M. (O.) bernardus and M. (N.) agilis (Figure 2). In all cases the species of parasite was not identified.
11.1. Macropus rufus (Red Kangaroo) and Leishmania
Macropus rufus was the first mammal species to be documented as naturally infected with Leishmania spp. in Australia by Rose et al. [71] . The authors observed that 11 kangaroos from Northern Territory had chronic, sometimes self-resolving, skin lesions and in 6 out of those 11 the parasites were isolated and characterized as Leishmania spp.
These findings raised speculation of the possibility of the local occurrence of unrecognized human cases of cutaneous leishmaniasis caused by an imported parasite that could become endemic in Australia (Table 5).
11.2. Macropus robustus (Northern Wallaroo) and Leishmania
Dougall et al. [72] observed that seven out of eight individuals of M. robustus that presented skin lesions suggestive of cutaneous leishmaniasis were seropositive to Leishmania in ELISA and the parasite was also isolated in culture (Table 5). In addition, PCR results demonstrated that the parasite corresponded to the same one observed in M. rufus by Rose et al. [71] .
11.3. Macropus bernardus (Black Wallaroo) and Leishmania
Dougall et al. [72] reported one individual of M. bernardus to be PCR positive to Leishmania spp. This individual exhibited ear lesions similar to those observed in M. robustus, but also small nodular lesions around the eyes, indicating possible mucosal involvement (Table 5).
11.4. Macropus agilis (Agile Wallaby) and Leishmania
Dougal et al. [72] observed that samples collected from sections of the ears from
Table 5. List of records of marsupials infected with Leishmania spp. in Australia.
Natural infection (N), Experimental infection (E), Histology (H), Culture (C), Imprint (I), Molecular Biology (MB), Smear (S), Enzyme-linked Immunosorbent assay (ELISA), Electron microscopy (EM), (a) Originally referred to by the authors as Pseudocheirus peregrinus.
two individuals of M. agilis tested positive for Leishmania spp by PCR test (Table 5).
Conversely, it was reported that M. agilis showed less severe symptoms than both M. robustus and M. bernardus. Moreover the lesions observed in M. agilis healed significantly faster, indicating a probable inherent immunity, as was suggested by the authors.
12. The Genus Trichosurus (Brushtail Possum)
There remains controversy over whether the number of species in the genus Trichosurus is five or three.
Those who recognize five species include the following: T. vulpecula (Common Brushtail Possum), T. arnhemensis (Northern Brushtail Possum), T. caninus (Short-eared Possum), T. cunninghami (Mountain Brushtail Possum) and T. johnstonii (Coppery Brushtail Possum) [73] . However, some consider Trichosurus arnhemensis and T. johnstonii to be subspecies of T. vulpecula, and thus only three species in the genus [74] .
Until now there have been only two records of Leishmania involving this genus, with both being Trichosurus vulpecula inoculated with L. donovani (Figure 2).
Trichosurus vulpecula (Common Brushtail Possum) and Leishmania
The first study of experimental infection involving a marsupial host was in 1945 when Armytage and Bollinger [75] inoculated a pouch young Trichosurus vulpecula of about two and a half years old, still in the pouch, with, as described by the authors, 1 ml of the broth washings of a tube culture of L. donovani. After ten weeks, the animal was necropsied and in a smear of the spleen, liver and bone marrow revealed numerous amastigotes mainly inside of macrophages. The parasite was also isolated in cultures made from spleen, liver, bone marrow and blood (Table 5).
The second report of infection of T. vulpecula with L. donovani was presented in the study of Bollinger and Backhouse in 1948 [76] . The authors demonstrated that 92% (22/25) of the animals became infected after different schedules of intracardiac and intraperitoneal inoculations, including saline suspension from cultures, fresh blood from injected animals and material from sternal puncture. The number of parasites inoculated was not provided (Table 5).
