Natural Science
Vol.4 No.8(2012), Article ID:21996,7 pages DOI:10.4236/ns.2012.48074
Isolation and molecular identification of Ascomycetes in sediments and waters of the Gulf of Aqaba, Red Sea
![]()
1Department of Biological Sciences, The University of Jordan, Amman, Jordan; *Corresponding Author: b.jaber@ju.edu.jo
2Department of Marine Biology, Faculty of Marine Sciences, The University of Jordan/Aqaba Branch, Aqaba, Jordan
Received 25 June 2012; revised 28 July 2012; accepted 10 August 2012
Keywords: Marine fungi; Ascomycete; 18S ssu rRNA; Gulf of Aqaba; Red Sea
ABSTRACT
Using molecular approach, we aimed to identify fungal phylotypes that exist in the Gulf of Aqaba, Red sea. Several samples were taken from sediments and seawater of three locations along 26 kilometers at 5 m depth. 18S small subunit rRNA gene was targeted for PCR amplification and sequencing. Partial sequences introduced as query in BLASTN phylogenetic analysis revealed 100% identity with Ascomycota, namely, Aspergillus sp. Penicillium sp. and its closely related Eupenicillium sp. The top scorer species in this analysis were Aspergillus sydowii, Aspergillus wentii, Aspergillus flocculosus, Penicillium expansum and Eupenicillium javanicum with 98% - 100% identity. Phylogenetic analyses demonstrates close relatedness among isolated fungi and potential association with Ascomycetes. This study reports a new geographical location in which facultative marine Ascomycetes exist in, and sheds some light on fungal diversity in Gulf of Aqaba.
1. INTRODUCTION
Fungi are found ubiquitously in the environment. They execute a wide range of important ecological functions especially ones associated with decomposition of organic substrates in both terrestrial as well as marine environments, and occasionally in extreme conditions [1-5]. The magnitude of fungal biodiversity was estimated to be 1.5 million species [6,7]. However, some estimation exceeded this number to 5.1 M [8]. Among the estimated 1.5 million, only 5% - 10% has been formally described [3,8]. Fungi that live in the sea are defined as obligate marine or facultative. Obligate marine fungi are those that grow and sporulate exclusively in a marine or estuarine habitat and are permanently or intermittently submerged in water. Whereas, facultative marine fungi are those that normally occupy freshwater habitats or terrestrial surroundings but are able to grow (and possibly to sporulate) in the marine environment [9,10]. Currently there are 444 known recognized species of obligate and facultative marine fungi. Ascomycota represented by 360 species whereas Basidiomycota are represented by 10 species and 74 species of mitosporic fungi [11]. Facultative marine fungi can be transferred to seawater by wind, rain or runoff soil. Some facultative marine fungi were able to evolve adaptation to marine environment and eventually became obligate marine [12].
Many reasons have posed the need to examine the extent of marine biodiversity of fungi. For example, the emergence marine life diseases to corals, sponges, sea fan corals and fish [3,13-15]. Some marine fungi are reported as sources of enzymes used in bioremediation [16-18]. New biosynthetic products have been revealed in marine-derived fungi [19-21]. Moreover, marine fungi can be exploited for monitoring marine environment pollution [22].
Literature search revealed no studies examining fungal biodiversity in Gulf of Aqaba, a semi-enclosed water body located in the most southern part of Jordan and at the northern end of the Red Sea. Only one study examined manglicolous fungal diversity in of the Red Sea in the upper Egypt [23].
