 Open Access Library Journal How to cite this paper: Milanesi, C., Costantini, L., Firmati, M., Antonucci, F., Faleri, C., Buracchi, A. and Cresti, M. (2014) Geometric Morphometry and Archaeobotany: Characterisation of Grape Seeds (Vitis vinifera L.) by Analysis of Form. Open Access Library Journal, 1: e634. http://dx.doi.org/10.4236/oalib.1100634 Geometric Morphometry and Archaeobotany: Characterisation of Grape Seeds (Vitis vinifera L.) by Analysis of Form Claudio Milanesi1,2*, Lorenzo Costantini3, Marco Firmati2, Francesca Antonucci4, Claudia Faleri1, Ariano Buracchi1, Mauro Cresti1 1Depart men t of Environmental Science, Siena University, Siena, Ita ly 2Archaeological Chianti Museum, Siena, Italy 3Department of Scientific Research, National Museum of Oriental Art “G. Tucci”, Rome, Italy 4Agricultural Engineering Research Unit of the Agricultural Research Council (CRA-ING), Rome, Italy Email: *milan esi@unisi.it Received 6 April 2014; revised 16 May 2014; accepted 26 June 2014 Copyright © 2014 by authors and OALib. This work is licensed under the Creative Commons Attribution International License (CC BY). http://creativ ecommon s.org/l icens es/by/4.0/ Abstract The external profile of seeds is a good phenotypic descriptor, indicative of the origin and diffusion of grape vines (Vi t is v inif era L.) over the centuries. Archaeological seeds conserved in an anaerobic environment after discovery were compared with modern seeds, belonging to eleven autochthon- ous cultivars, sampled in Tuscan vineyards. To study seed profiles we used geometric morpho- metric analysis (elliptical Fourier analysis) combined with multivariate statistical methods. When the values obtained were entered in a database, they produced a cladogram based on Euclidean distances between varieties of archaeological and modern vines, showing th at seeds belonging to the same group had phenotypic affinities. Keywords Archaeob otany, Grape Seeds, Elliptic Fourier Analysis, Etruscan, Viticulture History Subject Areas: Anth ropology, Archaeol ogy, His tory 1. Introduction Archaeobotany involves disciplines such as palynology (study of pollen), anthrocology [anthracology is more frequent] (study of charcoal), phytolith analysis (study of silica bodies of higher plants) and study of diatoms *Corresponding author.  C. Milanesi et al. OALibJ | DOI:10.4236/oalib.1100634 2 June 2014 | Volume 1 | (unicellular siliceous algae). Carpology (study of seeds and fruits) occupies a special place due to an abundance of economic and ecological information. Carpological form is a good phenotypic descriptor that can reveal cli- matic, ecological and sociocul tural cha nges [1] and provide indications about evolution, history and the domes- tication of plants useful to humans [2]. As reported by [3], quantification and description of the form of fruits can help describe and characterize an agricultural cultivar, while in the study by [4], an analytical approach similar to the one used here made it possible to study differences in the form of olive endocarps. During excavations in the period 2011-2013, new well-conserved