In general, the pattern of infection was characterized by parasitism disseminated in several tissues including spleen, liver, kidney, brain, eyes, pituitary, adrenal, ovary, testis, prostate gland, lung, alimentary canal, para-cloacal glands and pancreas. The most frequently infected organs were the spleen, liver and kidney, which exhibited parasitism in most animals. These organs were followed by brain, eyes, pituitary and adrenal glands, with other organs being less frequently parasitized. The authors concluded that the Common Brushtail Possum is highly susceptible to experimental L. donovani infections and under suitable conditions those animals could become reservoir hosts of Leishmania sp.
13. The Genus Pseudocheirus (Ringtail Possum)
The genus Pseudocheirus contains five species: P. peregrinus (Common Ringtail Possum), P. occidentalis (Western Ringtail Possum), P. canescens (Lowland Ringtail), P. forbesi (Painted Ringtail) and P. mayeri (Pygmy Ringtail).
There is one record of infection of a species of Pseudocheirus with Leishmania sp., were the authors experimentally infected the species Pseudocheirus laniginosus with L. donovani (Figure 2). However, P. laniginosus is no longer recognized and is currently considered a synonym of P. peregrinus.
Pseudocheirus peregrinus (Common Ringtail Possum) and Leishmania
The only information concerning infection of P. peregrinus with Leishmania comes from Bollinger and Backhouse [76] , who inoculated one animal with L. donovani. The parasites used were probably cultured forms obtained from blood of a previously infected T. vulpecula (Table 5). The infected P. peregrinus exhibited a disease pattern similar to that observed in T. vulpecula, with diffuse parasitism but with a smaller number of organs affected, such as spleen, liver, kidney, brain, eyes, pituitary and para-cloacal glands.
Histopathology found that in the central nervous system, and particularly in the choroid plexus, many endothelial cells were parasitized. In the eye, parasites were observed in both macrophage and endothelial cells of both the ciliary body and the choroid coat. The authors inferred that the Common Ringtail Possum could be a potential reservoir host of Leishmania sp. in Australia.
14. Conclusions
Presently, all records about the infection of marsupials with Leishmania are restricted to the New World and Australia, with Brazil being the country where the majority of the studies were performed.
In the Americas, the first studies occurred in the 1960s and all of them dealt with the family Didelphidae, which contains a great number of species that are sympatric with various species of Leishmania.
The situation in Australia is different. Although the first studies occurred earlier than in the New World (1940s) with experimental infections, it has only been relatively recently (2000s) that cases of natural infections have been reported, and all involving the family Macropodidae.
In general, and primarily in the New World, marsupials show a certain equilibrium in their relationship with Leishmania, with high rates of infection but few or no symptoms and a tendency to exhibit self-resolving skin lesions, suggesting an ancient parasite-host coevolution.
In conclusion, marsupials must be considered one of the most important mammal groups to serve as wild hosts of Leishmania spp. in the World.
In the future, in the New World considering the sylvatic cycles, most marsupial species cited they must maintain its importance as reservoir and probably new others will be described. On the other hand, because of not only the tendency of urbanization principally of the visceral leishmaniasis but also the cutaneous disease, in the peri-urban and urban habitats the didelphis probably will prevail because of its ease adaptability to degraded habitats.
Distinctively in the United States, the occurrence of more recent cases of canine and human leishmaniasis shows a tendency of the disease spreading northward. Likewise, the Didelphis also has been presenting a northward dispersion and it could have some roles on the maintenance of the parasite in some of those places.
In respect to Australia, because of the last cases describing autochthonous infections in Kangaroos, several questions must be solved, such as the role of mammal hosts, origin of the infections, the identification of vector species and verifying of the probable role of ceratopogonids in the transmission. It will be very important of the description of the eco-epidemiological picture in the endemic areas, pointing toward prophylactic measures to avoid the occurrence of human infections.
Acknowledgements
To Dr. Erik Wild who reviewed the English.
Competing Interests
Authors declare they have no conflicts of interest.
Author’s Contributions
Carreira JCA-conception, design, drafting and revising of the manuscript. Magalhães MAFM-analysis and interpretation of data of the maps. Brazil RP-re- vising of the manuscript. Silva AVM-drafting the manuscript.