The Gulf of Aqaba is a sub-tropical arid area between longitude 34˚25' to 35˚00'E and latitude 28˚00' to 29˚33'N (Figure 1). The Gulf is unique for its great depth (about 1830 m) in proportion to its width (maximum 25 km); the mean depth of the Gulf is about 800 m. Most of the rainfall in the region of the Gulf occurs during the period November-May. The average rainfall in Aqaba town is about 35 mm/yr. Daily temperature ranges between 14˚C in January to 45˚C in summer and evaporation rates of
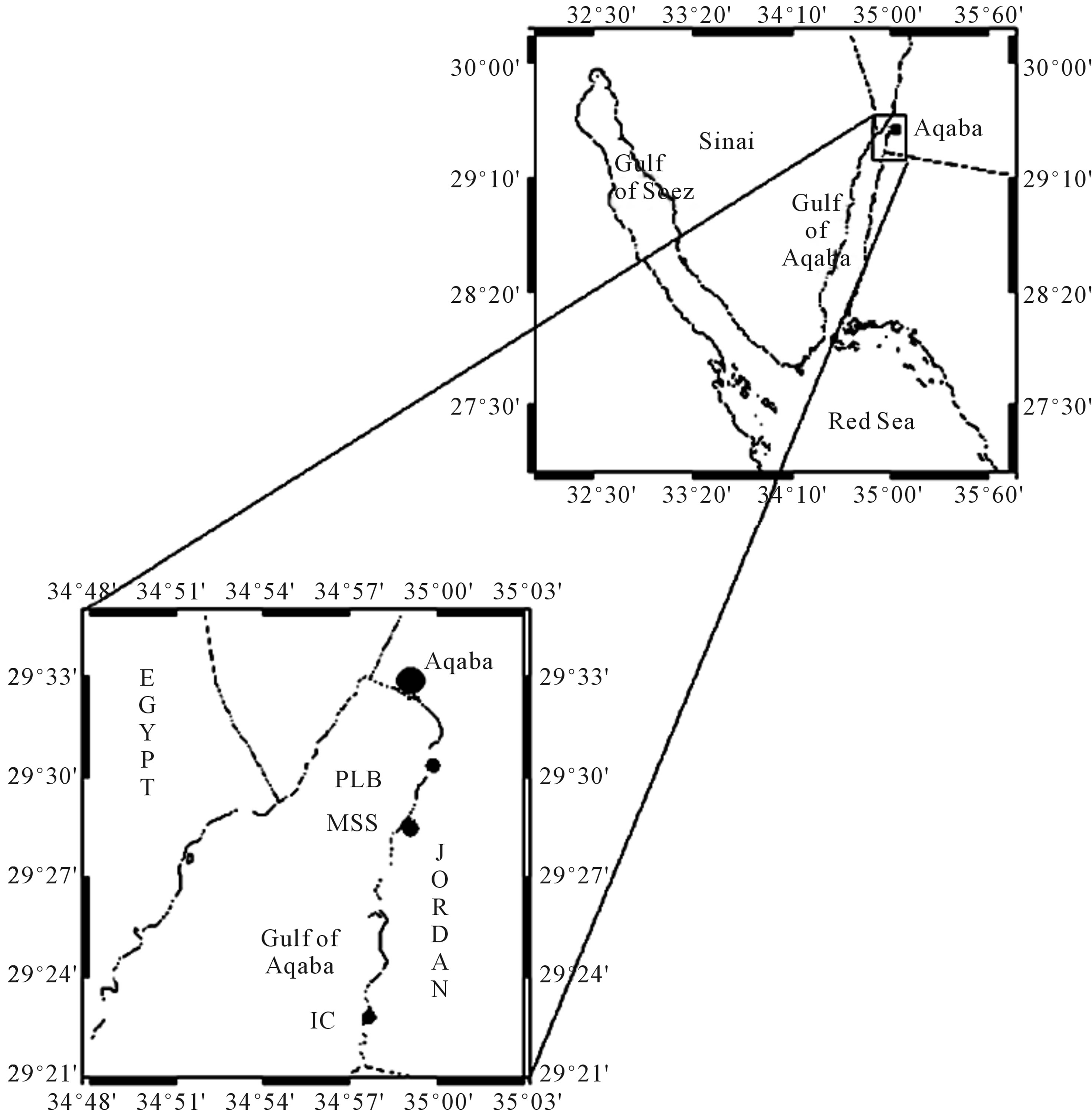
Figure 1. Location of the sampling sites along the coast of the Gulf of Aqaba.
seawater between 200 and 365 cm/yr [24]. The aim of this study is to investigate the existence of fungal life in seawater and sediments of the Gulf of Aqaba and determine their phylotypes. Using molecular identification approach, we report the isolation and identification of 5 Ascomycete sp. Partial sequences of the 18S small subunit (ssu) rRNA gene were used as queries in BLASTN search that revealed 98% - 100% identity. Phylogenetic analyses revealed close relationship among the isolated fungi and their potential affiliation with Ascomycetes. We propose further studies examining fungal biodiversity in this unique marine environment.
2. MATERIALS AND METHODS
2.1. Sampling Sites
Water samples were collected using 100 ml screw-cap sterilized bottle, from three selected locations, namely; Marine Science Station (MSS); Phosphate Loading Berth (PLB) and Industrial Complex area (IC) by snorkeling, or by SCUBA diving at a depth of 5 m, during Fall 2010.
The sampling sites are shown in Figure 1. Meanwhile sediment samples were collected from the same location using similar 100 ml sterilized plastic bottle. The sampling was performed twice during 2 months period in the fall 2010. All samples were transferred to the laboratory by using icebox and cultured within 40 hours of arrival.
2.2. Fungal Isolation and Culture Conditions
Seawater samples were diluted 1/10 using 0.5 M sterile NaCl solution, whereas sediment samples were first washed with sterile NaCl solution to prepare wash off solution. From both preparations, 100 μl of either diluted sample or wash off were taken to spread on 2.5% (w/v) Sabouraud Dextrose agar plates containing 50 μg/μl ampicillin. To rule out any possible contamination during manipulation, negative control plates received only sterile NaCl washing solution and were exposed to circulating air inside the laminar flow during culturing. All plates were incubated for 3 days at 30˚C. After incubation, cultured plates were screened to select isolated fungal colonies. Each selected colony was sub-cultured several times to ensure obtaining pure culture. Colony selection was primarily based on morphological differences between colonies. Although some colonies exhibited only slight differences, yet they were selected for further analysis. Nine colonies were selected for further identification analysis. Spores were harvested using sterile cotton swab pre-wetted with salt-tween [0.1% NaCl plus 0.1% tween-80 (vol/vol)] then released in salt-tween solution. After that, spores were counted using hemocytometer. Approximately 6 million spores from each isolate/colony were inoculated into Erlenmeyer flasks containing 25 mL of Sabouraud Dextrose Broth 2.5% (w/v) 50 μg/μl ampicillin. The flasks were incubated for 48 h at 30˚C and 150 rpm shaking. All inoculations and fungal handling were done under ClassII BIOAIR Laminar flow (Euroclone division, Italy) to avoid any possible air contamination.
