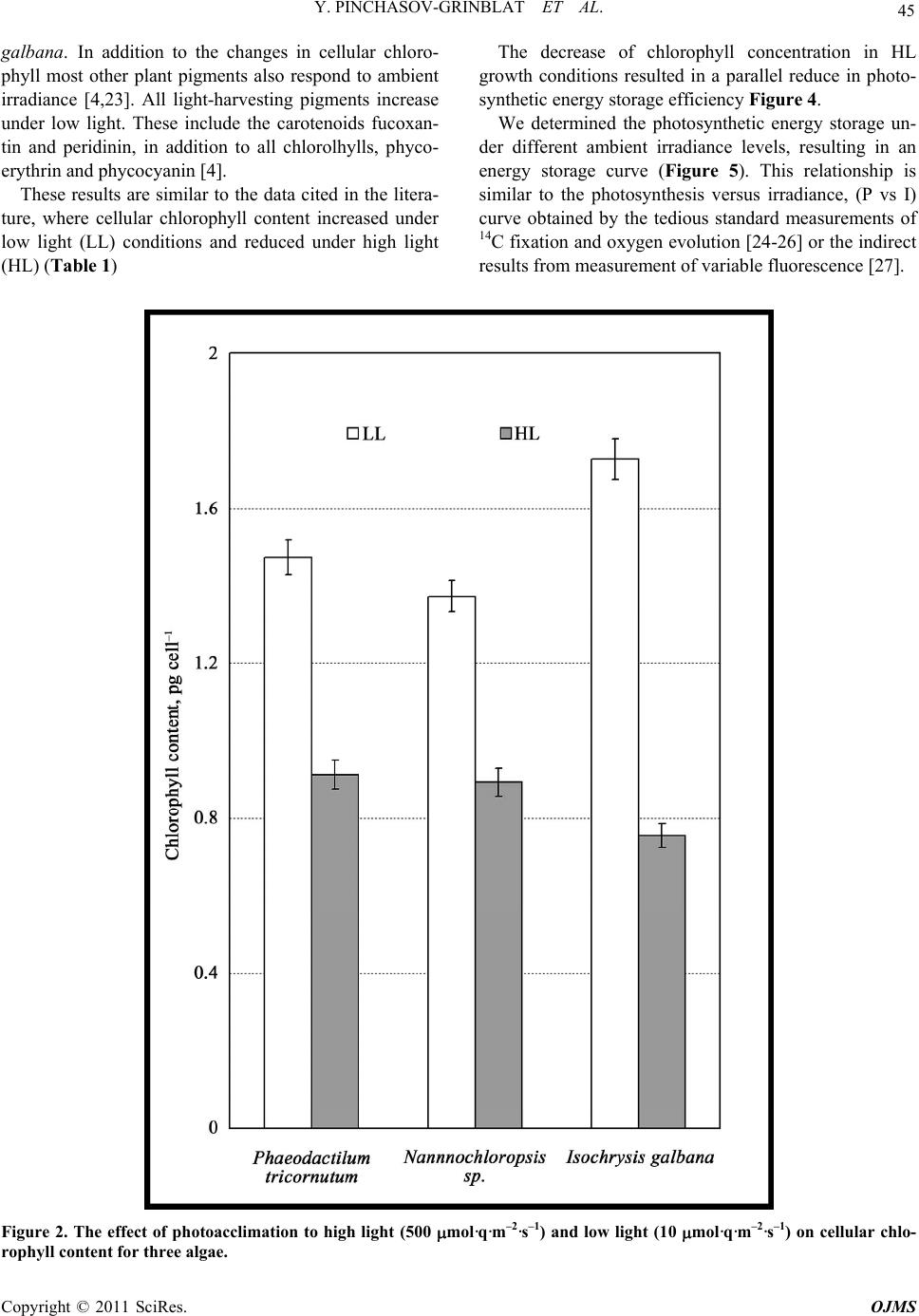
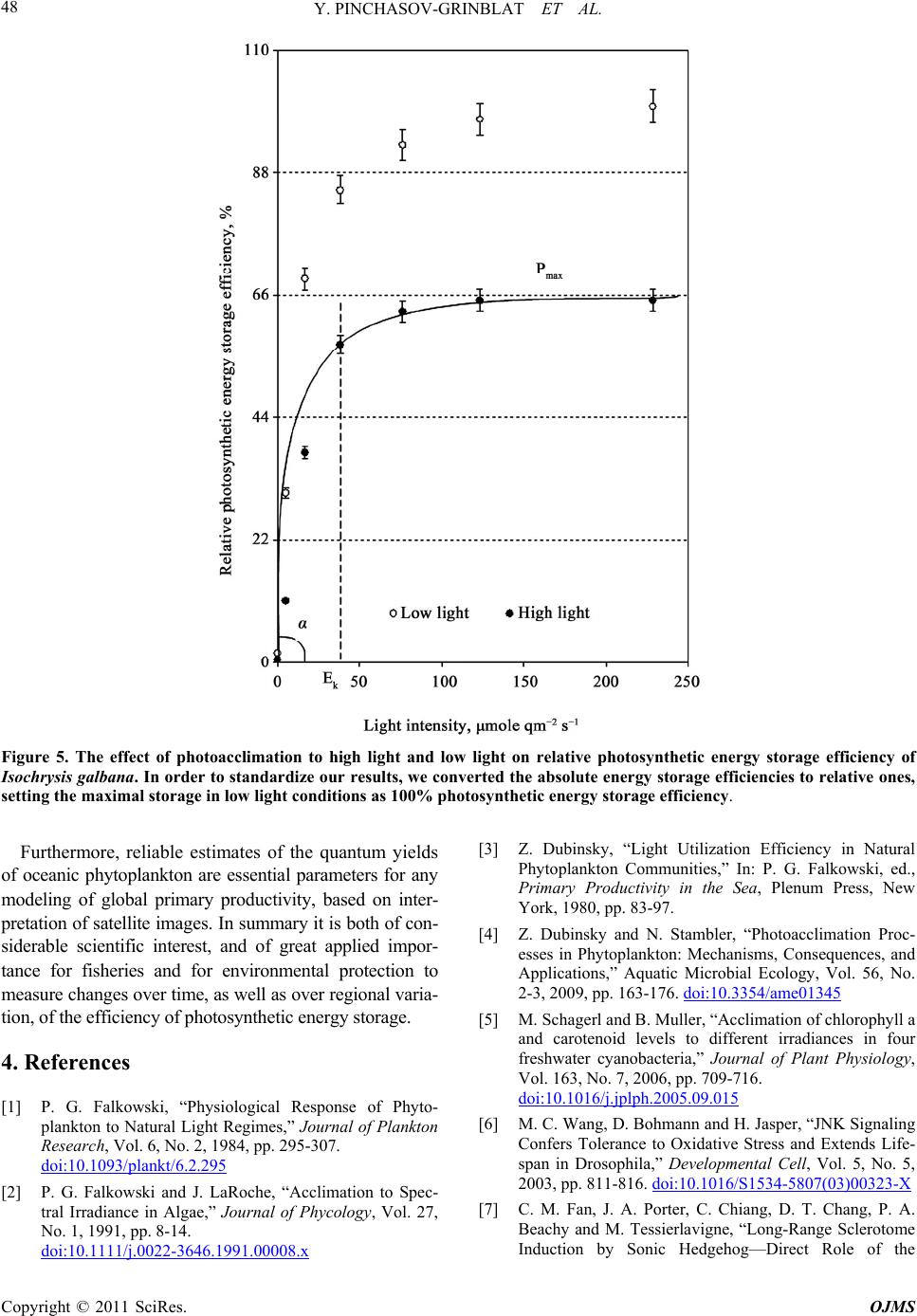
Y. PINCHASOV-GRINBLAT ET AL.
49
Amino-Terminal Cleavage Product and Modulation by
the Cyclic-AMP Signaling Pathway,” Cell, Vol. 81, No. 3,
1995, pp. 457-465. doi:10.1016/0092-8674(95)90398-4
[8] Z. Dubinsky, R.Matsukawa and I. Karube, “Photobi-
ological Aspects of Algal Mass Culture,” Journal of Ma-
rine Biotechnology, Vol. 2, 1995, pp. 61-65.
[9] Z. Dubinsky, P. G. Falkowski and K. Wyman, “Light
Harvesting and Utilization in Phytoplankton,” Plant &
Cell Physiology, Vol. 27, No. 7, 1986, pp. 1335-1350.
[10] T. Berner, K. Wyman, Z. Dubinsky and P. G. Falkowski,
“Photoadaptation and the “Package Effect” in Dunaliella
Tertiolecta (Chlorophyceace),” Journal of Phycology,
Vol. 25, No. 1, 1989, pp. 70-81.
doi:10.1111/j.0022-3646.1989.00070.x
[11] A. F. Post, Z. Dubinsky, K. Wuman and P. G. Falkowski,
“Kinetics of Light-Intensity Adaptation in a Marine
Planktonic Diatom,” Marine Biology, Vol. 83, No. 3,
1984, pp. 231-238. doi:10.1007/BF00397454
[12] S. Malkin and D. Cahen, “Photoacoustic Spectroscopy
and Radiant Energy Conversion: Theory of the Effect
with Special Emphasis on Photosynthesis,” Photochemis-
try and Photobiology, Vol. 29, No. 4, 1979, pp. 803-813.
doi:10.1111/j.1751-1097.1979.tb07770.x
[13] O. Canaani, S. Malkin and D. Mauzerall, “Pulsed Photo-
acoustic Detection of Flash-Induced Oxygen Evolution
from Intact Leaves and Its Oscillations,” Proceedings of
National Academy of Science of USA, Vol. 85, No. 13,
1988. pp. 4725-4729. doi:10.1073/pnas.85.13.4725
[14] S. Malkin, S. K. Herbert and D. C. Fork, “Light Distribu-
tion, Transfer and Utilization in the Marine Red Alga
Porphyra Perforata from Photoacoustic Energy-Storage
Measurements,” Biochimica et Biophysica Acta, Vol.
