 Vol.3, No.7, 573-579 (2011) Natural Science http://dx.doi.org/10.4236/ns.2011.37080 Copyright © 2011 SciRes. OPEN ACCESS A quantum-chemical model of the inhibition mechanism of viral DNA HIV-1 replication by Iodine complex compounds Gulnara Abd-Rashidovna Yuldasheva1, Georgii Mihailovich Zhidomirov2, Aleksandr Ivanovich Ilin1 1Anti-Infective Drug Research Center, Almaty, Kazakhstan; *Corresponding Author: yuldasheva57@rambler.ru, ilin_ai@mail.ru 2Boreskov Institute of Catalysis SB RAS, Novosibirsk, Russia; *Corresponding Author: zhidomirov@mail.ru Received 21 March 2011; revised 15 April 2011; accepted 21 April 2011. ABSTRACT The interaction of molecular iodine with virus DNA nucleotide is studied by ab initio RHF/3- 21G** method. Formation of the nucleoprotein complex of the HIV DNA, molecular iodine and the HIV-1 integrase co-factor is considered to cause the inhibition action of the integrase en- zyme. Experimental data on the anti-HIV effect of the molecular iodine complex compounds and the results of calculations suggest that mole- cular iodine contained in iodine polymer com- plexes may be considered as a compound inhi- biting the catalytic center of the integrase en- zyme. Unlike the known integrase inhibitors, molecular iodine also changes the virus DNA structure and produces the N-I bond in the purine bases of adenosine and guanosine nu- cleotides. Keyw ords: HIV; Integrase HIV; Quantum-Chemical Method Ab Initio 1. INTRODUCTION The human immunodeficiency virus (HIV) belongs to the series of retroviruses whose gene within the virion is represented by a RND molecule. After the virus pene- trates a human cell, a DNA copy of the virus genome is synthesized and such virus DNA is integrated into the genome of the human cell. Both processes are produced by the virus enzymes – transcriptase and integrase [1], respectively. A large number of agents inhibiting the activity of the HIV reverse transcriptase have been developed [2] and used for medicine production. However, the HIV reverse transcriptase is referred to the class of polymerases and most of its blockers that, to some extent, suppress the actions of this class of cellular enzymes producing sig- nificant toxic effects [3,4]. HIV integrase inhibitors have a high therapeutic effect [5] for two reasons. First, integrase is one of the key participants in the virus replication cycle [1]. Second, integrase has no cellular equivalent and, hence, the sup- pression of its activity should not disturb normal cellular metabolism processes [6]. HIV-1 integrase contains 288 amino acid residues and three domains can be distinguished in its structure: a short N-end domain containing 1-50 amino acid residues, a catalytic domain producing 51-212 amino acid resi- dues and a C-end domain containing 220-270 amino acid residues. It is typical for such family of enzymes to produce very stable complexes with a virus DNA. To be inte- grated, it is necessary for integrase to bind both - virus and cell - DNA molecules at the same time. The integra- tion proceeds in two stages and begins in the cytoplasm of HIV infected cells where, upon completion of the reverse transcription of the