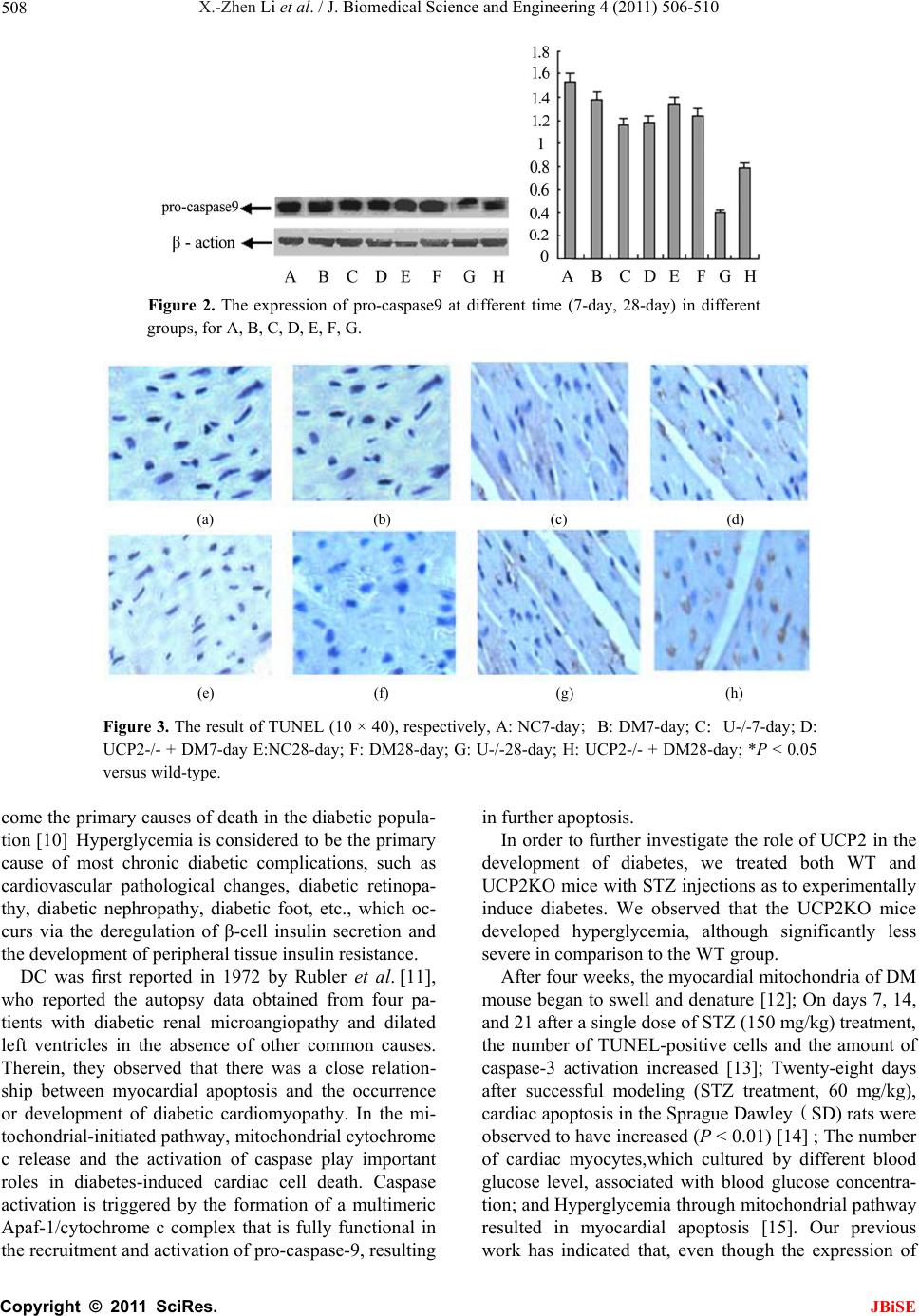
X.-Zhen Li et al. / J. Biomedical Science and Engineering 4 (2011) 506-510 509
cleaved-caspase-3 increased, in addition to cell shrink-
age and nuclear condensation in the DM-7-day group in
comparison to the WT group , this increase was not sta-
tistically significant (P > 0.05), which is consistent with
our reported experimental data.