Cite this paper
Carreira, J.C.A., de A.F.M. Magalhães, M., Brazil, R.P. and da Silva, A.V.M. (2017) Leishmania in Marsupials―An Overview of Infection Records in the Americas and Australia. Open Journal of Animal Sciences, 7, 315-343. https://doi.org/10.4236/ojas.2017.73025
References
- 1. Gardner, A.L. (2005) Order Didelphimorphia. In: Wilson, D.E. and Reeder, D.M., Eds., Mammal Species of the World: A Taxonomic and Geographic Reference, 3 rd Edition, Johns Hopkins University Press, 13-16.
- 2. Armati, P., Dickman, C. and Hume, I. (2006) Marsupials. Cambridge University Press, Cambridge.
https://doi.org/10.1017/CBO9780511541889 - 3. Gardner, A.L. (1973) Mammals of South America, Volume 1: Marsupials, Xernarthrans, Shrews, and Bats. The University Chicago Press, Chicago.
- 4. Cerqueira, R. (1985) The Distribution of Didelphis in South America (Polyprotodontia, Didelphidae). Journal of Biogeography, 12, 135-145.
https://doi.org/10.2307/2844837 - 5. Krause, W.J. and Krause, W.A. (2006) The Opossum: Its Amazing Story. University of Missouri Press.
- 6. Lemos, B. and Cerqueira, R. (2002) Morphological Differentiation in the White-Eared Opossum Group (Didelphidae: Didelphis). Journal of Mammalogy, 83, 354-369.
https://doi.org/10.1644/1545-1542(2002)083<0354:MDITWE>2.0.CO;2 - 7. Arias, J.R. and Naiff, R.D. (1981) The Principal Reservoir Host of Cutaneous Leishmaniosis in the Urban Areas of Manaus, Central Amazon of Brazil. Memórias do Instituto Oswaldo Cruz, 76, 279-286.
https://doi.org/10.1590/S0074-02761981000300005 - 8. Arias, J.R., Naiff, R.D., Miles, M.S. and Sousa, A.A. (1981) The Opossum (Didelphis marsupialis (Marsupiala Didelphidae) as a Reservoir Host of Leishmania braziliensis guyanensis in the Amazon Basin of Brazil. Transactions of the Royal Society of Tropical Medicine and Hygiene, 75, 537-541.
- 9. Lainson, R., Shaw, J.J., Ready, P.D., Miles, M.A. and Povoa, M. (1981) Leishmaniasis in Brazil. XVI Isolation and Identification of Leishmania Species in North Para State, with Particular Reference to Leishmania braziliensis guyanensis Causative Agente of “Pian Bois”. Transactions of the Royal Society of Tropical Medicine and Hygiene, 75, 530-536.
- 10. Schalling, H.D.F.H., da Silva, E.S., Van der Maide, W.F., Schoone, G.J. and Gontijo, C.M.F. (2007) Didelphis marsupialis (Common Opossum): A Potential Reservoir Host for Zoonotic Leishmaniasis in the Metropolitan Region of Belo Horizonte (Minas Gerais, Brazil). Vector-Borne and Zoonotic Diseases, 7, 387-393.
https://doi.org/10.1089/vbz.2006.0651 - 11. Sherlock, I.A., Miranda, J.C., Sadigursky, M. and Grimaldi Júnior, G. (1984) Natural Infection of the Opossum Didelphis albiventris (Marsupialia, Didelphidae) with Leishmania donovani, in Brazil. Memórias do Instituto Oswaldo Cruz, 79, 511.
https://doi.org/10.1590/S0074-02761984000400020 - 12. Sherlock, I.A., Miranda, J.C., Sadigursky, M. and Grimaldi Júnior, G. (1988) Observacoes sobre Calazar em Jacobina, Bahia. VI Investigacoes sobre reservatórios silvestres e comensais. Revista da Sociedade Brasileira de Medicina Tropical, 21, 23-27.
https://doi.org/10.1590/S0037-86821988000100005 - 13. Sherlock, I.A. (1996) Ecological Interactions of Visceral Leishmaniasis in the State of Bahia, Brazil. Memórias do Instituto Oswaldo Cruz, 671-683.