virus DNA genome, integrase binds the virus DNA copy producing the so-called pre- integration complex (PIC) which can be isolated from the HIV-infected cells [7]. A model of the integrase structure in the complex with a virus and a cell DNA was proposed in [8]. According to the model, only the catalytic domain takes part in the binding of the virus DNA, while all the three domains take part in the binding of the cell DNA. The structure of the catalytic domain of the HIV-1 in- tegrase is determined by the X-ray structural analysis. According to the data obtained, the catalytic domain of the enzyme in the crystal forms a spherical dimer with each monomer forming a semi-sphere. The three amino acid residues - Asp64, Asp116 and Glu152 closely lo- cated in the tertiary structure of the catalytic domain – form its active catalytic center with the active centers of each integrase monomer located on the opposite sides of the dimer sphere 35 Å apart. The X-ray structural analysis data clearly show one Mg2+ ion coordinated by Asp64 и Asp116 and two water 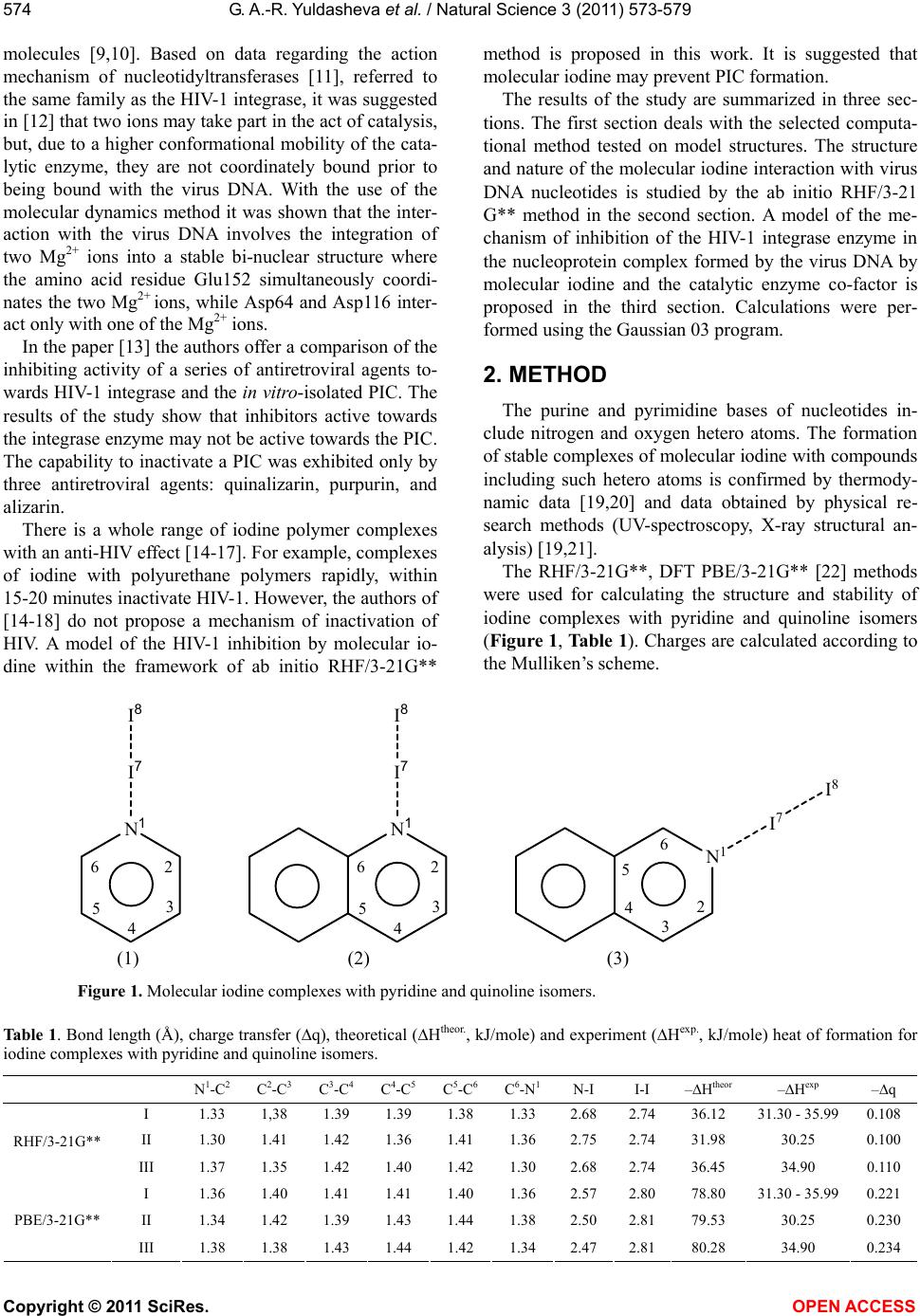 G. A.-R. Yuldasheva et al. / Natural Science 3 (2011) 573-579 Copyright © 2011 SciRes. OPEN ACCESS 574 molecules [9,10]. Based on data regarding the action mechanism of nucleotidyltransferases [11], referred to the same family as the HIV-1 integrase, it was suggested in [12] that two ions may take part in the act of catalysis, but, due to a higher conformational mobility of the cata- lytic enzyme, they are not coordinately bound prior to being bound with the virus DNA. With the use of the molecular dynamics method it was shown that the inter- action with the virus DNA involves the integration of two Mg2+ ions into a stable bi-nuclear structure where the amino acid residue Glu152 simultaneously coordi- nates the two Mg2+ ions, while Asp64 and Asp116 inter- act only with one of the Mg2+ ions. In the paper [13] the authors offer a comparison of the inhibiting activity of a series of antiretroviral agents to- wards HIV-1 integrase and the in vitro-isolated PIC. The results of the study show that inhibitors active towards the integrase enzyme may not be active towards the PIC. The capability to inactivate a PIC was exhibited only by three antiretroviral agents: quinalizarin, purpurin, and alizarin. There is a whole range of iodine polymer complexes with an anti-HIV effect [14-17]. For example, complexes of iodine with polyurethane polymers rapidly, within 15-20 minutes inactivate HIV-1. However, the authors of [14-18] do not propose a mechanism of inactivation of HIV. A model of the HIV-1 inhibition by molecular io- dine within the framework of ab initio RHF/3-21G** method is proposed in this work. It is suggested that molecular iodine may prevent PIC formation. The results of the study are summarized in three sec- tions. The first section deals with the selected computa- tional method tested on model structures. The structure and nature of the molecular iodine interaction with virus DNA nucleotides is studied by the ab initio RHF/3-21 G** method in the second section. A model of the me- chanism of inhibition of the HIV-1 integrase enzyme in the nucleoprotein complex formed by the virus DNA by molecular iodine and the catalytic enzyme co-factor is proposed in the third section. Calculations were per- formed using the Gaussian 03 program. 