1016, No. 2, 1990, pp. 177-189.
doi:10.1016/0005-2728(90)90056-A
[15] Z. Dubinsky, J. Feitelson and D. C. Mauzerall, “Listening
to Phytoplankton: Measuring Biomass and Photosynthe-
sis by Photoacoustics,” Journal of Phycology, Vol. 34,
No. 5, 1998, pp. 888-892.
doi:10.1046/j.1529-8817.1998.340888.x
[16] D. C. Mauzerall, J. Feitelson and Z. Dubinsky, “Dis-
criminating between Phytoplankton Taxa by Photoacous-
tics,” Israel Journal of Chemistry, Vol. 38, No. 3, 1998,
pp. 257-260.
[17] Y. Pinchasov, D. Kotlyarevsky, Z. Dubinsky, D. C. Mau-
zerall and J. Feitelson, “Photoacoustic as a Diagnostic
Tool for Probing the Physiological Status of Phytoplank-
ton,” Israel Journal of Plant Sciences, Vol. 53, No. 1,
2005, pp. 1-10. doi:10.1560/4DVV-JT78-G7VW-QTFM
[18] R. R. L. Guillard and J. H. Ryther, “Studies of Marine
Planktonic Diatoms. I. Cyclotella Nana Hustedt and
Detonula Confervacea (Cleve) Gran,” Canadian Journal
of Microbiology, Vol. 8, No. 2, 1962, pp. 229-239.
doi:10.1139/m62-029
[19] S. W. Jeffrey and G. F. Humphrey, “New Spectropho-
tometric Equation for Determining Chlorophyll A, B, C1
and C2 in Higher Plants, Algae and Natural Phytoplank-
ton,” Biochemie Physiolgie der Pflanzen, Vol. 167, 1975,
pp. 194-204.
[20] P. G. Falkowski, “Light-shade Adaptation in Marine
Phytoplankton Primary Productivity in the Sea,” Plenum
Press, New York, 1980, pp. 99-119.
[21] P. G. Falkowski, “Physiological Response of Phyto-
plankton to Natural Light Regimes,” Journal of Plankton
Research, Vol. 6, No. 2, 1984, pp. 295-307.
doi:10.1093/plankt/6.2.295
[22] M. Ritz, J. C. Thomas, A. Spilar and A. L. Etienne, “Ki-
netics of Photoacclimation in Response to a Shift to High
Light of the Red Alga Rhodella violacea Adapted to Low
Irradiance,” Plant Physiology, Vol. 123, No. 4, 2000, pp.
1415-1426. doi:10.1104/pp.123.4.1415
[23] H. L. MacIntyre, T. M. Kana and J. R. Geider, “The Ef-
fect of Water Motion on the Short-Term Rates of Photo-
synthesis by Marine Phytoplankton,” Trends in Plant
Science, 2000, Vol. 5, No. 1, pp. 12-17.
doi:10.1016/S1360-1385(99)01504-6
[24] E. Steemann-Nielsen, “The Use of Radioactive Carbon
(14C) for Measuring Organic Production in the Sea,”
Journal du Conseil, Perm International pour
l’Exploration de la Mer, Vol. 18, 1952, pp. 117-140.
[25] T. Berner, Z. Dubinsky, F. Schanz, J. U. Grobbelaar, U.
Uehlinger and P. Falkowski, “The Measurement of Pri-
mary Productivity in a High Rate Oxidation Pond,” Jour-
nal of Plankton Research, Vol. 8, No. 4, 1986, pp.
659-672. doi:10.1093/plankt/8.4.659
[26] Z. Dubinsky, P. G. Falkowski, A. F. Post and U. M. van
Hes, “A System for Measuring Phytoplankton Photosyn-
thesis in a Defined Light Field with an Oxygen Elec-
trode,” Journal of Plankton Research, Vol. 9, No. 4, 1987,
pp. 607-612. doi:10.1093/plankt/9.4.607
[27] P. G. Falkowski, K. Wyman, A. Ley and D. Mauzerall,
“Relationship of Steady State Photosynthesis to Fluores-
cence in Eucaryotic Algae,” Biochimica et Biophysica
Acta, Vol. 849, No. 2, 1986, pp. 183-192.
doi:10.1016/0005-2728(86)90024-1
[28] R. Iglesias-Prieto and R. K. Trench, “Acclimation and
Adaptation to Irradiance in Symbiotic Dinoflagellates. I.
Responses of the Photosynthetic Unit to Changes in Pho-
ton Flux Density,” Marine Ecology Progress Series, Vol.
113, 1994, pp. 163-175. doi:10.3354/meps113163
Copyright © 2011 SciRes. OJMS