https://doi.org/10.1590/S0074-02761996000600003 - 14. Brandao-Filho, S.P., Brito, M.E.F., Carvalho, F.G., Ishikawa, E.A., Cupolillo, E., Floeter-Winter, L., et al. (2003) Wild and Synanthropic Hosts of Leishmania (Viannia) Braziliensis in the Endemic Cutaneous Leishmaniasis Locality of Amaraji, Pernambuco State, Brazil. Transactions of the Royal Society of Tropical Medicine and Hygiene, 97, 291-296.
- 15. Lima, B.S., Dantas-Torres, F., Carvalho, M.R., Marinho-Junior, J.F., Almeida, E.L., Brito, M.E., et al. (2013) Small Mammals as Hosts of Leishmania spp. in a Highly Endemic Area for Zoonotic Leishmaniasis in North-Eastern Brazil. Transactions of the Royal Society of Tropical Medicine and Hygiene, 107, 592-597.
https://doi.org/10.1093/trstmh/trt062 - 16. Costa, A.P., Ferreira, J.I.G.S., Fournier, G.F.S.R., Lopes, M.G., Ramirez, D., Acosta, I.C.L., et al. (2014) Survey of Leishmania infantum chagasi in Wild and Domestic Animals in Urban Area and Atlantic Rainforest Fragment. Journal of Biodiversity, Bioprospecting and Development, 1, 1-5.
- 17. Quintal, A.P., Ribeiro, E.S., Rodrigues, F.P., Rocha, F.S., Floeter-Winter, L.M. and Nunes, C.M. (2011) Leishmania spp. in Didelphis albiventris and Micoureus paraguayanus (Didelphimorphia: Didelphidae) of Brazil. Veterinary Parasitology, 176, 112 -119.
- 18. Santiago, M.E.B., Vasconcelos, R.O., Fattori, K.R., Munari, D.P., Michelin, A.F. and Lima, V.M.F. (2007) An investigation of Leishmania spp. in Didelphis spp. from urban and peri-urban areas in Bauru (Sao Paulo, Brazil). Veterinary Parasitology, 150, 283-290.
- 19. Quaresma, P.F., Rego, F.D., Botelho, H.A., da Silva, S.R., Moura Junior, R.G., Teixeira Neto, et al. (2011) Wild, Synanthropic and Domestic Hosts of Leishmania in an Endemic Area of Cutaneous Leishmaniasis in Minas Gerais State, Brazil. Transactions of the Royal Society of Tropical Medicine and Hygiene, 105, 579-585.
- 20. Yoshida, E.L.A., Silva, R., Cortez, L.S. and Corrêa, F.M.A. (1979) Encontro de espécie do gênero Leishmania em Didelphis marsupialis aurita no Estado de Sao Paulo, Brasil. Revista do Instituto de Medicina Tropical de Sao Paulo, 21, 110-113.
- 21. Yoshida, E.L.A., Corrêa, F.M.A., Pacheco, R.S., Momen, H. and Grimaldi, G. (1985) Leishmania mexicana in Didelphis marsupialis aurita in Sao Paulo State, Brazil. Revista do Instituto de Medicina Tropical de Sao Paulo, 27, 172.
https://doi.org/10.1590/S0036-46651985000400002 - 22. Yoshida, E.L.A., Cuba, C.A., Pacheco, R.S., Cupolillo, E., Tavares, C.C., Machado, G.M.C., et al. (1993) Description of Leishmania (Leishmania) forattinii sp. n., a New Parasite Infecting Opossums and Rodents in Brazil. Memórias do Instituto Oswaldo Cruz, 88, 397-406.
https://doi.org/10.1590/S0074-02761993000300008 - 23. Carreira, J.C.A., Silva, A.V.M., Pereira, D.P. and Brazil, R.P. (2012) Natural Infection of Didelphis aurita (Mammalia: Marsupialia) with Leishmania infantum in Brazil. Parasites & Vectors, 5, 111-116.