2. METHOD The purine and pyrimidine bases of nucleotides in- clude nitrogen and oxygen hetero atoms. The formation of stable complexes of molecular iodine with compounds including such hetero atoms is confirmed by thermody- namic data [19,20] and data obtained by physical re- search methods (UV-spectroscopy, X-ray structural an- alysis) [19,21]. The RHF/3-21G**, DFT PBE/3-21G** [22] methods were used for calculating the structure and stability of iodine complexes with pyridine and quinoline isomers (Figure 1, Table 1). Charges are calculated according to the Mulliken’s scheme. N 1 I 7 I 8 2 3 4 5 6 N1 I7 I8 2 3 4 5 6 2 3 4 5 6 N 1 I 7 I 8 (1) (2) (3) Figure 1. Molecular iodine complexes with pyridine and quinoline isomers. Table 1. Bond length (Å), charge transfer (q), theoretical (Htheor., kJ/mole) and experiment (Hexp., kJ/mole) heat of formation for iodine complexes with pyridine and quinoline isomers. N 1-C2 C2-C3 C3-C4 C4-C5 C5-C6 C6-N1 N-I I-I –Htheor –Hexp –q I 1.33 1,38 1.39 1.39 1.38 1.33 2.68 2.7436.12 31.30 - 35.990.108 II 1.30 1.41 1.42 1.36 1.41 1.36 2.75 2.7431.98 30.25 0.100 RHF/3-21G** III 1.37 1.35 1.42 1.40 1.42 1.30 2.68 2.7436.45 34.90 0.110 I 1.36 1.40 1.41 1.41 1.40 1.36 2.57 2.8078.80 31.30 - 35.990.221 II 1.34 1.42 1.39 1.43 1.44 1.38 2.50 2.8179.53 30.25 0.230 PBE/3-21G** III 1.38 1.38 1.43 1.44 1.42 1.34 2.47 2.8180.28 34.90 0.234 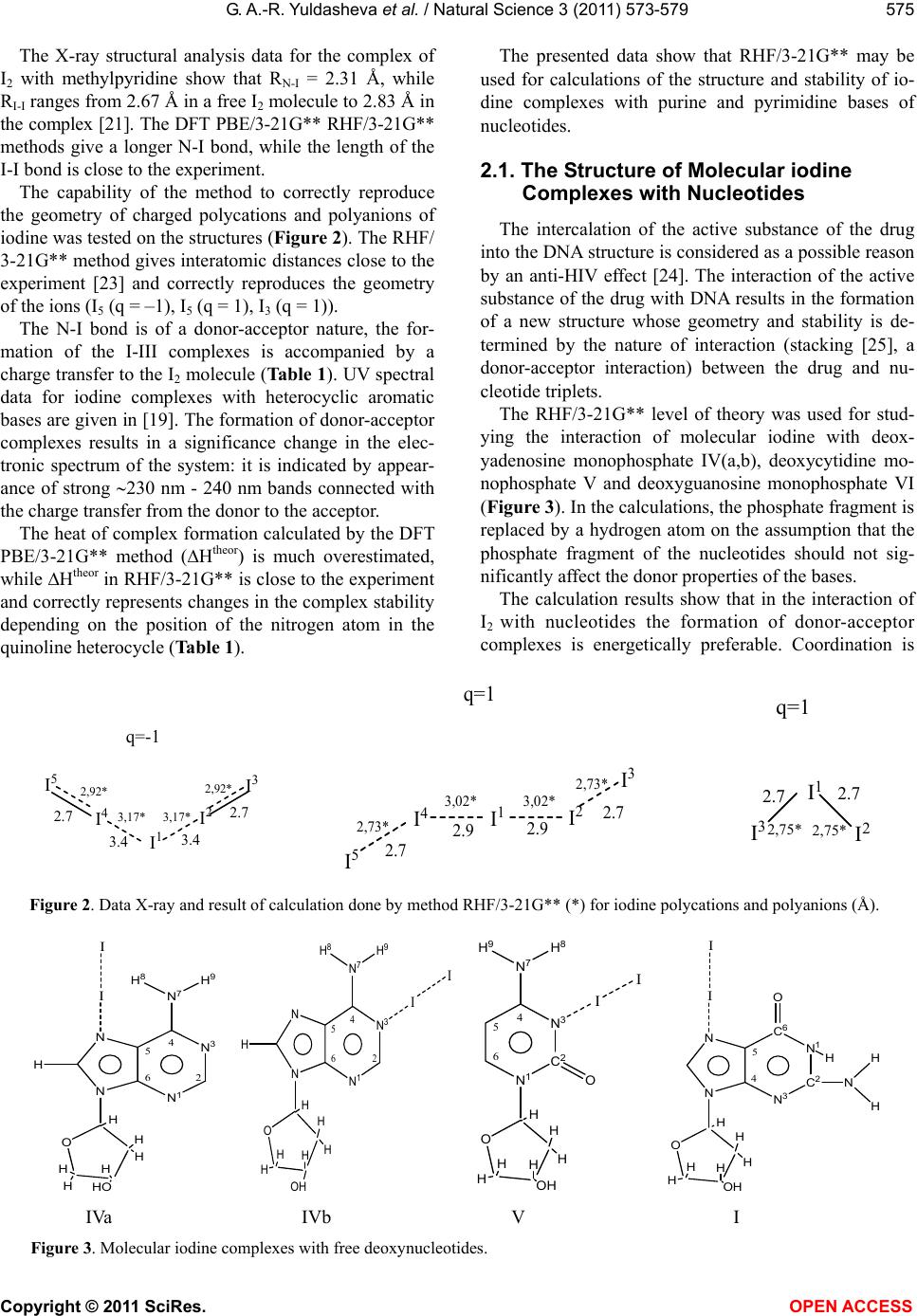 G. A.-R. Yuldasheva et al. / Natural Science 3 (2011) 573-579 Copyright © 2011 SciRes. OPEN ACCESS 575 The X-ray structural analysis data for the complex of I2 with methylpyridine show that RN-I = 2.31 Å, while RI-I ranges from 2.67 Å in a free I2 molecule to 2.83 Å in the complex [21]. The DFT PBE/3-21G** RHF/3-21G** methods give a longer N-I bond, while the length of the I-I bond is close to the experiment. The capability of the method to correctly reproduce the geometry of charged polycations and polyanions of iodine was tested on the structures (Figure 2). The RHF/ 3-21G** method gives interatomic distances close to the experiment [23] and correctly reproduces the geometry of the ions (I5 (q = –1), I5 (q = 1), I3 (q = 1)). The N-I bond is of a donor-acceptor nature, the for- mation of the I-III complexes is accompanied by a charge transfer to the I2 molecule (Table 1). UV spectral data for iodine complexes with heterocyclic aromatic bases are given in [19]. The formation of donor-acceptor complexes results in a significance change in the elec- tronic spectrum of the system: it is indicated by appear- ance of strong 230 nm - 240 nm bands connected with the charge transfer from the donor to the acceptor. The heat of complex formation calculated by the DFT PBE/3-21G** method (Htheor) is much overestimated, while Htheor in RHF/3-21G** is close to the experiment and correctly represents changes in the complex stability depending on the position of the nitrogen atom in the quinoline heterocycle (Table 1). The presented data show that RHF/3-21G** may be used for calculations of the structure and stability of io- dine complexes with purine and pyrimidine bases of nucleotides. 