W. M. Haggag et al. / Agricultural Science 2 (2011) 291-296
Copyright © 2011 SciRes. Openly accessible at http://www.scirp.org/journal/AS/
292
stage. Mangiferin due to its vegetative growth promoting
property tilted the hormonal balance of malformed pani-
cles in favour of vegetative growth resulting into trans-
formation of malformed florets into green leafy struc-
tures [5]. The mangiferin treated strains produced more
aerial hyphae but less pigment. Prolonged mangiferin
treatment affected the saprophytic ability of the strains
but improved its parasitism. The significance of mangiferin
induced changes in evolving the host-specific strain of
F. moniliforme of M. indicia lies in showing that an e-
cological disadvantage of survival in one niche (sapro-
phytic) may prove advantageous in another (parasite)
[8]. The fungal and mite populations were initially posi-
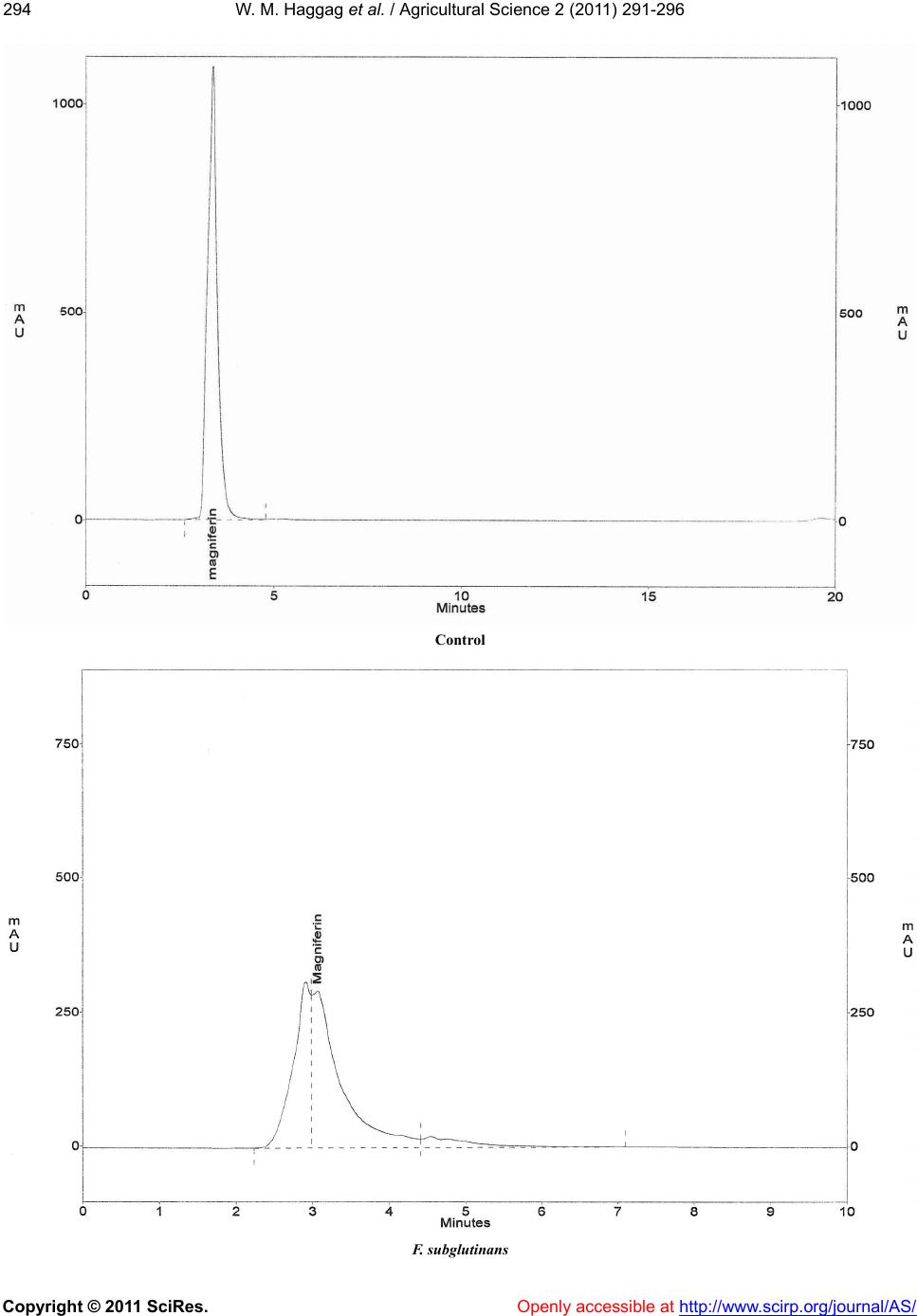
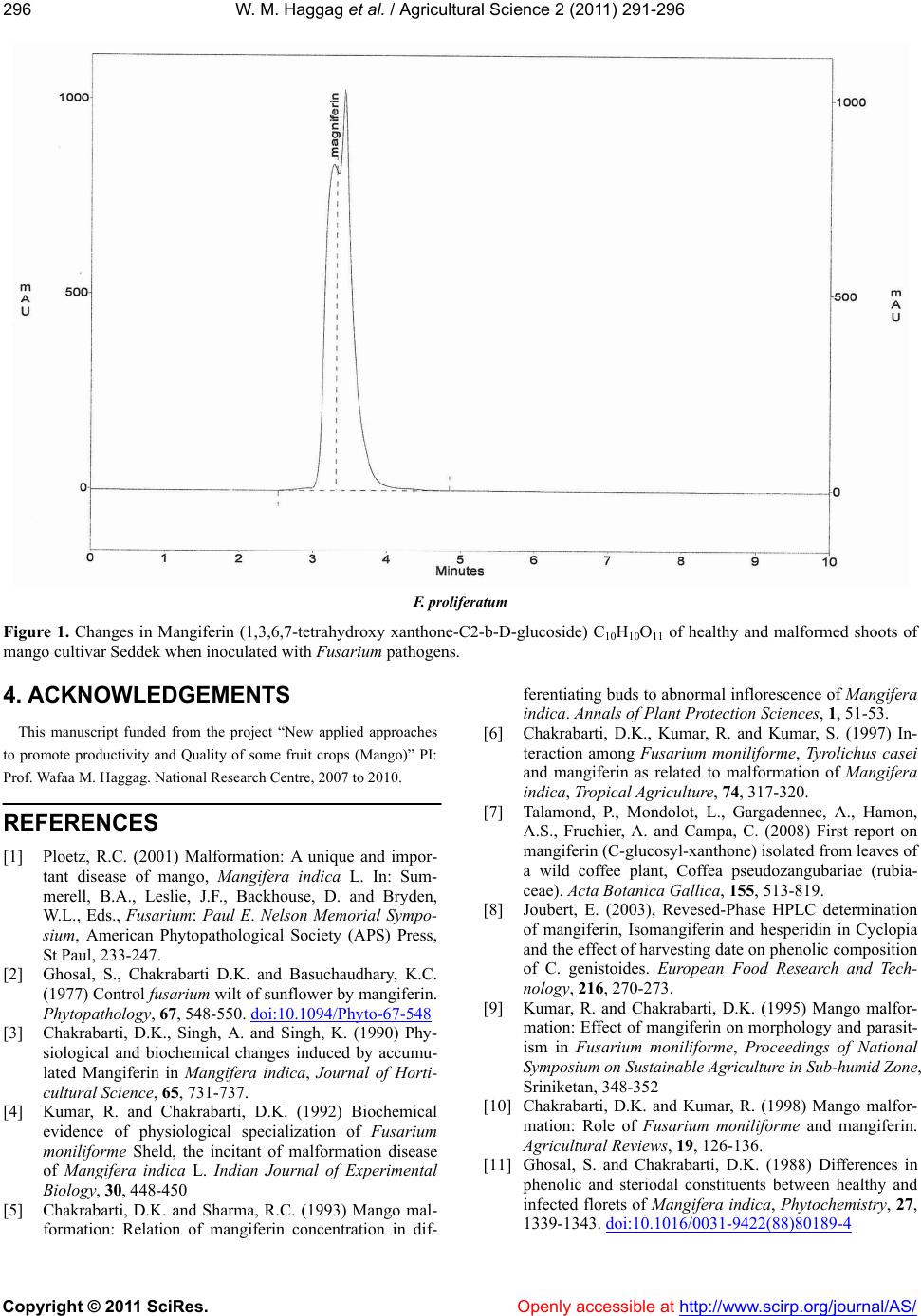
tively related to the mangiferin content and the disease
incidence. Further increase in mangiferin content re-
duced the fungal and mite populations [6]. The symp-
toms are the combined effects of aberrant host metabo-
lites produced in response to infection and phytotoxins
secreted by the pathogen. The pathogen has been identi-
fied as a physiological race of F. moniliforme (F.
