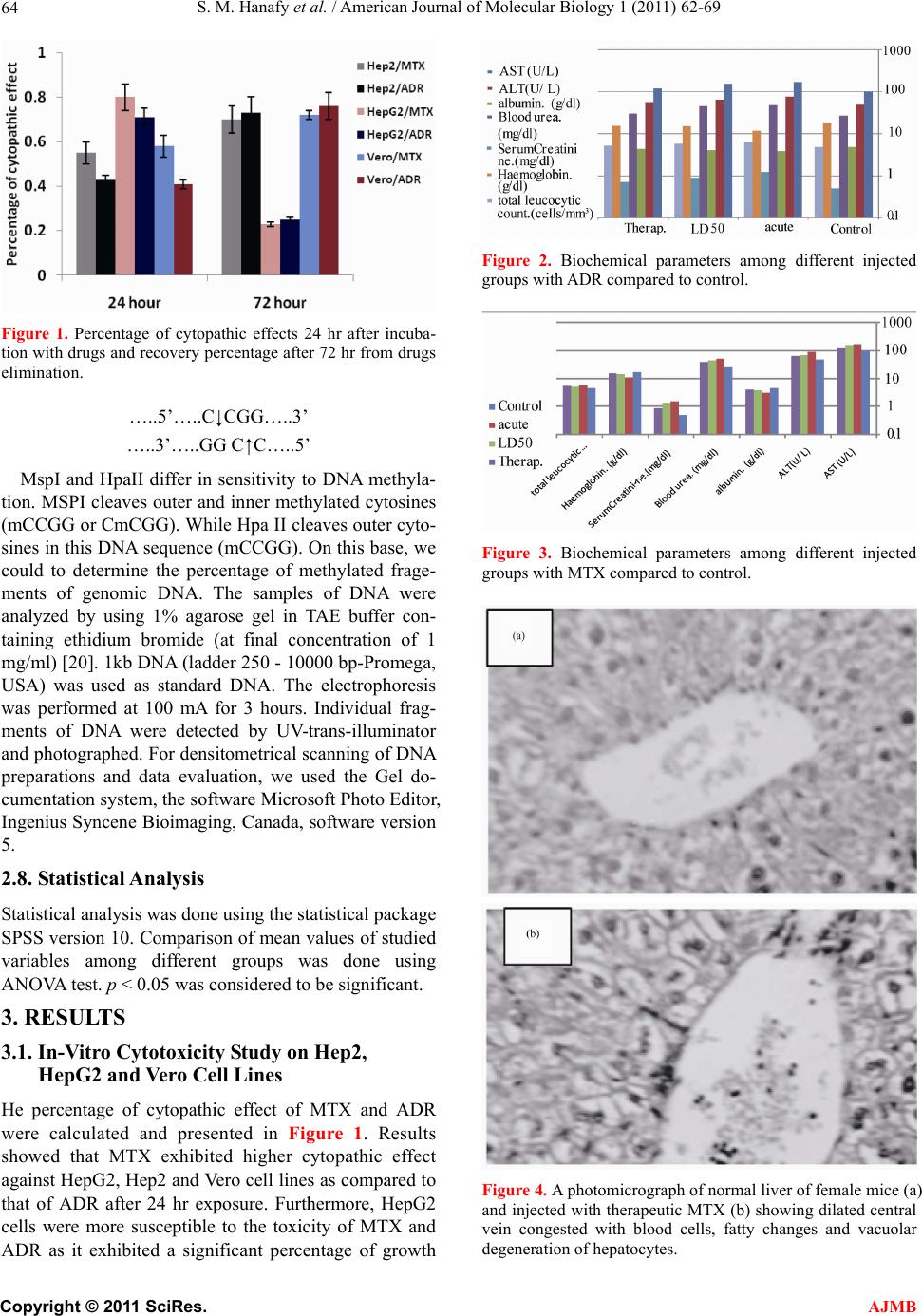
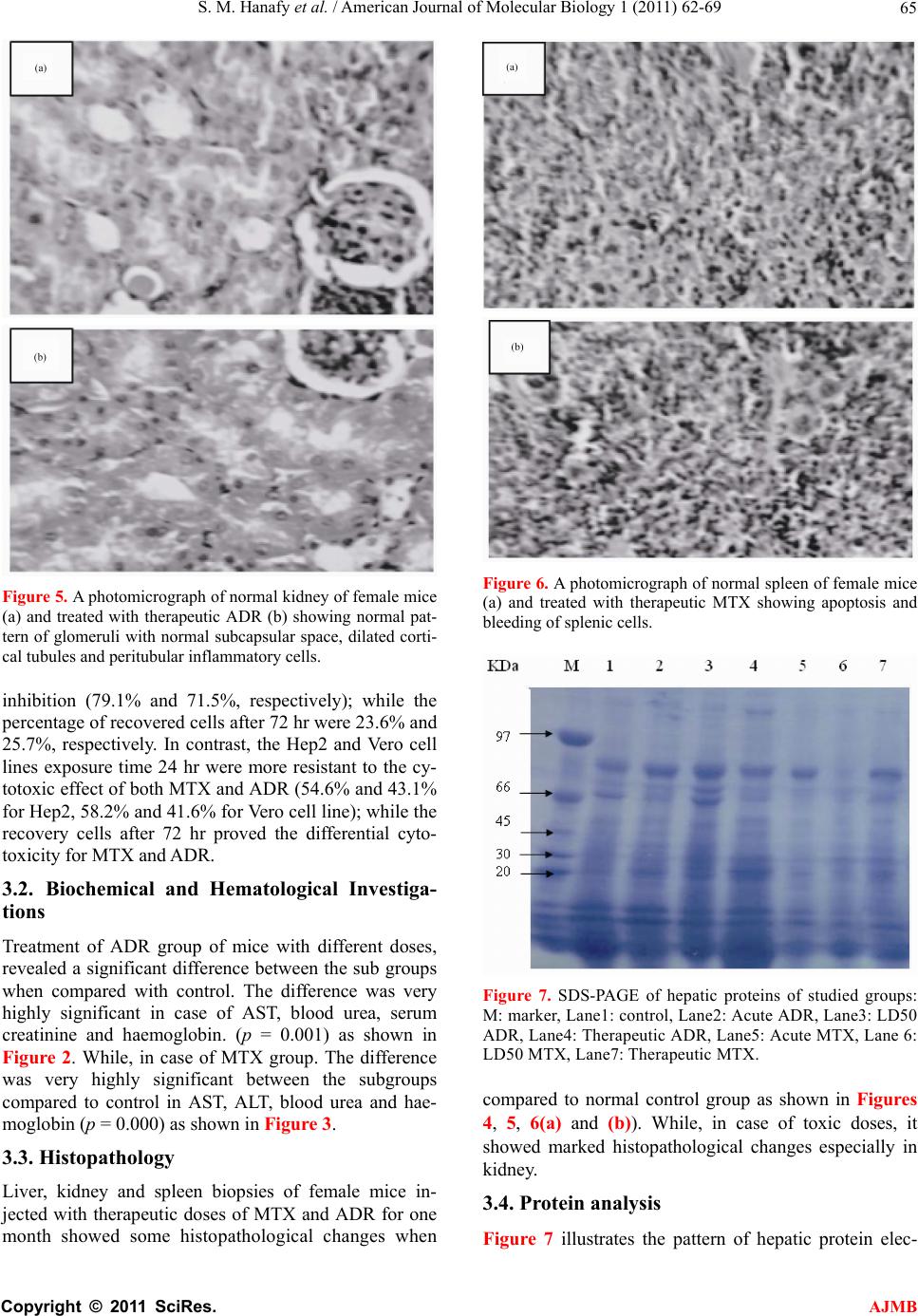
S. M. Hanafy et al. / American Journal of Molecular Biology 1 (2011) 62-69
68
es from treatment of tumor cells with the adriamycine.
DNA methylation level from CpG islands linked to
twenty eight genes whose expression levels had been
shown to contribute with the resistance against DNA
double strand break induced drugs. These data were
supported in some way to our data in documenting DNA
methylation by different doses of doxorubicin drug in
non-tumor female mice [28] assessed the CpG methyla-
tion aberrations induced by pixantrone and doxorubicin.
A characteristic that may determine the most cancer
types to specific drug treatments and is a marker of drug
sensitivity. Moreover, Winter-Vann, et al. [29] suggested
that MTX has an additional mechanism of action besides
it is a potent product inhibitor of cellular methyltrans-
ferases. It is also having an inhibitiory effect on Ras sig-
naling that regulates cell growth and differentiation. Be-
cause carboxyl methylation of Ras is important for
proper plasma membrane localization and function, they
reported that after MTX treatment of DKOB8 cells, Ras
methylation is decreased by almost 90% and subse-
quently inhibition of carboxyl methylation.
5. CONCLUSIONS
DNA methylation is influenced by anticancer drugs
(MTX and ADR) and this influence was in a dose-
dependent manner; as they exhibited reduction in DNA
methylation with varying degrees in liver genomic DNA.
Some specific protein bands may be considered as a po-
tential markers associated with high doses of anticancer
drugs. Treatment with DNA methylation inhibitors may
reactivate epigenetically silenced genes and has been
shown to restore normal gene function. Further studies
are recommended to characterize the protein fragments
associated with anticancer drugs treatment.
