D. M. Ribeiro et al. / Agricultural Science 2 (2011) 191-197
Copyright © 2011 SciRes. Openly accessible at http://www.scirp.org/journal/AS/
196
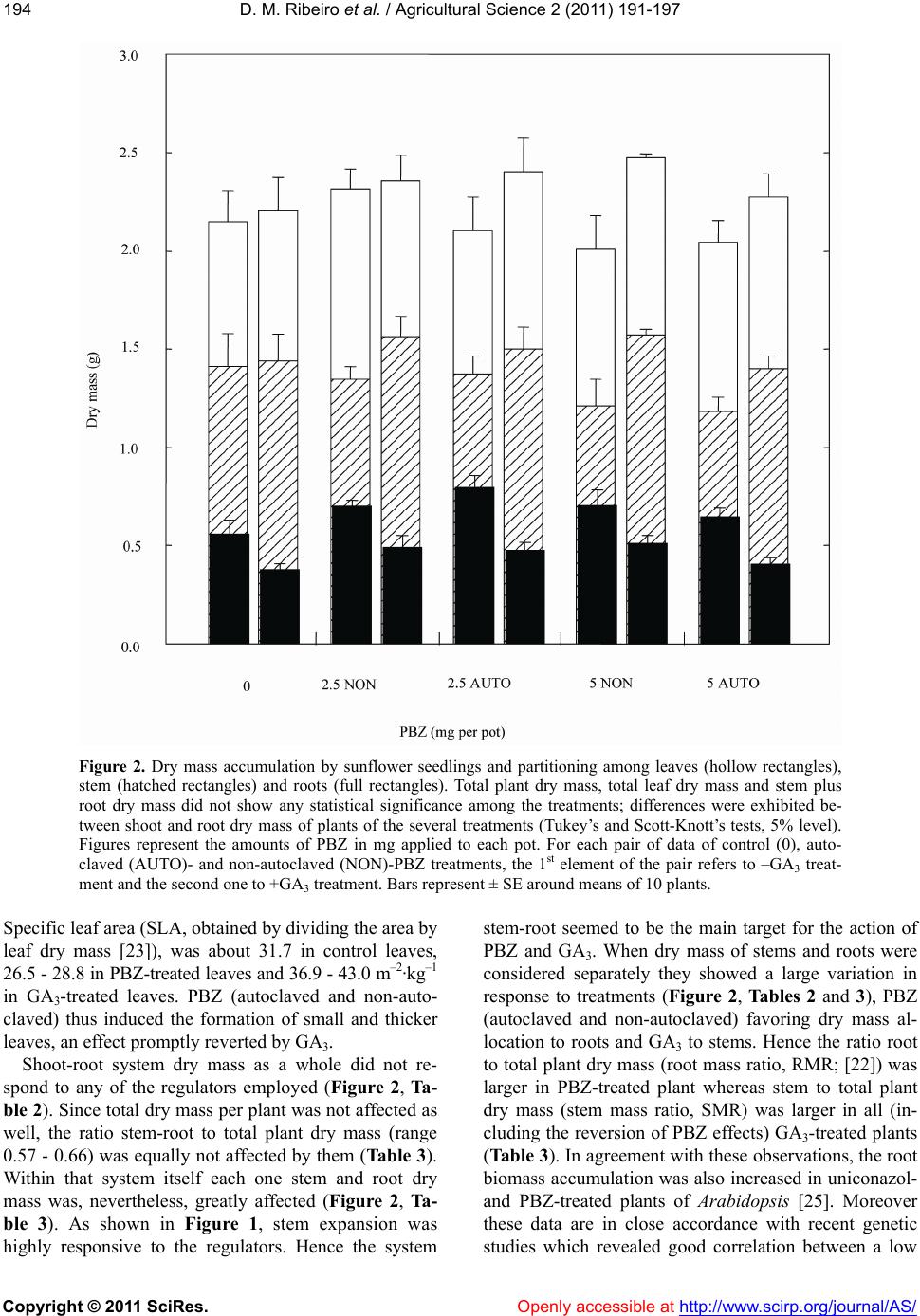
conclusion that GA deficiency caused by PBZ (or in
GA-deficient mutants) resulted in a pronounced stem
growth decrease and hence as stem-root system dry mass
did not vary to a large extent, assimilates are directed
towards the roots. Unequivocally, in the present investi-
gation, not at all affected by both GA3 and PBZ (auto-
claved and non-autoclaved) were mean leaf and inter-
node number, total plant dry mass, leaf dry mass per
plant, and the stem-root system dry mass per plant as a
whole.
4. ACKNOWLEDGMENTS
Thanks are due to FAPEMIG (Foundation for Research Sponsoring
of Minas Gerais State) for the scholarships granted to DMR (Post-
Doctoral training), JB and GBR (Scientific Initiation training), and for
the financial support in several occasions. CAPES (Coordination for
Improvement of Higher Education Training) awarded a scholarship to
CM. CNPq (Brazilian Council for Science and Technological Devel-
opment) has maintained RBS as an associated Fellow Researcher
member since long ago.
REFERENCES
[1] PGRSA (2007) Plant growth regulation handbook of the
Plant Growth Regulation Society of America. 4th Edition.
The Plant Growth Regulation Society of America, Ath-
ens.
[2] ICI (Imperial Chemical Industries) and PLC (1986) Plant
protection division. Clipper, Surrey.
[3] Rademarcher, W. (2000) Growth retardants: Effects on
gibberellins biosynthesis and other metabolic pathways.