2.3. Genomic DNA Extraction
Fungal mycelia of each isolate were collected by centrifugation at 10,000 rpm using regular bench-top centrifuge. Prior to that, microscopic examination was done to test the purity of the culture. Genomic DNA was extracted from 2 - 3 mg of mycelia using recombinant lyticase (Sigma, USA) and according to the protocol in wizard genomic DNA purification kit (Promega, USA).
2.4. Primers and 18S rRNA Gene PCR Amplification and Sequencing
Forward Primer 5’-ATT GGA GGG CAA GTC TGG TG-3’ and reverse primer 5’-CCG ATC CCT AGT CGG CAT AG-3’ bind to conserved regions of the fungal 18S ssu rRNA gene and enclosing containing the variable regions V7 and V9 as described before [25]. PCR amplification was carried out using ReadyMixTM Taq PCR Reaction Mix (Sigma, USA) which contains all required reagents except primers. PCR amplification was carried out in reaction volume of 50 µl of which 25 μl of ready mix Taq PCR which contained 1.5 units Taq DNA polymerase, 10 mM Tris-HCl, 50 mM KCl, 1.5 mM MgCl2, 0.001% gelatin and 0.2 mM dNTP, in addition to 2 µl of 10 μM of each primers and 5 µl (0.25 µg/µL) DNA templates were added to each reaction mixture. Blank that contained only water instead of DNA, was used in each PCR run to check for DNA contamination.
DNA amplification was carried out by thermal cycler (Biorad, USA). The samples were denatured at 94˚C for 2 minutes, followed by 30 cycles of repeated denaturation (30 s at 94˚C), annealing (45 s at 55˚C), enzymatic chain extension (60 s at 72˚C) and a final extension at 72˚C for 10 minute. Following amplification, PCR products were analyzed by 1% agarose gel electrophoresis in 1X TBE buffer (9 mM Tris-borate, 0.2 mM EDTA) and analyzed after staining with ethidium bromide.
2.5. Sequence Analysis
PCR products were sequenced at the Macrogen Company sequencing facility using Applied Biosystems 3730 mXL automated DNA sequencer (Macrogen, Korea). For identification, sequences were submitted to BLA-STN (National Center for Biotechnology Information; (http://www.ncbi.nlm.nih.gov). The 18S rRNA gene sequences from each species and the matched sequences from GenBank were aligned using ClustalW2 (EMBLEuropean Bioinformatics Institute-EBI; http://www.ebiac.uk). Phylogenetic analyses were performed using Phylogeney.fr [25]. Phylogentic tree was reconstructed using the maximum likelihood method implemented in the PhyML program (v3.0 alRT) and graphed using TreeDyn (V198.3) [26].
3. RESULTS
3.1. Phenotypic Characterization
Fungi were cultured from form the sediment and seawater of the Gulf of Aqaba, Red seas.
All isolates grew equally well on Sabouraud Dextrose agar plates at 30˚C. Most colonies grew up to 3 - 4 cm in diameter after 48 h incubation at 30˚C. No colonies grew on the negative control plates. According to their growth characteristics and colony morphology, 9 well-isolated colonies were selected for further characterization and identification.
3.2. PCR Amplification of the 18S ssu rRNA
The primers target a consensus sequence of the 18S ssu rRNA gene containing the variable regions V7 and V9. Amplification with the primers described above is expected to yield a 482- to 503-bp fragment, depending on the fungus tested. Figure 2 shows the amplicons resulted from PCR amplification. The figure does not reveal detectable size variation among amplicons.
