Advances in Bioscience and Biotechnology
Vol.05 No.13(2014), Article ID:51516,6 pages
10.4236/abb.2014.513117
Feeding Deterrence of Common Spices against Helicoverpa armigera Larvae
Weizheng Li, Jingjing Hu, Jing Yang, Guohui Yuan*, Xianru Guo, Meihao Luo
Email: *hnndygh@126.com
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 26 September 2014; revised 26 October 2014; accepted 13 November 2014
ABSTRACT
Edible spices with strong smells or heavy tastes may be a promising resource of feeding deterrents. We compared the feeding deterrence of the ethanol extracts of 21 common spices against the larvae of a generalist pest species, Helicoverpa armigera, using a multiple-choice leaf disc bioassay. The results show that Zanthoxylum bungeanum extract (as a reference) always evoked significant feeding deterrence, while Piper nigrum (both black pepper and white pepper), Piper longum, and Angelica dahurica evoked the strongest and equivalent feeding deterrence. The potent feeding deterrent activity of Piper species may be a common characteristic at genus level.
Keywords:
Helicoverpa armigera, Spice, Feeding Deterrent, Piper

1. Introduction
Plant-derived feeding deterrents have attracted increasing attention in integrated pest management [1] . Despite this agricultural importance, there have been only a limited number of applicable products [2] , almost none avai- lable on highly polyphagous species like Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). It has been demonstrated that specialist herbivores showed greater sensitivity and lower habituation to plant-derived feeding deterrents than generalists [3] , and thus it can be expected that an effective feeding deterrent targeting to a polyphagous species would also control the co-occurring oligophagous or monophagous ones [4] .
H. armigera is one of the most important economic insect pests in many parts of the world. The highly polyphagous larva has been recorded as a major pest of maize, sorghum, tomato, lucerne, tobacco, cotton, and cowpea. Extensive insecticide spraying resulted in low yields and high control costs, and the ever-increasing resistance has led to a renewed interest in developing alternatives to insecticidal control [5] . With this respect, several deterrent plants or their extracts have been reported for H. armigera [6] - [9] . However, common spices with strong smells or heavy tastes would provide a promising feeding deterrent source, but have not received enough attention.
We previously found the ethanol extract of Zanthoxylum bungeanum, a numb-taste spice, strongly inhibited H. armigera larvae feeding [10] , and thus it was an impetus to compare more spices. The first phase of this study involves the laboratory screening and comparison of the potential relative feeding deterrence of various pungent or hot-tasting spices so as to identify a few promising ones for subsequent detailed investigations.
2. Materials and Methods
2.1. Insect Culture
H. armigera larvae collected in a tomato field in Scientific & Education Yard, Henan Agricultural University were brought to the laboratory and maintained in a climatic incubator as continuous cultures under 16L:8D photoperiod, 26˚C ± 2˚C temperature, and 60% - 70% relative humidity, with the dark period between 23:00 and 7:00 h. Larvae were reared individually in 15 mL glass tubes and provided with a small block of wheat germ- based artificial diet. The larvae used in this study had been maintained for at least 10 generations and were used once only in the bioassays. Adults were kept in plastic cages and supplied with a 5% sucrose solution.
2.2. Plant Extraction
In this study, we tested the feeding deterrence of a total of 21 spices, obtained from Huanghe Food Market, Zhengzhou, China. Among them, Zanthoxylum bungeanum, which had strong feeding deterrent effect on H. armigera fourth-instar larvae reported previously [10] [11] , was used as a reference to determine the possible between-group difference (see below), and the remained 20 spices (Table 1) were selected arbitrarily on the basis of heavy smell and taste to human beings, irrespective of what might be regarded as “host plants” or “non-host plants”. In addition, feeding deterrence of ethanol extract of Z. bungeanum was much stronger than those extracted with the other solvents such as dichloromethane, distilled water, acetone, and n-hexane [11] , so we believe that ethanol is also suitable for extracting the other spice materials. Thirty grams of each of material dried powders (sieve 0.50 mm) were extracted in 225 mL absolute ethanol in a sealed glass container for 24 h by cold percolation, then the filtrates were added up to 225 mL with absolute ethanol to compensate for solvent evaporation.