https://doi.org/10.1186/1756-3305-5-111 - 24. Travi, B.L., Osorio, Y., Guarin, N. and Cadena, H. (1998) Leishmania (Leishmania) chagasi: Clinical and Parasitological Observations in Experimentally Infected Didelphis marsupialis, Reservoir of New World Visceral Leishmaniasis. Experimental Parasitology, 88, 73-75.
https://doi.org/10.1006/expr.1998.4214 - 25. Molina, R., Amela, C., Nieto, J., San-Andres, M., Gonzalez, F., Castillo, J.A., et al. (1994) Infectivity of Dogs Naturally Infected with Leishmania infantum to Colonized Phlebotomus perniciosus. Transactions of the Royal Society of Tropical Medicine and Hygiene, 88, 491-493.
- 26. Corredor, A., Gallego, J.F., Tesh, R.B., Peláez, D., Diaz, A., Montilla, M., et al. (1989) Didelphis marsupialis, an Apparent Wild Reservoir of Leishmania donovani chagasi in Colombia, South America. Transactions of the Royal Society of Tropical Medicine and Hygiene, 83, 195.
- 27. Travi, B.L., Jaramillo, C., Montoya, J., Segura, I., Zea, A., Goncalves, A., et al. (1994) Didelphis marsupialis, an Important Reservoir of Trypanosoma (Schizotrypanum) cruzi and Leishmania (Leishmania) chagasi in Colombia. The American Journal of Tropical Medicine and Hygiene, 50, 557-565.
https://doi.org/10.4269/ajtmh.1994.50.557 - 28. Adler, G.H., Becerra, M.T. and Travi, B.L. (2003) Feeding Success of Lutzomyia evansi (Diptera: Psychodidae) Experimentally Exposed to Small Mammal Hosts in an Endemic Focus of Leishmania chagasi in Northern Colombia. Biomedica, 23, 396-400.
https://doi.org/10.7705/biomedica.v23i4.1233 - 29. Corredor, A., Gallego, J.F., Tesh, R.B., Morales, A., De Carrasquilla, C.F., Young, D.G., et al. (1989) Epidemiology of Visceral Leishmaniasis in Colombia. The American Journal of Tropical Medicine and Hygiene, 40, 480-486.
https://doi.org/10.4269/ajtmh.1989.40.480 - 30. Travi, B.L., Osorio, Y., Becerra, M.T. and Adler, G.H. (1998) Dynamics of Leishmania chagasi Infection in Small Mammals of the Undisturbed and Degraded Tropical Dry Forests of Northern Colombia. Transactions of the Royal Society of Tropical Medicine and Hygiene, 92, 275-278.
- 31. Alexander, B., Lozano, C., Barker, D.C., McCann, S.H.E. and Adlee, G.H. (1998) Detection of Leishmania (Vianna) braziliensis Complex in Wild Mammals from Colombian Coffee Plantations by PCR and DNA Hybridization. Acta Tropica, 69, 41-50.
- 32. Ocampo, C.B., Ferro, M.C., Cadena, H., Gongora, R., Pérez, M., Valderrama-Adilla, C.H., et al. (2012) Environmental Factors Associated with American Cutaneous Leishmaniasis in a New Andean Focus in Colombia. Tropical Medicine & International Health, 17, 1309-1317.
https://doi.org/10.1111/j.1365-3156.2012.03065.x - 33. Scorza, J.V., Rezzano, S. and Márquez, J.C. (1986) Didelphis marsupialis, reservorio primario de Leishmania spp. en la ciudad de Trujillo, Venezuela. Bol Dir Malariol Saneam Amb, 26, 1-5.