3.3. 18S rRNA Gene Sequence Analysis
Table 1 summarizes the results of the 18S ssu rRNA gene sequence analysis for these fungal isolates. The amplified fungal sequences were used as BLASTN queries against the NCBI database. The sequences of the PCR amplicons were found to be 98% - 100% similar to the sequences of 18S rRNA regions of the respective genera and species of closely related fungi. For example, the fungal sequence amplified from Marine Science Station sediment isolate no. 5 (MSS_S5) showed 100% identity with an Aspergillus sydowi (Acc. No. JN938975) which was previously sequenced from Aspergillus sydowii strain DAOM 213727 18S ssu ribosomal RNA
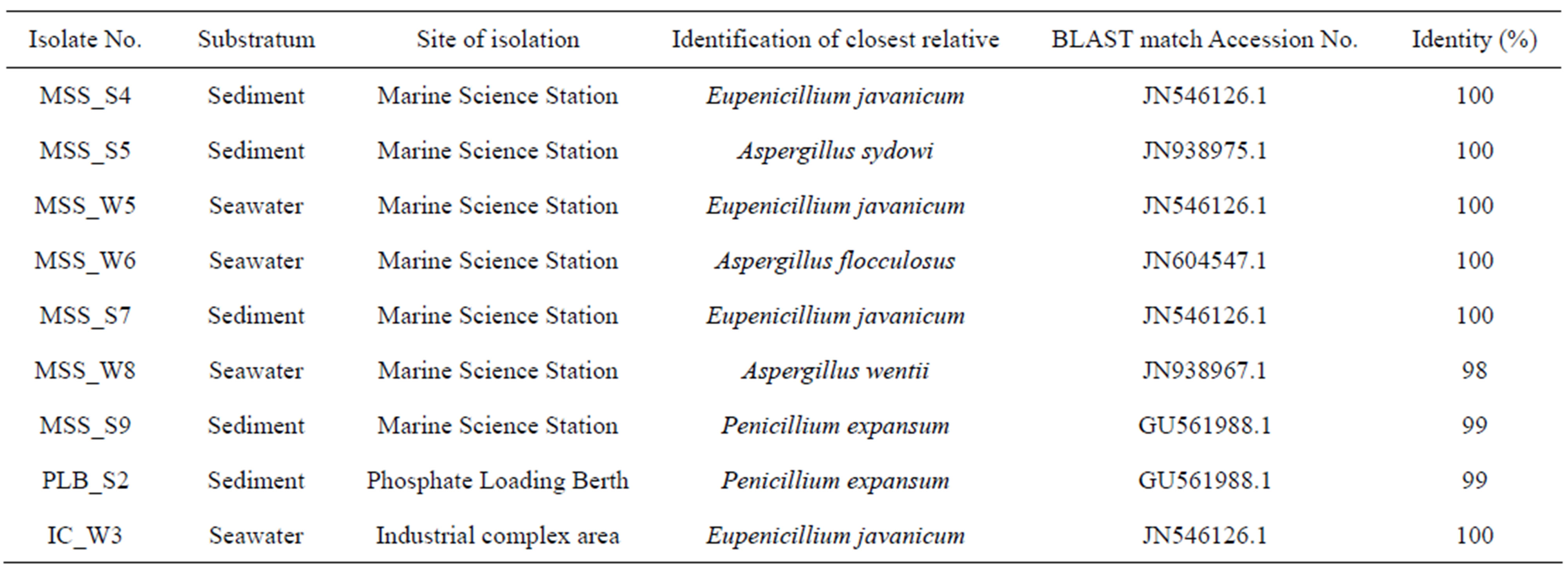
Table 1. Ascomycete isolates identified in this study with substratum, site of isolation, GenBank BLASTN match Acc. Nos. of 18S rRNA sequence and percentage identity.
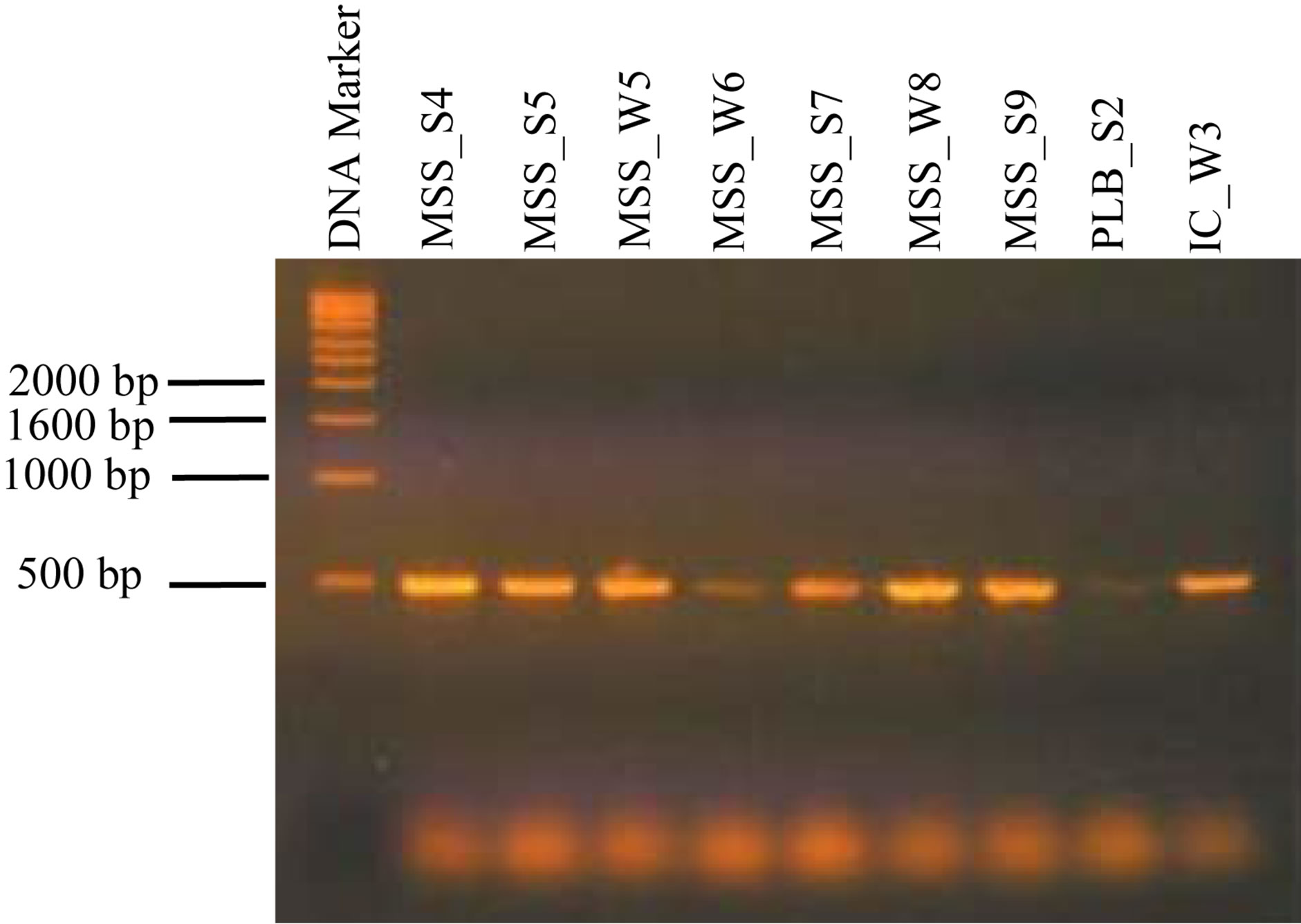
Figure 2. Amplification products of 18S ssu rRNA gene of fungal isolates. DNA marker (number on the left are in kilobase); MSS_S4, Eupenicillium javanicum; MSS_S5, Aspergillus sydowi; MSS_W5, Eupenicillium javanicum; MSS_W6, Aspergillus flocculosus; MSS_S7, Eupenicillium javanicum; MSS_W8, Aspergillus wentii; MSS_S9, Penicillium expansum; PLB_S2, Penicillium expansum; IC_W3, Eupenicillium javanicum. Fungal identification was based on closest relative resulted form BLASTN match.