2.1. The Structure of Molecular iodine Complexes with Nucleotides The intercalation of the active substance of the drug into the DNA structure is considered as a possible reason by an anti-HIV effect [24]. The interaction of the active substance of the drug with DNA results in the formation of a new structure whose geometry and stability is de- termined by the nature of interaction (stacking [25], a donor-acceptor interaction) between the drug and nu- cleotide triplets. The RHF/3-21G** level of theory was used for stud- ying the interaction of molecular iodine with deox- yadenosine monophosphate IV(a,b), deoxycytidine mo- nophosphate V and deoxyguanosine monophosphate VI (Figure 3). In the calculations, the phosphate fragment is replaced by a hydrogen atom on the assumption that the phosphate fragment of the nucleotides should not sig- nificantly affect the donor properties of the bases. The calculation results show that in the interaction of I2 with nucleotides the formation of donor-acceptor complexes is energetically preferable. Coordination is I 1 I 2 I 4 I 3 I 5 2.7 3.4 3.4 2.7 q=-1 2,92* 3,17* 3,17* 2,92* I1I2 I4 I3 I5 2.7 q=1 2.7 2.9 2.9 2,73* 3,02* 2,73* 3,02* I 1 I 2 I 3 q=1 2.7 2.7 2,75* 2,75* Figure 2. Data X-ray and result of calculation done by method RHF/3-21G** (*) for iodine polycations and polyanions (Å). N 3 N 1 N N N 7 H H 9 H 8 I I O H H H HHO HH 4 5 62 N 3 N 1 N N N 7 H H 9 H 8 I I O H 4 5 62 H H HH OH H O H H H HH H C 2 N 3 N 1 N 7 H 8 H 9 I I 4 5 6 O OH O H H H HH H OH C 2 N 1 C 6 N 3 N N I I 4 5 H N H H O IVa IVb V I Figure 3. Molecular iodine complexes with free deoxynucleotides. 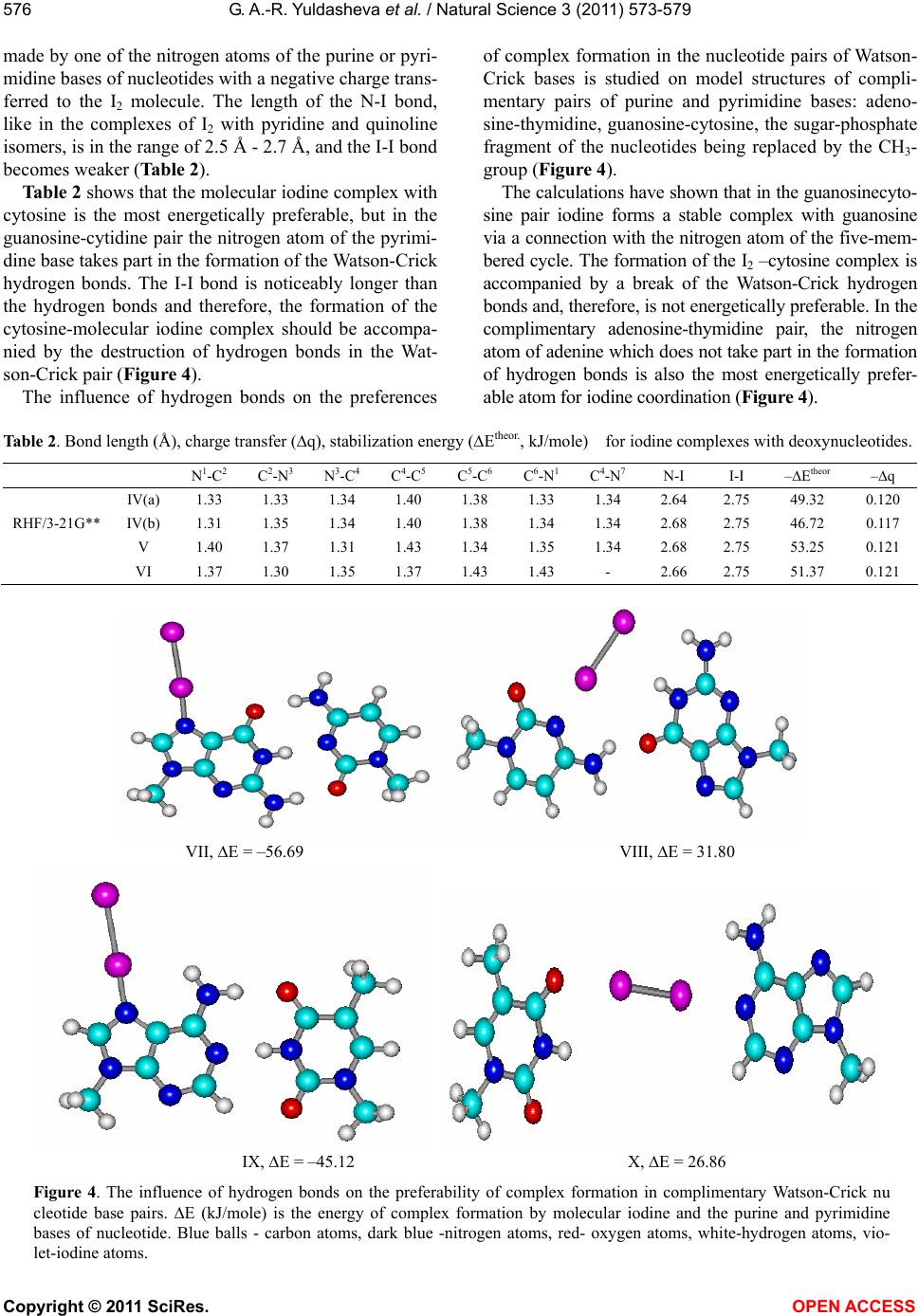 G. A.-R. Yuldasheva et al. / Natural Science 3 (2011) 573-579 Copyright © 2011 SciRes. OPEN ACCESS 576 made by one of the nitrogen atoms of the purine or pyri- midine bases of nucleotides with a negative charge trans- ferred to the I2 molecule. The length of the N-I bond, like in the complexes of I2 with pyridine and quinoline isomers, is in the range of 2.5 Å - 2.7 Å, and the I-I bond becomes weaker (Table 2). Table 2 shows that the molecular iodine complex with cytosine is the most energetically preferable, but in the guanosine-cytidine pair the nitrogen atom of the pyrimi- dine base takes part in the formation of the Watson-Crick hydrogen bonds. The I-I bond is noticeably longer than the hydrogen bonds and therefore, the formation of the cytosine-molecular iodine complex should be accompa- nied by the destruction of hydrogen bonds in the Wat- son-Crick pair (Figure 4). The influence of hydrogen bonds on the preferences of complex formation in the nucleotide pairs of Watson- Crick bases is studied on model structures of compli- mentary pairs of purine and pyrimidine bases: adeno- sine-thymidine, guanosine-cytosine, the sugar-phosphate fragment of the nucleotides being replaced by the СН3- group (Figure 4). The calculations have shown that in the guanosinecyto- sine pair iodine forms a stable complex with guanosine via a connection