UCP2 plays a significant role in the onset of diabetes,
specifically via the regulation of cellular ATP produc-
tion [16], islet beta-cell apoptosis [17] and reduction
ROS production [18]. UCP2 also affects the occurrence
or development of DM and participates in the myocar-
dial apoptosis of DC. UCP2KO developed lesser hyper-
glycemia than their WT counterparts after multiple
low-dose STZ injections in vivo [19]. TUNEL analysis
indicated that the number of apoptotic myocytes in the
UCP2KO + DM28-day group decreased by 30% in com-
parison to in the DM-28-day (P <0.05), while the num-
ber of apoptotic myocytes in the UCP2KO + DM7-day
group decreased by 0.2% in comparison to the
DM-7-day (P > 0.05). Our previous work has indicated
that cleaved-caspase-3 expression markedly decreases in
UCP2KO + DM-28-day mice,wherein chromatin was
condensed with high intensity fluorescence in compari-
son to DM-28-day mice, with a statistical significance (P
< 0.05); however, the expression of the cleaved-cas-
pase-3 slightly decreased in the UCP2-/- + DM-7-day
group, wherein chromatin was condensed with low in-
tensity fluorescence, in comparison to the DM-7-day
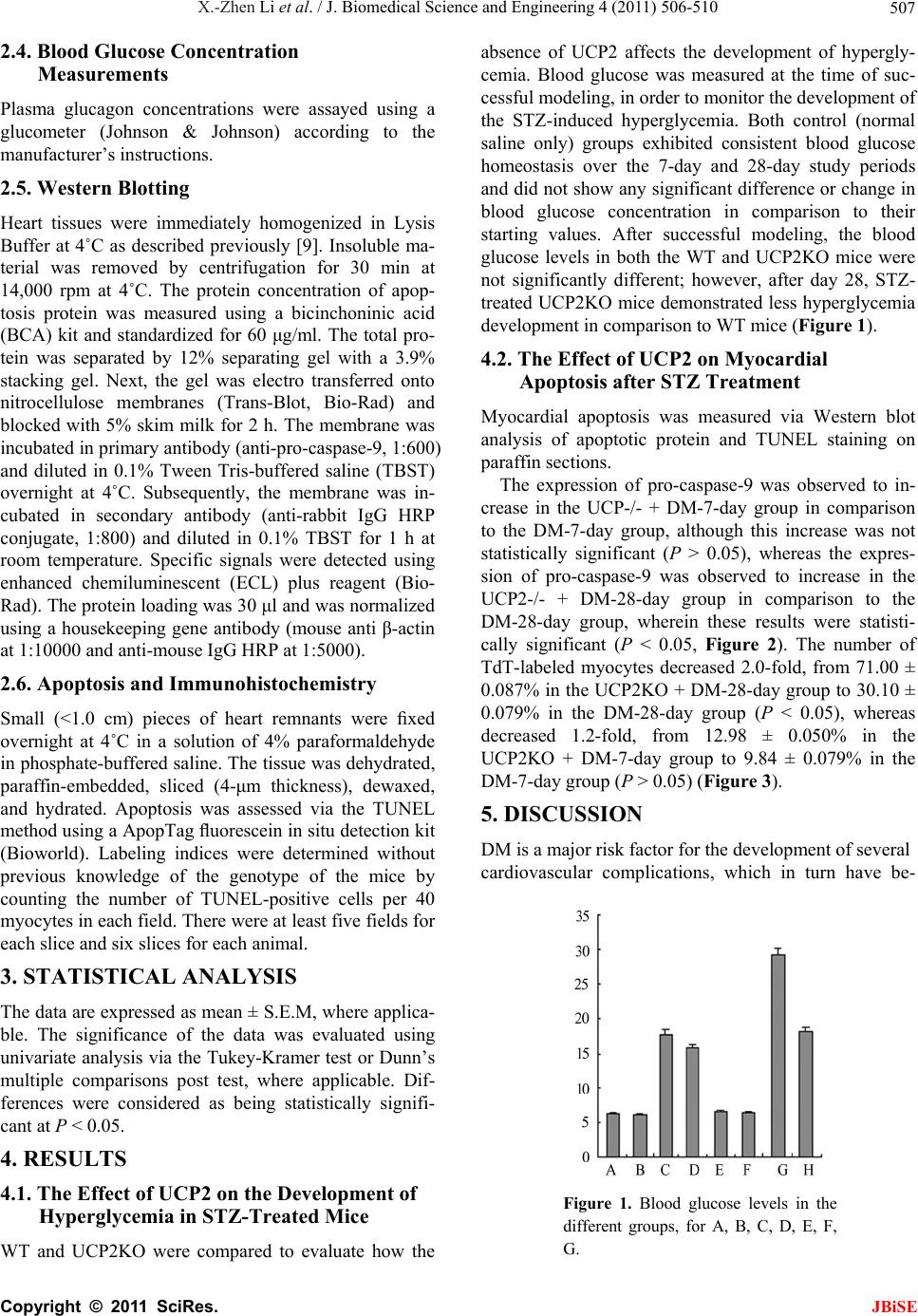
group, although the results were not statistically signifi-
cant (P > 0.05). Therefore, it can be concluded that
UCP2 reduces the number of apoptotic myocytes and
can provide time-dependent anti-apoptosis activity via
variable caspase-3 or caspase-9 expression.