- 34. Zulueta, A.M., Villarroel, E., Rodriguez, N., Feliciangeli, M.D., Mazzarri, M., Reyes, O., et al. (1999) Epidemiologic Aspects of American Visceral Leishmaniasis in an Endemic Focus in Eastern Venezuela. The American Journal of Tropical Medicine and Hygiene, 61, 945-950.
https://doi.org/10.4269/ajtmh.1999.61.945 - 35. Lainson, R. and Strangways-Dixon, J. (1962) Dermal Leishmaniasis in British Honduras: Some Host-Reservoirs of Leishmania braziliensis mexicana. A Preliminary Note. The British Medical Journal, 1, 1596-1598.
https://doi.org/10.1136/bmj.1.5292.1596 - 36. Lainson, R. and Strangways-Dixon (1964) The Epidemiology of Dermal Leishmaniasis in British Honduras. Part II. Reservoir-Hosts of Leishmania mexicana among the Forest Rodents. Transactions of the Royal Society of Tropical Medicine and Hygiene, 58, 136-153.
- 37. Llanos-Cuenta, E.A., Roncal, N., Villaseca, P., Paz, L., Ogusuku, E., Perez, J.E., Caceres, A. and Davies, C.R. (1999) Natural Infections of Leishmania peruviana in Animals in the Peruvian Andes. Transactions of the Royal Society of Tropical Medicine and Hygiene, 93, 15-20.
- 38. Hanson, W.L., Chapman, W.L. Jr. and Hendricks, L.D. (1980) Leishmania donovani in the Opossum (Didelphis marsupialis). Journal of Parasitology, 66, 700-701.
https://doi.org/10.2307/3280544 - 39. White, M.R., Chapman, W.L. and Hanson, W.L. (1989) A Comparison of Experimental Visceral Leishmaniasis in the Opossum, Armadillo and Ferret. Laboratory Animal Science, 39, 47-50.
- 40. White, M.R., Chapman, W.L. Jr., Hanson, W.L., Latimer, K.S. and Greene, C.E. (1989) Experimental Visceral Leishmaniasis in the Opossum. Veterinary Pathology, 26, 314-321.
https://doi.org/10.1177/030098588902600405 - 41. Rossi, R.V., Voss, R.S. and Lunde, D.P. (2010) A Revision of the Didelphid Marsupial Genus Marmosa. Part 1, The Species in Tate’s Mexicana and Mitis Sections and Other Closely Related Forms. Bulletin of the American Museum of Natural History, 334, 30.
https://doi.org/10.1206/334.1 - 42. Tate, G.H.H. (1933) A Systematic Revision of the Marsupial Genus Marmosa with a Discussion of the Adaptive Radiation of the Murine Opossums (Marmosa). Bulletin of the American Museum of Natural History, 66, 1-250.
- 43. Gardner, A.L. and Creighton, G.K. (1989) A New Generic Name for Tate’s microtarsus Group of South American Mouse Opossums (Marsupialia: Didelphidae). Proceedings of the Biological Society of Washington, 102, 3-7.
- 44. Patton, J.L., da Silva, M.N.F. and Malcolm, J.R. (1996) Hierarchical Genetic Structure and Gene Flow in Three Sympatric Species of Amazonian Rodents. Molecular Ecology, 5, 229-238.
https://doi.org/10.1111/j.1365-294X.1996.tb00310.x - 45. Jansa, S.A. and Voss, R.S. (2000) Phylogenetic Studies on Didelphid Marsupials I. Introduction and Preliminar Results from Nuclear IRBP Gene Sequences. Journal of Mammalian Evolution, 7, 43-77.
https://doi.org/10.1023/A:1009465716811 - 46. Steiner, C.M., Tilak, E.J.P., Douzer, Y. and Catzeflis, F.M. (2005) New DNA Data from Transthyretin Nuclear Intron Suggest an Oligocene to Miocene Diversification of Living South American Opossums (Marsupialia: Didelphidae). Molecular Phylogenetics and Evolution, 35, 363-379.
- 47. Voss, R.S. and Jansa, S.A. (2003) Phylogenetic Studies on Didelphid Marsupials. II. Nonmolecular Data and New IRBP Sequences: Separate and Combined Analyses of Didelphine Relationships with Denser Taxon Sampling. Bulletin of the American Museum of Natural History, 276, 1-82.
https://doi.org/10.1206/0003-0090(2003)276<0001:PSODMI>2.0.CO;2 - 48. Voss, R.S., Tarifa, T. and Yensen, E. (2004) An Introduction to Marmosops (Marsupialia: Didelphidae), with the Description of a New Species from Bolivia and Notes on the Taxonomy and Distribution of Other Bolivian Forms. American Museum Novitates, 3466, 1-40.