with the nitrogen atom of the five-mem- bered cycle. The formation of the I2 –cytosine complex is accompanied by a break of the Watson-Crick hydrogen bonds and, therefore, is not energetically preferable. In the complimentary adenosine-thymidine pair, the nitrogen atom of adenine which does not take part in the formation of hydrogen bonds is also the most energetically prefer- able atom for iodine coordination (Figure 4). Table 2. Bond length (Å), charge transfer (q), stabilization energy (Еtheor., kJ/mole) for iodine complexes with deoxynucleotides. N 1-C2 C2-N3 N3-C4 C4-C5 C5-C6 C6-N1 C4-N7 N-I I-I –Еtheor –q IV(a) 1.33 1.33 1.34 1.40 1.38 1.33 1.34 2.64 2.75 49.32 0.120 IV(b) 1.31 1.35 1.34 1.40 1.38 1.34 1.34 2.68 2.75 46.72 0.117 RHF/3-21G** V 1.40 1.37 1.31 1.43 1.34 1.35 1.34 2.68 2.75 53.25 0.121 VI 1.37 1.30 1.35 1.37 1.43 1.43 - 2.66 2.75 51.37 0.121 VII, E = –56.69 VIII, E = 31.80 IX, E = –45.12 X, E = 26.86 Figure 4. The influence of hydrogen bonds on the preferability of complex formation in complimentary Watson-Crick nu cleotide base pairs. Е (kJ/mole) is the energy of complex formation by molecular iodine and the purine and pyrimidine bases of nucleotide. Blue balls - carbon atoms, dark blue -nitrogen atoms, red- oxygen atoms, white-hydrogen atoms, vio- let-iodine atoms. 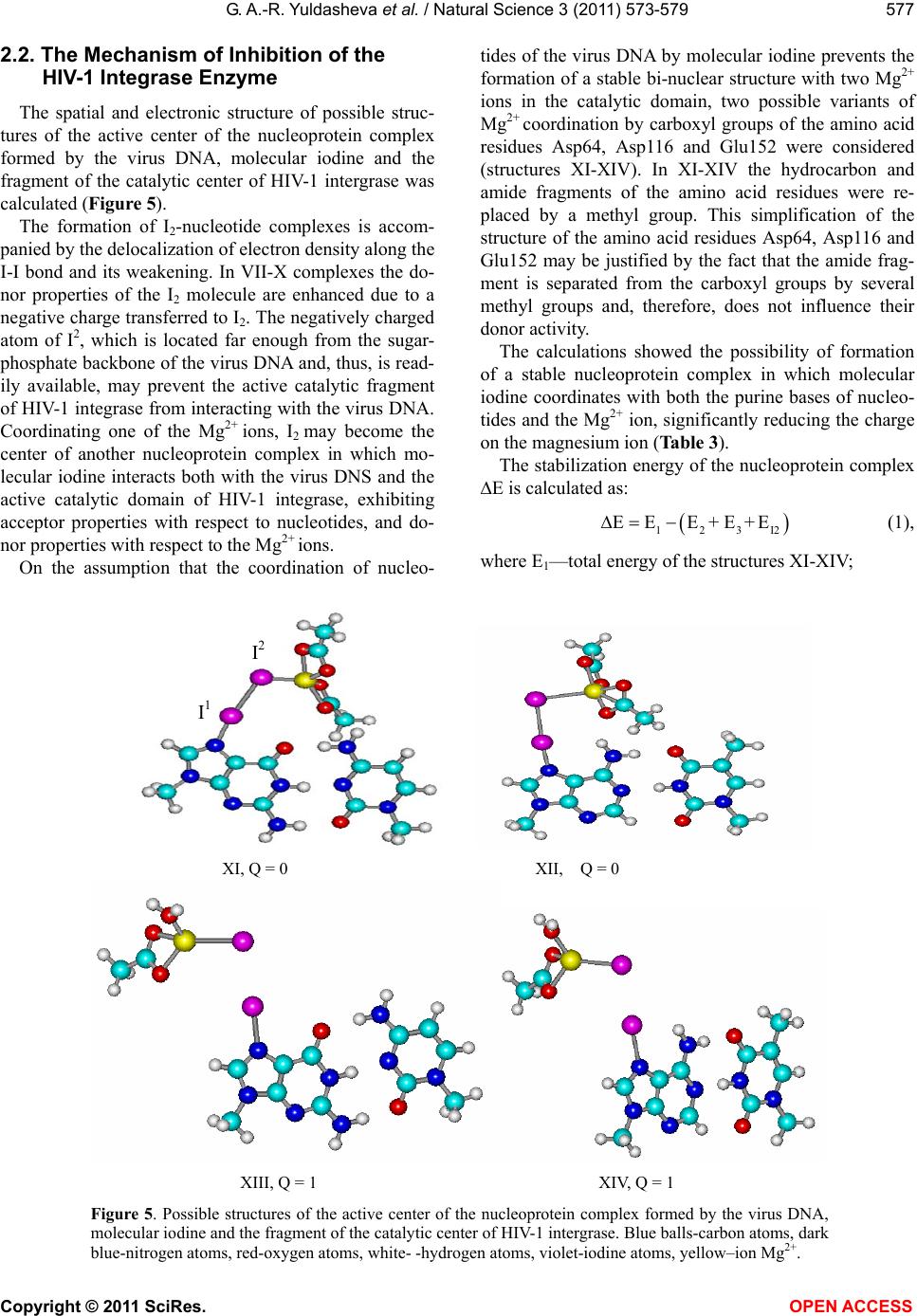 G. A.-R. Yuldasheva et al. / Natural Science 3 (2011) 573-579 Copyright © 2011 SciRes. OPEN ACCESS 577 2.2. The Mechanism of Inhibition of the HIV-1 Integrase Enzyme The spatial and electronic structure of possible struc- tures of the active center of the nucleoprotein complex formed by the virus DNA, molecular iodine and the fragment of the catalytic center of HIV-1 intergrase was calculated (Figure 5). The formation of I2-nucleotide complexes is accom- panied by the delocalization of electron density along the I-I bond and its weakening. In VII-X complexes the do- nor properties of the I2 molecule are enhanced due to a negative charge transferred to I2. The negatively charged atom of I2, which is located far enough from the sugar- phosphate backbone of the virus DNA and, thus, is read- ily available, may prevent the active catalytic fragment of HIV-1 integrase from interacting with the virus DNA. Coordinating one of the Mg2+ ions, I2 may become the center of another nucleoprotein complex in which mo- lecular iodine interacts both with the virus DNS and the active catalytic domain of HIV-1 integrase, exhibiting acceptor properties with respect to nucleotides, and do- nor properties with respect to the Mg2+ ions. On the assumption that the coordination of nucleo- tides of the virus DNA by molecular iodine prevents the formation of a stable bi-nuclear structure with two Mg2+ ions in the catalytic domain, two possible variants of Mg2+ coordination by carboxyl groups of the amino acid residues Asp64, Asp116 and Glu152 were considered (structures XI-XIV). In XI-XIV the hydrocarbon and amide fragments of the amino acid residues were re- placed by a methyl group. This simplification of the structure of the amino acid residues Asp64, Asp116 and Glu152 may be justified by the fact that the amide