gene partial sequence [27]. Similar patterns were observed amongst isolates of sediment MSS_4 and seawater MSS_W6 that also showed 100% identities to Eupenicillium javanicum and Aspergillus flocculosus respectively. Additionally, within our dataset, isolates from Marine Science Station sediment (MSS_S9) showed 99% Penicillium expansum whereas seawater isolate MSS_W8 exhibited 98% identity with Apergillus wentii. Therefore, 5 different species belonging to 3 different genera have been isolated, all of which belong to phylum Ascomycota. No other fungal phyla have been isolated from these samples. Based on 18S ssu rRNA gene sequences, a phylogenetic tree analyses using Phylogeny.fr (Figure 3) represents phylogenetic positions of Ascomycetes iso-
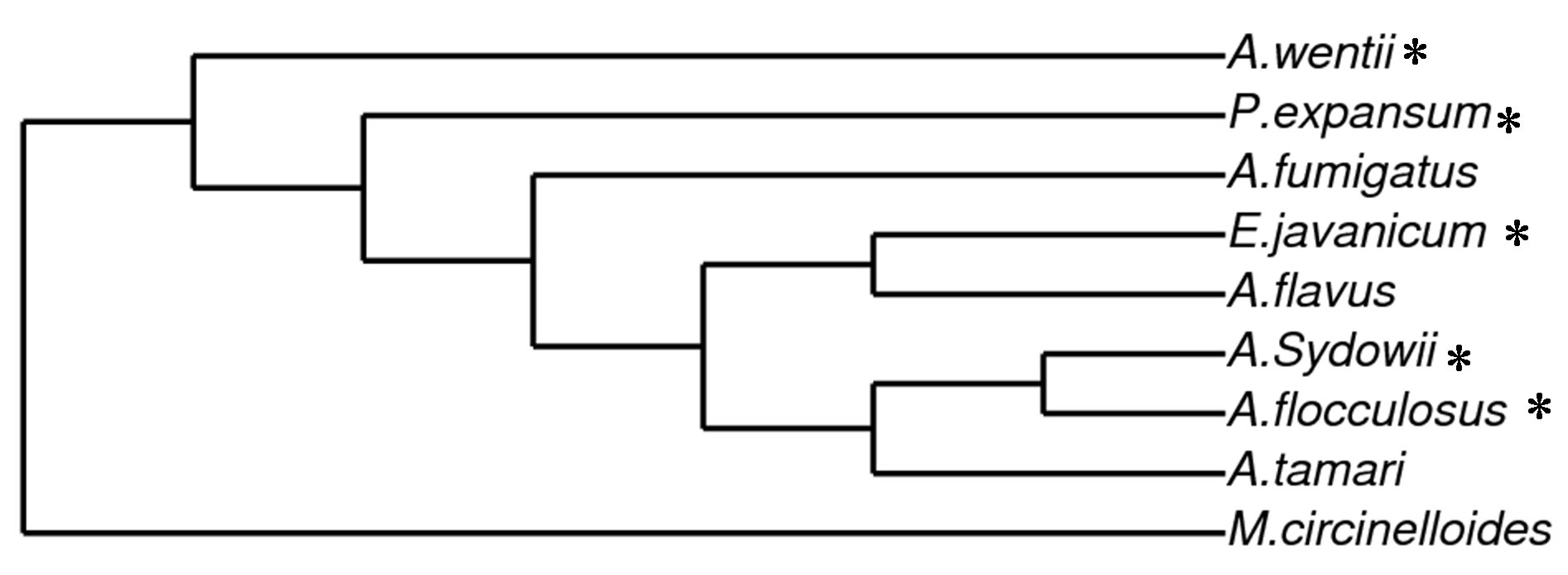
Figure 3. Maximum likelihood phylogram inferred from partial 18S rDNA (450 base pairs) sequence data showing phylogenetic relationship of Ascomycetes isolated in this study (indicted in asterisks) to other Aspergillus sp. (A. fumigatus, A. flavus and A. tamari, Acc. No HQ871898.1, JF824683.1 and AF516140.1 respectively) and Zygomecete (Mucor circinelloides, Acc. No. JN939019) as an outgroup. The phylogenetic tree was implemented in the PhyML program (v3.0 aLRT) and graphed using TreeDyn (V198.3).