REFERENCES
[1] Shkreta, L., Froehlich, U., Paquet, E.R., Toutant, J., Elela,
S.A., Chabot, B. (2008) Anticancer drugs affect the
alternative splicing of Bcl-x and other human apoptotic
genes. Molecular Cancer Therapeutics, 7, 1398-1409.
doi:10.1158/1535-7163.MCT-08-0192
[2] Laque-Ruperez, E., Ruiz-Gomez, M.J., de la Pena, L.,
Gil, L. and Martinez-Morillo, M. (2003) Methotrexate
cytotoxicity on MCF-7 breast cancer cells is not altered
by exposure to 25 Hz, 1.5 mT magnetic field and iron (III)
chloride hexahydrate. Bioelectrochemistry, 60, 81-86.
doi:10.1016/S1567-5394(03)00054-9
[3] Senter, P.D., Vrudhula,V.M., Wallace, P.M., Somervil-
le, J.A., Wang, I. and Lowe, D.A. (1995) Sulfated eto-
poside and nitrogen mustard prodrugs and their activa-
tion by streptomyces arylsulfatase. Drug Delivery, 2,
110-116. doi:10.3109/10717549509031358
[4] Cronstein, B.N., Naime, D. and Ostad, E. (1993) The
anti-inflammatory mechanism of methotrexate: Increased
adenosine release at inflamed sites diminishes leukocytes
accumulation in an in vivo model of inflammation.
Journal of Clinical Investigation, 92, 2675-2682.
doi:10.1172/JCI116884
[5] Rosenthal, G.J., Weigand, G.W. and Germolec, D.R.
(1988) Suppression of B cell functions by methotrexate
and trimethotrexate. Evidence for inhibition of purine
biosynthesis as a major mechanism of action. The Jour-
nal of Immunology, 141, 410-416.
[6] Kosuke, Y., Kenichi, S., Motofumi, S., Atsuhiro, T.,
Sakiko, O., Hitomi, U., Shinichi, O., Jun, W., Ryo, N.,
Daisuke, O., Yasushi, S. and Hirofumi, M. (2005) Metho-
trexate Prevents Renal Injury in Experimental Diabetic
Rats via Anti-Inflammatory Actions. Journal of the
American Society of Nephrology, 16, 3326-3338.
doi:10.1681/ASN.2004111011
[7] Mazzotta, P., Kwasnicka, A. and Kutas, G.J. (2001)
Cancer Chemotherapy: The role of pharmacological
agents in the management of haematological malignan-
cies. University of Toronto Medical Journal, 79, 38-45.
[8] Hendrich, B. and Bird, A. (1998) Identification and
charcterization of family of mammalian methyl-CpG
binding proteins. Molecular and Cellular Biology, 18,
6538-6547.
[9] Antequera, F. (2003) Structure, function and evolution of
CpG island promoters. Cellular and Molecular Life
Sciences, 60, 1647-1658.
doi:10.1007/s00018-003-3088-6
[10] Oakes, C.C., Smiraglia, D.J., Plass, C., Trasler, J.M.,
Robaire, B. (2003) Aging results in hypermethylation of
ribosomal DNA in sperm and liver of female rats.
Proceedings of the National Academy of Sciences of the
Uniited States of Americia, 100, 1775-1789.
doi:10.1073/pnas.0437971100
[11] Freshney, R.I. (2005) Culture of Animal Cells, a Manual
of Basic Technique. 5th Edition, John Wiley & Sons,
Hoboken. doi:10.1002/9780471747598
[12] Reitman, S. and Frankel, S. (1975) A colorimetric me-
thod for the determination of serum glutamic oxalacetic
and glutamic pyruvic transaminases. American Journal
of Clinical Pathology, 28, 56-63.
[13] Doumas, B., Watson, W. and Biggs, H. (1971) Albumin
standards and the measurement of serum albumin with
bromcresol green. Clinica Chimica Acta, 1, 87-96.
doi:10.1016/0009-8981(71)90365-2
[14] Young, D.S. (2001) Effects of diseases on clinical lab.
tests. 4th Edition, American Association for Clinical
Chemistry, Inc., Washington DC.
[15] Bartels, H., Bohmer, M. and Heierli, C. (1972) Serum
creatinine determination without protein precipitation.
Clinica Chimica Acta, 37, 193-197.
doi:10.1016/0009-8981(72)90432-9
[16] Tiez, N.W. (1976) Fundamentals of Clinical Chemistry.
W.B. Saunders Co., Philadelphia.
[17] Portmann, B., Talbol, I.D. and Day, D.W. (1975) Histo-
pathological changes in the liver following paracetamol
overdose: Correlation with clinical and biochemical
parameters. The Journal of Pathology, 117, 169-181.
doi:10.1002/path.1711170307
[18] Laemmli, U.K. (1970) Cleavage of structure proteins
during assembly of head bacteriophage T4. Nature, 227,
680-685. doi:10.1038/227680a0
[19] Bradford, M.M. (1976) A rapid and sensitive for the
C
opyright © 2011 SciRes. AJMB