frag- ment is separated from the carboxyl groups by several methyl groups and, therefore, does not influence their donor activity. The calculations showed the possibility of formation of a stable nucleoprotein complex in which molecular iodine coordinates with both the purine bases of nucleo- tides and the Mg2+ ion, significantly reducing the charge on the magnesium ion (Table 3). The stabilization energy of the nucleoprotein complex Е is calculated as: 123I2 EE E+E+E (1), where E1—total energy of the structures XI-XIV; I 2 I 1 XI, Q = 0 XII, Q = 0 XIII, Q = 1 XIV, Q = 1 Figure 5. Possible structures of the active center of the nucleoprotein complex formed by the virus DNA, molecular iodine and the fragment of the catalytic center of HIV-1 intergrase. Blue balls-carbon atoms, dark blue-nitrogen atoms, red-oxygen atoms, white- -hydrogen atoms, violet-iodine atoms, yellow–ion Mg2+.  G. A.-R. Yuldasheva et al. / Natural Science 3 (2011) 573-579 Copyright © 2011 SciRes. OPEN ACCESS 578 Table 3. Stabilization energy (Е, kJ/mole), spatial (interatomic bond, Å) and electron characteristics of possible models of the active center of the nucleoprotein complex formed by the virus DNA, molecular iodine and the catalytic fragment of HIV-1 intergrase. XI XII XIII XIV –Е 110.07 108.35 318.69 259.70 N-I 2.42 2.53 2.15 2.13 I-I 2.81 2.79 3.12 3.21 Mg-I 2.95 2.95 2.69 2.68 Q (I1) 0.204 0.187 0.396 0.432 Q (I2) –0.235 –0.186 –0.333 –0.365 Q (Mg) 0.767 0.789 0.775 0.758 *) QMgCOOCOO = 0.885; QMgCOOH2O = 1.099. E2—total energy of Mg(COOCH3)2 for XI,XII and Mg(COOCH3)H 2O for XIII, XIV; E3—total energy of the Watson-Crick pair adeno- sine-thymidine in XI, XIII and guanosine-cytosine in XII, XIV; EI2—total energy of I2 molecule. In the structures XIII, XVI the I-I bond is broken and RN-I indicates that there is a new N-I bond in the purine bases of adenosine and guanosine (Table 3). Among the recently developed drugs inhibiting the activity of HIV-1 integrase there are those whose inhib- iting activity is connected with the formation of coordi- nation bonds with two Mg2+ ions [12,26] of the catalytic fragment of HIV-1 integrase. The calculations revealed that the molecular iodine contained in iodine polymer complexes could be referred to this class of compounds, but, unlike the known inhibitors of HIV-1 integrase, I2 also changes the structure of the virus DNA. 3. CONCLUSIONS The computation results suggest that molecular iodine in drugs containing molecular iodine complexes may be referred to compounds inhibiting the catalytic center of the HIV-1 integrase enzyme. It is shown that molecular iodine prevents the forma- tion of PIC and inhibits the HIV-1 intergrase enzyme inside the nucleoprotein complex where I2 interacts both with the virus DNA and the active center of the catalytic domain of the HIV-1 intergrase exhibiting acceptor properties with respect to the nucleotides of the virus DNA and donor properties with respect to Mg2+ ions. The breaking of the I-I bond and the formation of a new N-I bond in the purine bases of adenosine and gua- nosine may be observed in the structure of the nucleo- protein complex. REFERENCES [1] Delelis, O., Carayon, K., Saïb, A., Deprez, E. and Mous- cadet, J.F. (2008) Integrase and integration: Biochemical activities of HIV-1 integrase. Retrovirology, 5. [2] Long, M.C., King, J.R. and Acosta, E.P. (2009) Pharma- cologic aspects of new antiretroviral drugs. Current HIV/ AIDS Reports, 6, 43-50. doi:10.1007/s11904-009-0007-y [3] Emmelkamp, J.M. and Rockstroh, J.K. (2008) Maraviroc risks and benefits: a review of the clinical literature. Ex- pert Opinion on Drug Safety, 7, 559-569. doi:10.1517/14740338.7.5.559 [4] Borrás-Blasco, J., Navarro-Ruiz, A., Borrás, C. and Cas- terá E. (2008) Adverse cutaneous reactions associated with the newest antiretroviral drugs in patients with hu- man immunodeficiency virus infection. Journal of An- timicrobial Chemotherapy, 62, 879-888. doi:10.1093/jac/dkn292 [5] Sen, S., Mathur, A.G., Gupta, R.M., Kapila, K. and Cho- pra, G.S. (2008) Investigational antiretroviral drugs. Re- cent Patents on Anti-Infective Drug Discovery, 3, 199- 205. doi:10.2174/157489108786242387 [6] Ciuffi, A. (2008) Mechanisms governing lentivirus inte- gration site selection. Current Gene Therapy, 8, 419-429. doi:10.2174/156652308786848021 [7] Farnet, C.M. and Haseltine, W.A. (1990) Integration of human immunodeficiency virus type 1 DNA in vitro. Proceedings of the National Academy of Sciences, 87, 4164-4168. doi:10.1073/pnas.87.11.4164 [8] Podtelezhnikov, A.A., Gao, K., Bushman, F.D. and Mc- Cammon, J.A. (2003) Modeling HIV-1 integrase com- plexes based on their hydrodynamic properties. Bio- polymers, 68, 110-120. doi:10.1002/bip.10217 [9] Dyda, F., Hickman, A.B., Jenkins, T.M., Engelman, A., Craigie, R. and Davies, D.R. (1994) Crystal structure of catalytic domain of HIV-1 integrase similarity to other polynucleotidyl transferases. Science, 266, 1981-1986. doi:10.1126/science.7801124 [10] Goldgur, Y., Dyda, F., Hickman, A.B., Jenkins, T.M., Craigie, R. and Davies, D.R. (1998) Three new structures of the core domain of HIV-1 integrase: an active site that binds magnesium. Proceedings of the National Academy of Sciences, 95, 9150-9154. doi:10.1073/pnas.95.16.9150 [11] Sawaya, M.R., Prasad, R., Wilson, S.H., Kraut, J. and Pelletier, H. (1997) Crystal structures of human DNA polymerase beta complexed with gapped and nicked DN- A Evidence for an induced fit mechanism. Biochemistry, 36, 11205-11215. doi:10.1021/bi9703812 [12] Chen, X., Tsiang, M., Yu, F., Hung, M., Jones, G.S.,  G. A.-R. Yuldasheva et al. / Natural Science 3 (2011) 573-579 Copyright © 2011 SciRes. OPEN ACCESS 579 Zeynalzadegan, A., Qi, X., Jin, H., Kim, C.U., Swami- nathan, S. and Chen, J.M. (2008) Modeling, analysis and validation of a novel HIV integrase structure provide in- sights into the binding modes of potent integrase inhibi- tors. Journal of Molecular Biology, 380, 504-519. doi:10.1016/j.jmb.2008.04.054 [13] Farnet, C.M., Wang, B., Lipford, J.R. and Bushman, F.D. (1996) Differential inhibition of HIV-1 preintegration complexes and purified integrase protein by small mole- cules. Proceedings of the National Academy of Sciences, 93, 9742-9747. doi:10.1073/pnas.93.18.9742 [14] International Application (2000) Ilyin Alexandr, Gab- rielyan