lated in this study in relation to closely related Aspergillus sp. (A. fumigatus, A. flavus and A. tamari, Acc. No. HQ871898.1, JF824683.1 and AF516140.1 respectively) of which their 18S ssu rRNA sequences were obtained from GenBank. The selected Aspegillus sp. were among the list of hits obtained in BLASTN search. A Zygomecete (Mucor circinelloides Acc. No. JN939019) was used as an out-group. This results further supports observation that the isolated fungi are closely related and are most likely belong to the Ascomycetes group.
4. DISCUSSION
The 18S small subunit rRNA gene is one of the most frequently used genes in phylogenetic studies and considered as a key target for environmental biodiversity screening [28-30]. In this study, we isolated fungi from three different locations that cover coastline of 26 kilometers long in the Gulf of Aqaba, Red sea. Identification of isolated fungi was achieved using PCR amplification of partial 18S ssu rRNA gene followed by DNA sequence analyses. Five different isolates belonging to 3 different genera were identified, namely; Eupenicillium, Penicillium and Aspergillus all of which belong to phylum Ascomycota. It has been well documented that Ascomycetes include a group of obligate marine fungi that tremendously contribute to marine ecosystem and represents variety of saprobes, pathogens and symbiont of coastal and deep-see habitats [9,22,31,32]. Twenty six marine Ascomycetes have been previously isolated form mangrove wood and leaves in the intertidal zone of Red sea area at upper Egypt [22]. Eurotiomycetes of the phylum Ascomycota are the most frequently detected fungal taxa from deep-sea environments, followed by the classes Saccharomycete, Dothideomycete and Sordariomycete. The majority of species belonging to the Eurotiomycete are members of the Aspergillus and Penicillium genera, which are known to be globally distributed and ubiquitous in marine as well as terrestrial environments [5].
Although Aspergillus sp. is more commonly known as terrestrial fungi, many species of these mycelial fungi have been isolated from marine habitats. For example the ubiquitous existence of A. sydowii, A. niger, A. wentii, A. flavus, A. insulicola and A. Awamori. in marine environment have been established by both molecular and culture-dependent techniques [9,15,19,32-34]. In our study, two Aspergillus sp. isolated from sediment and seawater of the Gulf of Aqaba showed strong sequence homology to A. sydowii and A. wentii respectively. In addition, we isolated Penicillium sp. and Eupenicillium sp. from sediment samples. Penicillium has previously described as a marine-derived fungus [21]. However, to our knowledge, no reports have documented Eupenicillium sp. as a marine-derived fungus. It is anticipated that sequence data of 18S ssu RNA gene obtained from various sea environments will reveal more facultative marine fungi and perhaps novel fungal groups. Although members of Basidiomycota and lower fungi Chytridiomycota have been reported as inhabitants of marine fungi [5,9], we did not find any Basidiomycete or Chytridiomycete in our samples. Perhaps larger scale sampling will increase likelihood of isolating members of these groups in the Gulf of Aqaba.
Another aspect that highlights the importance of this study is based on the ability of fungi to overcome natural selection. Fungi to ensure well growth, sporulation and multiplication must barriers and acquire new capabilities that enable them adjust to new environmental conditions such as utilization of new carbon sources, adjustment to new osmotic—as well as hydrostatic—pressures and, in some instances, new temperature conditions [4,32]. Therefore, terrestrial fungi differ from its marine counterparts in; at least, metabolic activities and patterns of carbon source utilization. This suggests that, unlike terrestrial fungi, marine-derived fungi perhaps produce new secondary metabolites that can be exploited environmentally, biologically and perhaps pharmaceutically. For example, Sorbiterrin with a novel cholinesterase activity was isolated from marine-derived fungus Penicillium terrestre [20]. Cytotoxicity against selected cancer cells has been measured in vitro with compounds isolated from marinederived fungus Aspergillus insulicola [19]. Marine derived fungi have shown their potential in polycyclic aromatic hydrocarbon bioremediation [16]. Penicillium griseofulvum as a eukaryotic organism has been exploited to screen heavy metal pollution in marine contaminated sediment [21]. Taken together, these studies indicate that filamentous fungi of marine environments might be considered as valuable sources of by-product compounds and enzymes that can be of industrial, pharmaceutical and environmental importance.