Annual Review of Plant Physiology and Plant Molecular
Biology, 51, 501-531.
doi:10.1146/annurev.arplant.51.1.501
[4] Kulkarni, V.J. (1988) Chemical control of tree vigour and
promotion of flowering and fruiting in mango (Mangif-
era indica L.) using paclobutrazol. The Journal of Hor-
ticultural Science, 63, 557-566.
[5] Wilkinson, R.I. and Richards, D. (1991) Influence of
paclobutrazol on growth and flowering of Rhododrendon
‘Sir Robert Peal’. HortScience, 26, 282-284.
[6] Tukey, L.D. (1983) Vegetative control and fruiting on
mature apple trees treated with PP333. Acta Horticulture,
137, 103-109.
[7] Gosh, A., Chikara, J., Chandhari, D.R., Prakashi, A.R.,
Boricha, G. and Zala, A. (2010) Paclobutrazol arrests
vegetative growth and unveils unexpressed yield poten-
tial of Jathropa curcas. Journal of Plant Growth Regula-
tion, 29, 307-315. doi:10.1007/s00344-010-9137-0
[8] Early, J.D. and Martin, G.C. (1988) Translocation and
breakdown of 14C-labeled paclobutrazol in ‘Nemaguaro’
peach seedlings. HortScience, 23, 196-200.
[9] Müller, C. (2007) Efeito do paclobutrazol na germinação
e no metabolismo do etileno em sementes de Stylosantes
humilis HBK. M.Sc. Dissertation, Universidade Federal
de Viçosa, Viçosa.
[10] Snir, I. (1988) Influence of paclobutrazol on in vitro
growth of sweet cherry shoots. HortScience, 23, 304-305.
[11] Panaia, M., Senaratna, T., Bunn, E., Dixon, K.W. and
Sivasithamparam, K. (2000) Micropropagation of the
critically endangered Western Australian species, Sy-
monanthus bancroftii (F. Muell.), L. Haegi (Solanaceae).
Plant Cell Tissue and Organ Culture, 63, 23-29.
doi:10.1023/A:1006457624282
[12] Albany, N.R., Vilchez, J.A., Garcia, L. and Jiménez, E.
(2005) Comparative study of morphological parameters
of Grand Nain banana (Musa AAA) after in vitro multi-
plication with growth retardants. Plant Cell Tissue and
Organ Culture, 83, 357-361.
doi:10.1007/s11240-005-6307-z
[13] Haughan, P.A., Burden, R.S., Lenton, J. and Goad, J.
(1989) Inhibition of celery cell growth and sterol bio-
synthesis by the enantiomers of paclobutrazol. Phyto-
chemistry, 3, 781-787.
doi:10.1016/0031-9422(89)80115-3
[14] Chen, J.-T. and Chang, W.-C. (2003) Effects of GA3,
ancymidol, cycocel and paclobutrazol on direct somatic
embryogenesis of Oncidium in vitro. Plant Cell Tissue
and Organ Culture , 72, 105-108.
doi:10.1023/A:1021235700751
[15] Kucharska, D. and Olikowska, T. (2008) The influence of
paclobutrazol in the rooting medium on the quality of
chrysanthemum vitroplants. The Journal of Fruit and
Ornamental Plant Research, 16, 417-424.
[16] Kyte, L. and Kleyn, J. (1996) Plant from test tubes: An
introduction to micropropagation. 3rd Edition. Timber
Press, Portland.
[17] Richards, F.J. (1969) The quantitative analysis of growth.
In: Steward, F.C., Ed., Plant Physiology—a Treatise,
Academic Press, New York, 3-76.
[18] Hunt, R., Causton, D.R., Shipley, B. and Askey, A.P.
(2002) A modern tool for classical plant growth analysis.
Annals of Botany, 90, 485-488. doi:10.1093/aob/mcf214
[19] LeCain, D.R., Schekel, K.A. and Wample, R.L. (1986)
Growth retarding effects of paclobutrazol on weeping fig.
HortScience, 21, 1150-1152.
[20] Bultynck, L. and Lambers, H. (2004) Effects of applied
gibberellic acid and paclobutrazol on leaf expansion and
biomass allocation in two Aegilops species with con-
strasting leaf elongation rates. Physiologia Plantarum,
122, 143-151. doi:10.1111/j.1399-3054.2004.00383.x
[21] Nagel, O.W. and Lambers, H. (2002) Changes in the
acquisition and partitioning of carbon and nitrogen in the
gibberellin-deficient mutants A70 and W335 of tomato
(Solanum lycopersicum L.). Plant, Cell and Environment,
25, 883-891. doi:10.1046/j.1365-3040.2002.00871.x
[22] Auras, N.E. (1997) Efeitos do paclobutrazol sobre a mor-
fologia e anatomia foliar, crescimento e parte aérea, dis-
tribuição de biomassa e trocas gasosas em girassol. D.Sc
Dissertation, Universidade Federal de Viçosa, Viçosa.
[23] Hunt, R. (1990) Basic growth analysis. Unwin Hyman,
London.
[24] Cox, A.C. and Keever, G.J. (1988) Paclobutrazol inhibits
growth of Zinnia and Geranium. HortScience, 23,
1029-1030.
[25] Bidadi, H., Yamaguchi, S., Asahina, M. and Satoh, S.
(2010) Effects of shoot-applied gibberellin/gibberellin-
biosynthesis inhibitors on root growth and expression of
gibberellins biosynthesis genes in Arabidopsis thaliana.
Plant Root, 4, 4-11. doi:10.3117/plantroot.4.4