specimens including archaeological grape seeds were found. With respe ct to the analysis described by [4], the specimens included an additional three ne w archaeological seeds found in Florence (Italy) in spring 2011, three seeds from the excavations conducted at Miranduolo castle (Chiusdino, Siena, Italy) in April-May 2012 and 17 seeds from Poggio Bacherina (Chian- ciano Terme, Siena, Italy). Four archaeological seeds from Poggio la Croce (Radda in Chianti, Siena, Italy) found in 2009 and four seeds from S hahr-i Sokhta (Iran) found in 2007 were also incl uded in the analys is. To ensure their conservation, the seeds found were immediately placed in sterile capsules sealed in vacuo in an anaerobic environment (absence of oxygen) and stored in the refrigerator (0˚C - 4˚C). Later they were characterized morphologically by elliptical Fourier analysis combined with multivariate sta- tistical metho ds to obtain the result s presented in this paper. 2. Analysis of Form Anal ysi s o f the form of a rchaeol ogi ca l a nd mo d er n se eds wa s co nd uc ted anal ysi n g t he fro nta l externa l p rofile o f all specimens as described by [5]. The number of seeds analysed was 813 and they consisted of Tuscan ar- chaeological seeds [Firenze (43˚768591'N, 11˚254714'E, 13th century AD, seven seeds), Miranduolo castle (43˚120348'N, 11˚079410'E, 11th century AD, six seeds), Poggio Bacherina (43˚039420'N, 11˚815203'E, 2nd century BC, twenty seeds), Poggio la Croce (43˚39'45.25"N, 11˚23'45.25"E, 6th century BC, four seeds)], Ira- nian seeds [Shahr-i Sokhta (30˚6500003'N, 61˚399999'E, 23rd century BC, four seeds)] and modern Tuscan seeds [70 seeds per cultivar: Sangiovese Casciano, Sangiovese Montalcino, Cabernet Bossi, Merlot Bossi, Ca- naiolo Monti in Chianti, Canaiolo Montalbuccio, Trebbiano Montalbuccio, Trebbiano Tognazza, Malvasia Monti in Chia nti, Albano Mo ntalbuccio a nd Silvestris (se lected in Septe mber 201 3 in the Mediterr anean biocli- matic area between 43˚02' - 43˚25'N and 11˚25' - 11˚28'E)]. The first name of the modern seeds indicates culti- var and the second provenance, except for Vitis vinifera ssp. sylvestris. They were selected from specimens of autochthonous varieties of ancient Tuscan vineyards, having the highest morphological affinities (size, length and fo rm) wit h the arc haeological varieties. The term autochthonous refers to varieties that have adapted to the host a re a t o t he p o int of b elo n gin g, a nd have t he ir o r igi ns in t he a nc ient hi sto ry of t he ar e a . To analyse form, the seeds were disposed on a back-lighted surface to enhance their profile (Figure 1(a)) and photographed with a digital ca mera at high resolution. An automated procedure was used to transform the photos into black and white (Figure 1(b)), from which the profiles of the seeds were extracted (Figure 1(c)—green). The forms of all seeds were then processed in sequence by elliptical Fourier analysis, extracting the coeffi- cients of the harmonic equations of each to enable comparison of samples of different origins and age, and ex- Figure 1. (a) Original frontal images of grape seeds (Vitis vinifera) on a back-lit surface. (b) The same image converted automatically into black and white. (c) Profiles (green) extracted automatically in a clockwise direction starting from the seed apex (r ed). The co ordinates were then process ed by elliptical Fo urier analysis. 