In this article, myocardial apoptosis slightly increased
in the negative control group in comparison to the blank
group, although this increase was not statistically sig-
nificant (P > 0.05). Although cardiac mitochondrial
UCP2-mediated uncoupling respiration was impaired,
short-term mitochondrial efficacy was increased, which
(7-day or 28-day) improved the function of the myocar-
dium.
Diabetic cardiomyopathy results in myocardial dys-
function. Meanwhile,it has been reported that the loss of
myocardial cell quality and quantity could act as an im-
portant factor in heart dysfunction because they are
non-renewable. UCP2 plays a crucial role in mitochon-
drial energy metabolism and in the occurrence or devel-
opment of DC. Furthermore, it is reported that UCP2 is
involved in determining lifespan [20]. Further research
into UCP2 should investigate the relationship between
UCP2 and myocardial apoptosis, and therein, attempt to
identify the causes so as to improve treatment protocols
and provide a more developed basis for the diagnosis
and treatment of DC.
REFERENCES
[1] Ricquier, D. and Bouillaud, F. (2000) The uncoupling pro-
tein homologues: UCP1, UCP2, UCP3, StUCP and AtUCP.
Biochemical Journal, 345, 161-179.
doi:10.1042/0264-6021:3450161
[2] Blanc, J., Alves-Guerra, M.C., Esposito, B., et al. (2003)
Protective role of uncoupling protein 2 in atherosclerosis.
Circulation, 107, 388-390.
doi:10.1161/01.CIR.0000051722.66074.60
[3] Chan, C.B., Saleh, M.C., Koshkin, V., et al. (2004)
Uncoupling protein 2 and islet function. Diabetes, 53,
136-142. doi:10.2337/diabetes.53.2007.S136
[4] Arsenijevic, D., Onuma, H., Pecqueur, C., et al. (2000)
Disruption of the uncoupling protein-2 gene in mice re-
veals a role in immunity and reactive oxygen species
production. Nature Genetics, 26, 435-439.
doi:10.1038/82565
[5] Teshima, Y., Akao, M., Jones, S.P., et al. (2003) Uncou-
pling protein-2 overexpression inhibits mitochondrial
death pathway in cardiomyocytes circulation research.
Circulation Research, 93, 192-200。
doi:10.1161/01.RES.0000085581.60197.4D
[6] Brand, M.D., Buckingham, J.A., Esteves, T.C., et al.
(2004) Mitochondrial superoxide and aging: Uncou-
pling-protein activity and superoxide production. Bio-
chemical Society Symposia, 71, 203-213.
[7] Zhang, C., Baffy, G., Perret, P., et al. (2001) Uncoupling
protein-2 negatively regulates insulin secretion and is a
major link between obesity, β-cell dysfunction, and type2
diabetes. Cell, 105, 745-755.
doi:10.1016/S0092-8674(01)00378-6
[8] King, L.M., Sidell, R.J., Wilding, J.R., Radda, G.K. and
Clarke, K. (2001) Free fatty acids, but not ketone bodies,
protect diabetic rat hearts during low-flow ischemia.
American Journal of Physiology—Heart and Circulatory
Physiology, 280, H1173-H1181.
[9] Li, Z.H., Zhang, T., Dai, H.Y., et al. (2007) Involvement
of endoplasmic reticulum stress in myocardial apoptosis
of streptozocin-induced diabetic rats. Journal of Clinical
Biochemistry and Nutrition, 41, 58-67.
doi:10.3164/jcbn.2007008
[10] Devereux, R.B., Roman, M.J., Paranicas, M., et al. (2000)
Impact of diabetes on cardiac structure and function: The
strong heart study. Circulation, 101, 2271-2276.
[11] Rubler, S., Dlugash, J., Yuceoglu, Y.Z., et al. (1997) New
type of cardiomyopathy associated with diabetic Glomeru-
losclerosis. American Journal of Cardiology, 30, 595-602.
doi:10.1016/0002-9149(72)90595-4
[12] Wei, X.X., Zhou, X.P., He, D., et al. (1992) Cardiac dis-
ease in streptozocin-induced diabetes. Chinese Journal of
Endocrinology and Metabolism, 4, 222-225.
[13] Cai, L., Wang, Y., Zhou, G., et al. (2006) Attenuation by
metallothionein of early cardiac cell death via suppression
of mitochondrial oxidative stress results in a prevention of
diabetic cardiomyopathy. Journal of the American College
of Cardiology, 48, 1688-1697.
doi:10.1016/j.jacc.2006.07.022
[14] Sung, P.H., Sun, C.K., Ko, S.F., et al. (2009) Impact of
hyperglycemic control on left ventricular myocardium a
C
opyright © 2011 SciRes. JBiSE