Our results report the presence of facultative marine Ascomycetes in the Gulf of Aqaba, Red sea and set the stage towards future investigations concerning marine fungal biodiversity in this area. We further propose exploration of marine-derived fungi as potential sources of valuable products.
5. ACKNOWLEDGEMENTS
This study was funded by the Deanship of Academic Research of the University of Jordan.
REFERENCES
- Kis-Papo, T., Oren, A. and Nevo, E. (2003) Survival of filamentous fungi in hypersaline Dead Sea water. Microbial Ecology, 45, 183-190. doi:10.1007/s00248-002-3006-8
- Lovejoy, C., Massana, R. and Pedros-Alio, C. (2006) Diversity and distribution of marine microbial eukaryotes in the Arctic Ocean and adjacent seas. Applied Microbiology and Microbiology, 72, 3085-3095.
- Gao, Z., Binglin, L., Zheng, C. and Wang, G. (2008) Molecular detection of fungal communities in the howian marine sponges Suberites seteki and Mycale armata. Applied Environmental and Microbiology, 74, 6091-6101. doi:10.1128/AEM.01315-08
- Burgaud, G., Le Calves, T., Arzur, D., Vandenkoornhuyse, P. and Barbier, G. (2009) Diversity of culturlable marine filamentous fungi from deep-sea hydrothermal vents. Environmental Microbiology, 11, 1588-1600. doi:10.1111/j.1462-2920.2009.01886.x
- Nagano, Y. and Nagahama, T. (2012) Fungal diversity in deep-sea extreme environments. Fungal Ecology, 5, 463- 471. doi:10.1016/j.funeco.2012.01.004
- Hawksworth, D.L. (1991) The fungal dimension of biodiversity: Magnitude, significance, and conservation, Mycological Research, 95, 641-655. doi:10.1016/S0953-7562(09)80810-1
- Hawksworth, D.L. (2001) The magnitude of fungal diversity: The 1.5 million species estimated revisited. Mycological Research, 105, 1422-1432. doi:10.1017/S0953756201004725
- Blackwell, M. (2011) The fungi: 1,2,3…5.1 million species. American Journal of Botany, 98, 426-438. doi:10.3732/ajb.1000298
- Kohlmeyer, J. and Kohlmeyer, E. (1979) Marine mycology—The higher fungi. Academic Press, New York.
- Hyde, K.D., Jones, E.B.G., Leano, E., Pointing, S.B., Poonyth, A.D. and Vrigmoed, L.P. (1989) Role of fungi in marine ecosystems. Biodiversity and Conservation, 7, 1147-1161. doi:10.1023/A:1008823515157
- Barata, M. (2006) Marine fungi from Mira river salt marsh in Portugal. Revista Iberoamerican de Micologia, 23, 179-184. doi:10.1016/S1130-1406(06)70040-7
- Zuluaga-Montero, A., Rameriz-Cameho, L., Rauscher, J. and Bayman, P. (2010) Marine isolates o Aspergillus fumigatus: Denzins of the deep or lost at sea? Fungal Ecology, 3, 386-391. doi:10.1016/j.funeco.2010.05.003
- Zuluaga-Montero, A., Toledo-Hernandez, C., Rodriguez J.A., Sabat, M. and Bayman, P. (2010) Spatial variation in fungal communities isolated from healthy and diseased sea fans Gorgonia ventalina and seawater. Aquatic Biology, 8, 151-160. doi:10.3354/ab00218
- Bourne, D.G., Garren, M., Work, T.M., Rosenburg, E., Smith, G.W. and Harvell, C.D. (2009) Microbial disease and the coral holobiont. Tends in Microbiology, 17, 554- 562. doi:10.1016/j.tim.2009.09.004
- Kales, S.K., De Witte-Orr, S.J., Bols, N.C. and Dixon, B. (2007) Response of rainbow trout monocyte/macrophage cell line, RTS11 to the water molds Achyla and Saprolegnia. Molecular Immunology, 44, 2303-2314. doi:10.1016/j.molimm.2006.11.007
- Basheer, S.M., Chellappan, S., Beena, P.S., Sukumaran, R.K., Elyas, K.K. and Chandrasekaran, M. (2011) Lipase from marine Aspergilus awamori BTMFW032; Production, partial purification and application in oil effluent treatment. New Biotechnology, 28, 627-638. doi:10.1016/j.nbt.2011.04.007
- Passarini, M.R.Z., Rodrigues, M.V.N., daSilva, M. and Sette, L.D. (2011) Marine-derived filamentous fungi and their potential application for polycyclic aromatic hydrocarbons bioremediation. Marine Pollution Bulletin, 62, 364-370. doi:10.1016/j.marpolbul.2010.10.003
- Rocha, L.C. Ferreira, H.V., Luiz, R.F., Sette, L.D. and Porto, A.L.M. (2011) Seroselctive bioreduction of 1-(4- Methoxypheyl) ethanone by whole cells marine-derived fungi. Marine Biotechnology, In Press.