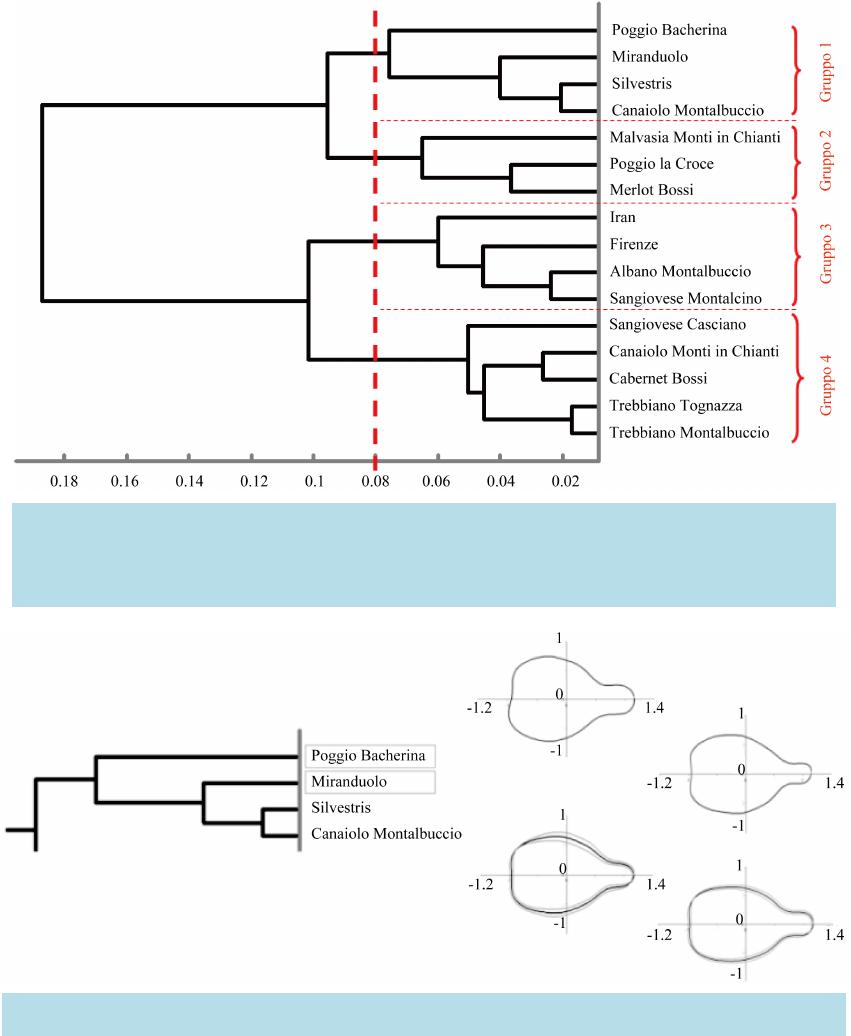 C. Milanesi et al. OALibJ | DOI:10.4236/oalib.1100634 3 June 2014 | Volume 1 | tracting a cladogram based on Euclidean distances that revealed four groups (Figure 2). Figures 3-6 show details of the four groups generated by the cladogram. The mean forms of seeds belonging to each cultivar are sho wn on the right ha nd side . It is evident that seeds of the same group have simi lar for ms. Statistical analysis involved 770 modern seeds (77 per cultivar) which is a good statistical basis for testing hy- potheses of relationship. 2.1. Group 1 The forms of Group 1 (Fig ure 3 ) are round ish with a slight piriform depression in the posterior part. Two culti- Figure 2 . Cladogram based on Euclidean distances of the coefficients of harmonic equations extracted by ellipti- cal Fourier analysis from profiles of archaeological and 770 modern grape seed specimens. The cladogram ex- presses th e degree of mutu al similarit y between sp ecimens: the s horter the di stance of the li nes between two cu l- tivars, the greater their similarit y. The bold dashed red line represents the level at which the cladogram is cut to generate the four groups shown on the right. Figure 3. Group 1 with the mean forms of seeds of the cultivars Poggio Bacherina, Miranduolo, Silvestris and Ca- naiolo Montalbuccio shown on the right. The archaeological specimens are indicat ed in rectangles. 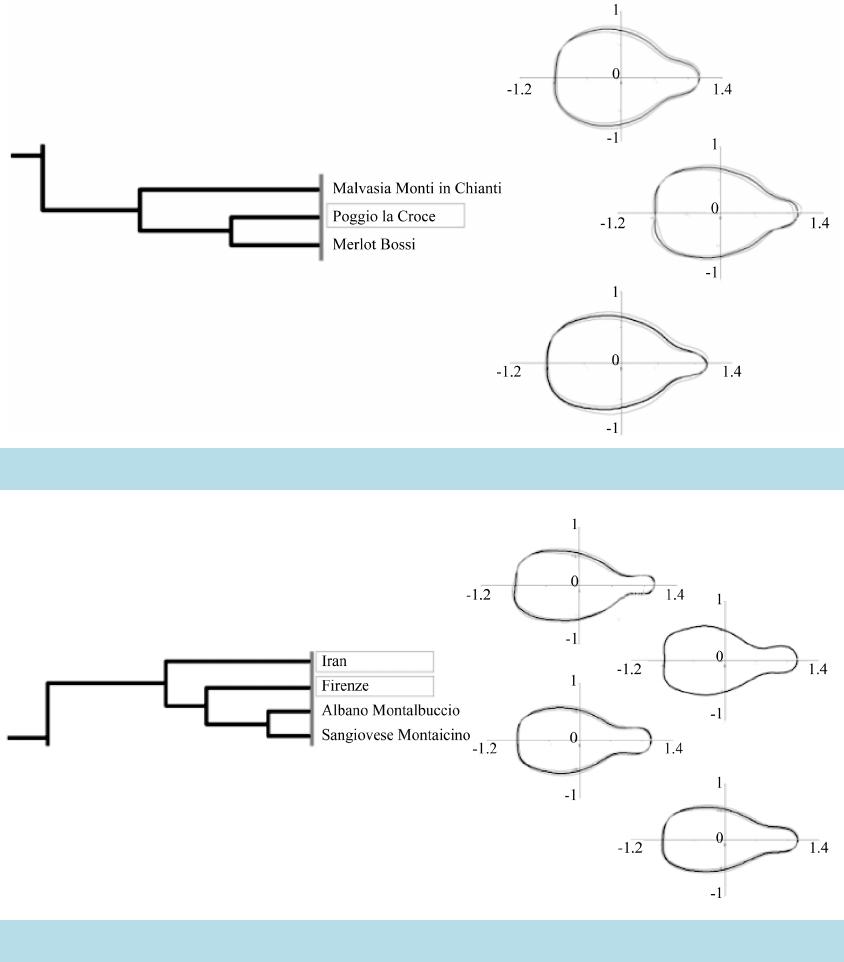 C. Milanesi et al. OALibJ | DOI:10.4236/oalib.1100634 4 June 2014 | Volume 1 | Figure 4. Group 2 with the mean forms of seeds of the cultivars Malvasia Monti in Chianti, Poggio la Croce and Merlo t Bossi shown on the right. The arch aeological specimens are indicat ed in rectangles. Figure 5 . Group 3 with the mean forms of seeds of the cultivars Iran, Firenze, Albano Montalbuccio and Sangiovese Montalcino shown on the right. The archaeological specimens are i ndicated in rectangl es. vars from archaeological sites (Poggio Bacherina and Miranduolo) and two modern cultivars (Silvestris and Ca- naiolo Montalbuccio) belong to this group. The archaeological seeds are further apart than the modern seeds and also distanced from them (longer cladogram distances between cultivars). 2.2. Group 2 The forms belonging to Group 2 (Figure 4) are the roundest. Again the group included archaeological (Poggio la Croce) and modern seeds (Malvasia Monti in C hianti and Merlot Bossi). 2.3. Group 3 The forms belonging to Group 3 (Figure 5) are more elongated. The re mainin g two cul tivar s from ar chae ologi- cal sites (Iran and Firenze) and two modern cultivars (Albano Montalbuccio and Sangiovese Montalcino) belong 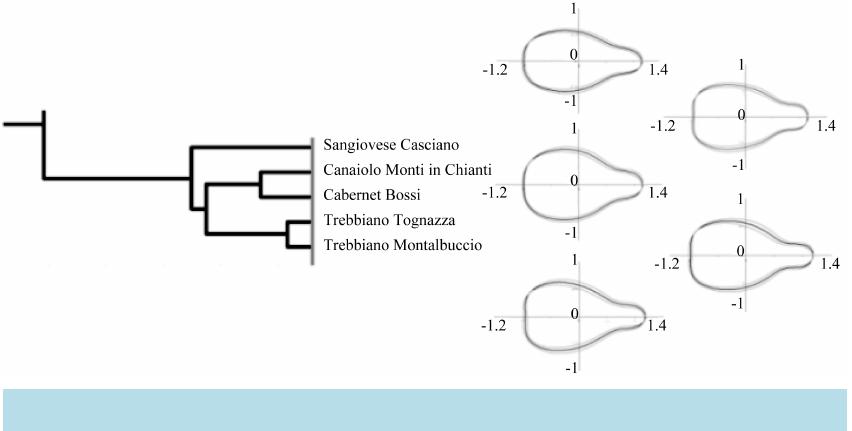 C. Milanesi et al. OALibJ | DOI:10.4236/oalib.1100634 5 June 2014 | Volume 1 | Figure 6. Group 4 with the mean forms of seeds of the cultivars Sangiovese Casciano, Canaiolo Monti in Chianti, Cabernet Bossi, Trebbiano Tognazza and Trebbiano Montalbuccio shown on the right. to thi s gro up . As in Group 1, the archaeological seeds are fur ther apart than the modern seeds and also distanced from them (longer distances bet ween cultivars in the cladogram). 