moniliforme f. sp. mangifera) developed due to interac-
tion with the host metabolite, mangiferin for a prolonged
period. The disease cycle is greatly influenced with the
biochemical changes in the host tissues. Host metabo-
lites also effect the seasonal variation of population of
the pathogen Vis-a-Vis disease incidence. Proper bal-
ance of mangiferin and the Fusarium population is re-
quired for disease manifestation. Either by suppressing
or avoiding elicitation of hypersensitive reaction of the
host at the initial stage of infection, colonization by the
pathogen and subsequent symptom production could be
effected [3].Thus, objective of the present study is to
evaluate mangiferin change in mango plant due to infec-
tion with pathogenic fungi.
2. MATERIALS AND METHODS
Mango (Mangifera indica L.) seedling cv. Sedekia
(three years old) was inoculated with 105 colony form-
ing units of Fusarium spp. i.e. F. subglutinans , F. ster-
ilihyphosum, F. oxysporum and F. proliferatum, the
causal organisms of mango malformation as inoculated
soil. Sterilized water was used as a control. Transplanted
seedlings were monitored for development of malforma-
tion. At the end of the experiment (120 days), all surviv-
ing seedlings were examined for apical disease symp-
toms. Samples for mangiferin study were taken from
leaves one-two cm below the tip of young seedlings.
2.1. Isolation of Mangiferin
The leaves were macerated with acetone in a high
speed blender. After 4 h, the mixture was filtered and the
solvent was removed under pressure. The extractions
were poured into 100 ml distilled water and the suspen-
sion was successfully extracted with ether and ethyl
acetate. At the interface of the aqueous ethyl acetate,
brown solid mass was precipitated and this was collected
by filtration. The identity of the brown residue was de-
termined as mangiferin according to Ghosal et al. [2].
Subsequently, a small volume of the extract was filtered
through 13 mm membrane filter (0.45 µm; polypropyl-
ene) directly into HPLC sample vial for injection with-
out further dilution.
2.2. Mangiferin Analysis
Extract were done using HPLC system (prominence
Lc, Shimaduz, Kyoto, Japan) equipped with a Lichro-
spher 100RP-18 (5 µm) column (250 mm × 4 mm,
Merck, Darmstadt, Germany), a C18 guard column and a
photoiode-array detector (Shimadzu, SPD-M20A). THE
elution system (0.8 mL·min–1) involved 2 mM phospho-
ric acid in water (eluent A) and MeOH (eluent B). The
gradient was as follows: 0 min, 25% eluent B,0 - 40 min,
80% eluent B, linear the retention time and spectral
characteristics of each sample were compared to a ref-
erence sample of mangiferin (Extrasynthese, Lyon,
France) [7].
2.3. HPLC Analysis and Method
Development
The HPLC system consisted of a ternary solvent pump
(Gynkotek Model 480), autosampler (Gynkotek Gina
50), decade electrochemical detector with a glassy car-
bon electrode (Antec) and a diode array detector
(Gynkotek 340 S). Gynko soft software V5.60 was used
to control the HPLC system and for data acquisition and
analysis. The equipment was supplied by Dionex Softron
(Idstein, Germany). Three columns, i.e. Multosphere
C18 (3 μm; 125; 4 mm ID), Phenomenex Synergy
MAX-RP C12 80 A with TMS end-capping (4 μm; 150;
4.6 mm ID) and Phenomenex Synergi Polar RP (ether
linkedphenyl phase with polar end-capping) were tested
for the chromatographic separation of the above-men-
tioned substances. The Multosphere column was purchased
from CS, Langer-wehe, Germany and Phenomenex, As-
chaffenburg, Germany supplied the Phenomenex col-
umns. Peak identify was determined by means of reten-
tion time and UV spectra that were recorded for all sam-
ples 250 nm. During method development, three solvent
gradients were tested: program I: 0 - 6 min (12% B), 7
min (18% B), 14 min (25% B), 19 min (40% B), 24 min
(50% B), 29 min (12% B) (solvent A = 2% acetic acid in
aqueous solution (v/v) and solvent B = acetonitrile);
program II: 0 min (5% B, 5% C); 4,5 min (6,5% B, 5%
C), 7 min (18% B), 14 min (25% B),19 min (40% B), 24