2.0.CO;2>https://doi.org/10.1206/0003-0082(2004)466<0001:AITMMD>2.0.CO;2 - 49. Voss, R.S., Lunde, D.P. and Jansa, S.A. (2005) On the Contents of Gracilinanus Gardner and Creighton, 1989, with the Description of a Previously Unrecognized Clade of Small Didelphid Marsupials. American Museum Novitates, 3481, 31.
https://doi.org/10.1206/0003-0082(2005)482[0001:otcogg]2.0.co;2 - 50. Voss, R.S. and Jansa, S.A. (2009) Phylogenetic Relationships and Classification of Didelphid Marsupials, and Extant Radiation of New World Metatherian Mammals. Bulletin of the American Museum of Natural History, 332, 177.
- 51. Voss, R.S., Gutiérrez, E.E., Solari, S., Rossi, R.V. and Jansa, S.A. (2014) Phylogenetic Relationships of Mouse Opossums (Didelphidae, Marmosa) with a Revised Subgeneric Classification and Notes on Sympatric Diversity. American Museum Novitates, 3817, 1-27.
https://doi.org/10.1206/3817.1 - 52. Lainson, R. and Shaw, J.J. (1969) Leishmaniasis in Brazil: III. Cutaneous leishmaniasis in an Opossum, Marmosa murina (Marsupialia, Didelphidae) from the Lower Amazon Region. Transactions of the Royal Society of Tropical Medicine and Hygiene, 63, 738-740.
- 53. Lainson, R. and Shaw, J.J. (1970) Leishmaniasis in Brazil: V. Studies on the Epidemiology of Cutaneous Leishmaniasis in Mato Grosso State, and Observations on Two Distinct Strains of Leishmania Isolated from Man and Forest Animals. Transactions of the Royal Society of Tropical Medicine and Hygiene, 64, 654-667.
- 54. Miles, M.A., Póvoa, M.M., Souza, A.A., Lainson, R. and Shaw, J.J. (1980) Some Methods for the Enzymic Characterization of Latin-American Leishmania with Particular Reference to Leishmania mexicana amazonensis and Subspecies of Leishmania hertigi. Transactions of the Royal Society of Tropical Medicine and Hygiene, 74, 243-252.
- 55. Tikasingh, E.S. (1969) Leishmaniasis in Trinidad. A Preliminary Report. Transactions of the Royal Society of Tropical Medicine and Hygiene, 63, 411.
- 56. Tikasingh, E.S. (1974) Enzootic Rodent Leishmaniasis in Trinidad, West Indies. PAHO Bulletin, 3, 232-242.
- 57. Herrer, A., Telford, S.R. and Christensen, H.A. (1971) Enzootic Cutaneous Leishmaniasis in Eastern Panama. I. Investigation of the Infection among Forest Mammals. Annals of Tropical Medicine and Parasitology, 65, 349-358.
https://doi.org/10.1080/00034983.1971.11686764 - 58. Telford, S.R., Herrer, A. and Christensen, H.A. (1972) Enzootic Cutaneous Leishmaniasis in Eastern Panama. Ill. Ecological Factors Relating to the Mammalian Hosts. Annals of Tropical Medicine and Parasitology, 66, 173-179.
https://doi.org/10.1080/00034983.1972.11686813 - 59. Herrer, A., Christensen, H.A. and Beumer, R.J. (1973) Detection of Leishmanial Activity in Nature by Means of Sentinel Animals. Transactions of the Royal Society of Tropical Medicine and Hygiene, 67, 870-879.