2.4. Group 4 The forms belonging to Group 4 (Figure 6) are slightly elongated and triangular. No archaeological seeds be- longed to t his group. 3. Conclusions In thi s stud y we exa mined mor e than 800 grape seed specimens. The varietal characterisations proved to be par- ticularly descriptive, efficacious and economical. Specifically: • In Group 1, Etruscan archaeological seeds from Poggio Bacherina (2nd century BC) and medieval seeds from Miranduolo (11th century) showed compatibility with modern seeds Canaiolo and Montalbuccio. Ca- naiolo is an ancient Tuscan variety mentioned in “Dè Opera di agricoltura” by Pietro de Crescenzi (1233- 1320). Besides carpological similarities between modern seeds Canaiolo Montalbuccio and Silvestris, we also observed pomological similarities (small leaf, loose bunch, vine trunk up to 40 cm in dia meter). • In Group 2, the affinity between Poggio la Croce (6th century BC) and Merlot Bossi (modern) seeds from Gironde in France is in line with the hypothesis that French wines had origins in Etruscan wines. This is sho wn by resear ch conduct ed on Etruscan amphora e from 6th c entury BC fou nd in the harbour si te of Lattara in the south of France [6]. There is a fascinating affinity between archaeological Etruscan seeds from Poggio la Croce and modern seeds of Malvasia Monti in Chianti. The Etruscans loved light, aromatic, sparkling white wine s that the y drank with rich food s . Etr uscan t ombs and va s es (8-4th century BC) depict vines strung from tall tree s that acted as li ving trellise s. In Tuscany there are still pergolas of Malvasia that produce light, sparkling, thirst-q uenching, low-alco hol white wines. • In Group 3, the affinity of Iranian archaeological seeds (23rd century BC) and Florentine medieval seeds (12 th centur y) wi th the moder n culti var Sa ngiove se Montalcino, the basis of all Tuscan red wines, including Brunello and Rosso di Montalcino, Chianti and Nobile di Montepulciano, adds weight to scientific sources showing that in prehistoric times, domestication of the vine originated in the area between the Black Sea and Iran ([7]-[10]). T his hypothesis may be confirmed by the affinity of archaeological seeds with modern seeds of Albano Montalbuccio, a variety of the Po valley and the area between the Apennines and the Adriatic coast, where the vine presumably entered in Italy in antiquity, imported from the east, and from whence it spread. Finally, the method of morphometric analysis applied to palaeobotanical specimens is immediate and eco- nomica l, but is not enough in itself to e stablish the descent of modern material from ancient specimens. Genetic  C. Milanesi et al. OALibJ | DOI:10.4236/oalib.1100634 6 June 2014 | Volume 1 | studies could provide