- Wang, F., Fang, Y., Zhang, M., Lin, A, Zhu, T., Gu, Q. and Zhu, W. (2008) Six new ergosterol from marinederived fungus Rhizopus sp. Steroids, 73, 19-26. doi:10.1016/j.steroids.2007.08.008
- Wu, Q.X., Jin, X.J., Draskovic, M., Crews, M.S., Tenney, K., Valeriote, F.A., Yao, X.J. and Crews, P. (2012) Unraveling the numerous biosynthetic products of the marine sediment-derived fungus, Aspergillus insulicola. Phytochemistry Letters, 5, 114-117. doi:10.1016/j.phytol.2011.11.005
- Chen, L., Zhu, T., Ding, Y., Khan, I.A., Gu, Q., Li, D. and Sorbiterrin, A. (2012) A novel sorbicillin derivative with cholinesterase inhibition activity from the marine-derived fungus Penicillium terrestre. Tetrahedron Letters, 53, 325- 328. doi:10.1016/j.tetlet.2011.11.038
- Shi, Z., Bai, S., Tian, L., Jiang, H. and Zhang, J. (2011) Molecular detection of Penicillium griseofulvum as the coastal pollution indicator. Current Microbiology, 62, 396-401. doi:10.1007/s00284-010-9720-4
- El-Sharouny, H.M., Raheem, A.M. and Abdel-Wahab, M.A. (1998) Manglicolous fungi of the red sea in upper Egypt. Microbiological Research, 153, 81-96. doi:10.1016/S0944-5013(98)80025-6
- Hulings, N.C. (1989) A review of the marine science research in the Gulf of Aqaba, Marine Science Station, Aqaba-Jordan.
- Loeffler, J., Henke, N., Hebart, H., Schmidt, D., Hagmeyer, L., Schumacher, U. and Einsele, H. (2000) Quantification of fungal DNA by using fluorescence resonance energy transfer and light cycler system. Journal of Clinical Microbiology, 38, 586-590.
- Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., Dufayard, J.F., Guindon, S., Lefort, V., Lescot, M., Claverie, J.M. and Gascuel, O. (2008) Phylogeny.fr: Robust phylogenetic analysis for the nonspecialist. Nucleic Acid Research, 36, W465-W469. doi:10.1093/nar/gkn180
- Schocha, C.L., Seifert, K.A, Huhndorf, S., Robert, V., Spougea, J.L., Levesque, C.A., Chen, W. and Consortiuma, F.B. (2012) Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proceedings of National Academy of Science, 109, 6241-6246. doi:10.1073/pnas.1117018109
- Geiser, D.M., Gueidan, C., Miadlikowska, J., Lutzoni, F., Kauff, F., Hofstetter, V., Fraker, E., Schoch, C.L., Tibell, L., Untereiner, W.A. and Aptroot, A. (2006) Eurotiomycetes: Eurotiomycetidae and Chaetothyriomycetidae. Mycologia, 98, 1053-1064. doi:10.3852/mycologia.98.6.1053
- Wu, Z., Tsumura, Y., Blomquist, G. and Wang, X. (2003) 18S rRNA gene variation among common airborne fungi, and development of specific oligonucleotide probes for the detection of fungal isolate. Applied and Environmental Microbiology, 69, 5389-5397. doi:10.1128/AEM.69.9.5389-5397.2003
- Chenuil, A. (2006) Choosing the right molecular genetic markers for studying biodiversity: From molecular evolution to practical aspects. Genetica, 1, 101-120.
- Nambiar, G.R. and Raveendran, K. (2008) A checklist of marine fungi from Kerala state, India. American-Eurasian Journal of Botany, 3, 73-77.
- Bhadury, B., Bik, H., Lambshead, J.D., Austen, C.M., Smerdon, G.R. and Rogers, A.D. (2011) Molecular diversity of fungal phylotypes co-amplified alongside nematodes from coastal and deep-sea marine environments. PLoS One, 6, e26445. doi:10.1371/journal.pone.0026445
- Raghukumar, C., Raghukumar, S., Sheelu, G., Gupta, S.M., Nath, B.N. and Rao, B.R. (2004) Buried in time: Culturable fungi in a deep-sea sediment core from the chagos Trench, Indian Ocean. Deep-Sea Research I, 51, 1759-1768.
- Takishita, K., Yubuki, N., Kakizoe, N., Inagaki, Y. and Maruyama, T. (2007) Diversity of microbial eukaryotes in sediment at a deep-sea methane cold seep: Surveys of ribosomal DNA libraries from raw sediment samples and two enrichment cultures. Extremophiles, 4, 563-576. doi:10.1007/s00792-007-0068-z