- 60. Van Wynsbergue, N.R., Canto-Lara, S.B., Sosa-Bibiano, E.L., Rivero-Cardenas, N.A. and Andrade-Narvaez, F.J. (2009) Comparison of Small Mammal Prevalence of Leishmania (Leishmania) mexicana in Five Foci of Cutaneous Leishmaniasis in the State of Campeche, Mexico. Revista do Instituto de Medicina Tropical de Sao Paulo, 51, 87-94.
https://doi.org/10.1590/S0036-46652009000200006 - 61. Matschie (1916) Marmosops. Sitzungsberichte der Gesellschaft Naturforschender Freunde, 1, 267.
- 62. Lund, P.W. (1840) View of the Fauna of Brasil, Previous to the Last Geological Revolution. The Magazine of Natural History, 4.
- 63. Johnson, P. and Hertig, M. (1970) Behavior of Leishmania in Panamanian Phlebotominae Sandflies Fed in Infected Animal. Experimental Parasitology, 27, 281-300.
- 64. Cardoso, R.M., Araujo, N.N.S.L., Romero, G.A.S., Souza, T.T.C.M., Dietrich, A.G., Mendes, J.D., et al. (2015) Expanding the Knowledge about Leishmania Species in Wild Mammals and Dogs in the Brazilian Savannah. Parasites & Vectors, 8, 171-179.
https://doi.org/10.1186/s13071-015-0780-y - 65. Solari, S.A. (2004) New Species of Monodelphis (Didelphimorphia: Didelphidae) from Southeastern Peru. Mammalian Biology, 69, 145-152.
https://doi.org/10.1078/1616-5047-00129 - 66. Lew, D. and Pérez-Hernández, R. (2004) Una nueva especie del género Monodelphis (Didelphimorphia: Didelphidae) de la Sierra de Lema, Venezuela. Memoria de la Fundación La Salle de Ciencias Naturales, 159-160, 7-25.
- 67. Gardner, A.L. (2007) Mammals of South America. Volume 1 Marsupials, Xenarthrans, Shrews, and Bats. The University of Chicago Press, Chicago.
- 68. Lim, B.K., Engstrom, M.D., Patton, J.C. and Bickham, J.W. (2010) Molecular Phylogenetics of Reig’s Short-Tailed Opossum (Monodelphis reigi) and Its Distributional Range Extension into Guyana. Mammalian Biology, 75, 287-293.
- 69. (2011) IUCN Red List of Threatened Species. Epi Info. http://www.iucnredlist.org
- 70. Patton, J. and da Silva, M. (2007) Genus Philander Brisson, 1762. In: Gardner, Ed., Mammals of South America: Marsupials, Xenarthrans, Shrews and Bats, Vol. 1., The University of Chicago Press, Chicago, 27-35.
- 71. Rose, K., Curtis, J., Baldwin, T., Mathis, A., Kumar, B., Sakthianandeswaren, A., et al. (2004) Cutaneous Leishmaniasis in Red Kangaroos: Isolation and Characterization of the Causative Organisms. International Journal for Parasitology, 34, 655-664.
- 72. Dougall, A., Shilton, C., Low Choy, J., Alexander, B. and Walton, S. (2009) New Reports of Australian Cutaneous Leishmaniasis in Northern Australian Macropods. Epidemiology & Infection, 137, 1516-1520.
https://doi.org/10.1017/S0950268809002313 - 73. Wilson, D.E. and Reeder, D.M. (2005) Mammal Species of the World: A Taxonomic and Geographic Reference. 3rd Edition, Johns Hopkins University Press.
- 74. Kerle, J.A. and How, R.A. (2008) Common Brushtail Possum. In: Van Dyck, S. and Strahan, R., Eds., The Mammals of Australia, Reed New Holland, Sydney.
- 75. Armytage, J.E. and Bolliger, A. (1945) Transmission of Kala-Azar to the Pouch Young of the Common Australian Possum (Trichosorus vulpecula). Nature, 155, 145-146.
https://doi.org/10.1038/155145b0 - 76. Bolliger, A. and Backhouse, T.C. (1948) Transmission of Kala Azar to the Australian Marsupials Trichosorus vulpecula and Pseudocheirus laniginosus. Transactions of the Royal Society of Tropical Medicine and Hygiene, 41, 797-814.