better correlatio ns, but in our experience the genetic profiles of older palaeobotanic mate- rial entered in databases show tenuous affini ties that can in most cases be calculated at 1% - 3%. Moreover, a DNA studies are destructive of specimens, which precludes the possibility of using seeds from archaeological museums. In future, phylogenetic comparisons of palaeobotanic and modern materials will be sustained by im- proved methods, larger databases and inter disciplinary studies. References [1] Terral, J.F., Alonso, N., Buxò, R., Capdevila, I., Chatti, N., Fabre, L., Fiorentino, G., Marinval, P., Pérez-Jorda, G., Pradat, B., Rovira, N. and Alibert, P. (2004) Historical Biogeography of Olive Domestication (Oleaeuropaea L.) as Revealed by Geometrical Morphometry Applied to Biological and Archaeol ogical Material. Journal of Biogeography, 31, 63-77. http://dx.doi.org/10.1046/j.0305-0270.2003.01019.x [2] Terral, J.F., Tabard, E., Bouby, L., Ivorra, S., Pastor, T., Figueiral, I., Picq, S., Chevance, J.B., Jung, C., Fabre, L., Tardy, C., Compan, M., Bacilieri, R., Lacombe, T. and This, P. (2010) Evolution and History of Grapevine (Vitis vi- nifera) under Do mestication: N ew Morphometric Per s pective to Understand Seed Domestication Syndrome and Reveal Origins and Ancient European Cultivars. Annals of Botany London, 105, 443-455. http://dx.doi.org/10.1093/aob/mcp298 [3] Antonucci, F., Costa, C., Pallottino, F., Paglia, G., Rimatori, V., De Giorgio, D. and Menesatt i, P. (2012) Quantitative Method for Shape Descrip tion of Almond Cultivars (Prunus amygdalus Batsch). Food and Bioprocess Technology, 5, 768-785. http://dx.doi.org/10.1007/s11947-010-0389-2 [4] Milanesi, C., Sorbi, A., Paolucci, E., Antonucci, F., Menesatti, P., Costa, C., Pallottino, F., Vignani, R., Cimato, A., Ciacci, A. and Cresti, M. (2011) Pomology Observations, Morphometric Analysis, Ultrastructural Study and Allelic Profiles of “Oli vastra Seggianese” Endocarps from Ancient Olive Trees (Olea europaea L.). Comptes Rendus Biolo- gies, 334, 39-49. http://dx.doi.org/10.1016/j.crvi.2010.11.006 [5] Milanesi, C., Antonucci, F., Menesatti, P., Costa, C., Faleri, C. and Cresti, M. (2011) Morphology and Molecular Analysis of Ancien t Grape Seeds. Interdisciplinaria Archaeologica, 2, 95-100. [6] McGovern, P., Luley, P.B., Rovira, N., Mirzoian, A., Callahan, M.P., Smith, K.E., Hall, G.R., David son, T. and Hen- kin, J.M. (2012) Beginning of Viticulture in France. Pnas, 110-25, 1014 7-10152. [7] Zohary, D. and Hopf, M. (2000) Domestication of Plants in the Old World. Oxford University Press, New York, 151-159. [8] McGovern, P.E. (2004) Ancient Wine: The Search for the Origins of Viticulture. Princeton University Press, Princeton. [9] This, P., Lacombe, T. and Thomas, M.R. (2006) Historical Origins and genetic Diversity of wine Grapes. Trends in Genetics, 22, 511-519. http://dx.doi.org/10.1016/j.tig.2006.07.008 [10] Pieraccini, L.C. (2011) The Wonder of Wine in Etruria. In: De Grummond, N.T. and Edlund-Berry, I., Eds., The Ar- chaeology of Sanctuaries and Ritual in Etruria, Rhode Island, Portsmouth, 127-138.
|