Emil, Mkhitaryan, Levon Antiviral and antibacte- rial pharmaceutical preparation “Armenicum” And its use for treatment of infectious diseases. Patent No.: PCT/AM 2000/000002. [15] Shikani, A.H., St Clair, M. and Domb, A. (1996) Poly- mer-iodine inactivation of the human immunodeficiency virus. Journal of the American College of Surgeons, 183, 195-200 [16] Ilyin A.I. and Kulmanov, M.E. (2004) Bactericide and viricidal pharmaceutical preparation for prophylaxis and treatment mono and mixtinfections, manner of prophy- laxis o/or treatment viral, bacterium and mixtinfection. Patent No.: 15116. [17] Davtyan, T.K., Mkhitaryan, L.M. and Gabrielyan, E.S. (2009) Desing of iodine-lithium- -dextrin liguid crystal with potent antimicrobaial and anti-inflammatory proper- ties. Current Pharmaceutical Design, 15, 1172-1186. doi:10.2174/138161209787846829 [18] Mkhitaryan, L.M., Davtyan, T.K., Gabrielyan, E.S. and Gevorkyan, L.A. (2007) Anti-HIV and anti-inflammatory action of iodine-lithium-α-dextrin is accompanied by the improved quality of life in AIDS patients. International Journal of Biotechnology, 9, 301-317. doi:10.1504/IJBT.2007.014252 [19] Krishna, V.G. and Bhowmik, B.B. (1968) Charge-tran- sfer intensities of iodine complexes with N-heterocyclics. Journal of the American Chemical Society, 90, 1700- 1705. doi:10.1021/ja01009a004 [20] McKinney, W.J. and Popov, A.I. (1969) Studies on the chemistry of halogen and of polyhalides. XXX. The in- fluence of solvent properties on formation of pyridine- -iodine charge-transfer complexes. Journal of the Amer- ican Chemical Society, 91, 5215-5218. doi:10.1021/ja01047a005 [21] Hassel, O., Romming, C. and Tufte, T. (1961) Crystal structure of the 11 addition compound formed by 4-pi- coline and iodine. Acta Chemica Scandinavica, 15, 967- 974. doi:10.3891/acta.chem.scand.15-0967 [22] Perdew, J.P., Burke, K. and Ernzerhof, M. (1996) Gener- alize gradient approximation made simple. Physical Re- view Letters, 77, 3865-3868. doi:10.1103/PhysRevLett.77.3865 [23] Tretyakov, U.D. (2004) Inorganic chemistry in Russian. Academia, Moscow. [24] Sun, Z., Ma, Z., Zhang, W., Wang, X., Fan, C. and Li, G. (2004) Electrochemical investigations of baicalin and DNA-baicalin interactions. Analytical and Bioanalytical Chemistry, 379, 283-286. doi:10.1007/s00216-004-2542-0 [25] Reha, D., Kabelac, M., Ryjacek, F., Sponer J., Sponer, J.E., Elstner, M., Suhai, S. and Hobza, P. (2002) Interca- lators. 1. Nature of stacking interaction between interca- lators (ethidium, daunomycin, ellipticine and 4,6-dia- minide-2-phenylindole) and DNA base pairs. Ab initio quantum chemical, density functional theory and empiri- cal potential study. Journal of the American Chemical Society, 124, 3366-3376. doi:10.1021/ja011490d [26] Globler, J.A., Stillmock, K., Hu, B., Witmer, M., Felock, P., Espeseth, A.S., Wolfe, A., Egbertson, M., Bourgeois, M., Melamed, J., Wai, J.S., Young, S., Vacca, J. and Ha- zuda, D.J. (2002) Diketo acid inhibitor mechanism and HIV-1 integrase Implications for metal binding in the active site of phosphotransferase enzymes. Proceedings of the National Academy of Sciences, 99, 6661-6666. doi:10.1073/pnas.092056199
|