2.3. Bioassay
Multiple-choice leaf disc bioassay was used, because there were a great many of candidate spices in this study, and by using this method we could compare several materials in a direct confrontation mode. The materials were firstly divided into five groups, each group included a reference (Z. bungeanum ethanol extract), a control (solvent ethanol only), and four randomly chosen extracts (Groups 1-5, Table 2). Each group was repeated for 17 - 22 times. To overcome possible grouping effect, the extracts whose leaf consumptions were significantly lower than corresponding controls in the first round were re-grouped, each group included a reference, a control, and three or four samples (Groups 6-8, Table 2). Each group was repeated for 10 times.
Tobacco (Nicotiana tabacum var. K326) leaf was used as a feeding substrate, since that H. armigera larvae preferred tobacco leaves over those of corn, cotton, tomato, and hot pepper [11] . Leaf discs (15 mm ID), with 20 μL extracts or solvent applied on their upper surface, were randomly arranged at an equal interval in the periphery of the Petri dishes (140 mm ID × 20 mm H) lined with moistened filter paper. After ca. 10 min, fourth instar larvae starved for 4 h were introduced individually in the centre of each Petri dish. The dishes were transferred into a climatic chamber at a temperature of 25˚C ± 2˚C, relative humidity (60% - 70%) and 16L:8D. The bioassay terminated after 10 h feeding, then insects removed, feeding amounts were measured using transparent millimeter—square graph paper.
2.4. Data Analysis
All of the following analyses were performed on SPSS 19.0 for Windows. The activities of feeding deterrence was calculated by the following formula: Feeding Deterrence Index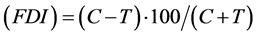 , where C
, where C

Table 1. The plant materials used in the present study.
aPeppercorn picked when still green and dried in the sun until it turns black; bPeppercorn ripens fully on the vine before picked.
represents the average consumed areas of the control discs and T the average consumed areas of treated discs within a group [12] [13] . The overall differences in feeding amounts within each group were compared using Friedman test, because the leaf consumptions within a Petri dish were not independent [14] [15] . Wilcoxon signed ranks test was used to verify any significant differences between each treatment and its corresponding control, to do this all P-values are corrected by Bonferroni-Dunn method.
3. Results
The mean feeding amounts and FDI values of each group are summarized in Table 2. Generally, the overall feeding amount difference in each group is significant, as shown in Table 2, and the reference sample always evoked significant feeding deterrence. In Group 1, the feeding amounts of leaf discs coated with the extracts of P. longum, Z. bungeanum, and M. fragrans were significant lower than that of the control, and P. longum is the strongest material with a FDI value 88.76 and mean feeding amount only 3.95 mm2. In Group 2, E. globulus extract exhibited significant feeding deterrent activity but not stronger than the reference. In Group 3, the extract of A. dahurica produced the strongest activity with a FDI value as high as 98.03, the mean leaf area consumed was only 1.00 mm2, followed by I. verum extract being equal to the reference. In Group 4, P. nigrum (white pepper) evoked the maximum FDI value of 97.49, and the mean leaf area consumed was only 0.88 mm2. In Group 5, P. nigrum (black pepper) extract had the strongest feeding deterrence activity with a FDI value of 93.86, and the mean leaf area consumed was only 1.05 mm2, followed by F. vulgare, Z. bungeanum, and C. reticulate. However, in multiple-choice bioassay, a sample exhibiting “significant” effect may be caused by its members, as shown by the FDI values of the reference in each group (ranging from 42.34% to 55.50%), we believe that regrouping may resolve this problem.
Groups 6, 7, and 8 was on the basis of the above results. In the second round, the extracts of I. verum, C. reticulata, M. fragrans, E. globulus, and F. vulgare were excluded from the active extract list, because their mean
Table 2. Feeding deterrence of ethanol extracts of test spices against H. armigera.
aMean feeding amounts followed by “*” and “**” within each group are significantly different from the corresponding control at P = 0.05 and P = 0.01 levels, Friedman test with Bonferroni-Dunn correction.
percent feeding amounts had not significant difference compared to their corresponding controls. Taken together, the three materials from Piperaceae [P. nigrum (black), P. nigrum (white), and P. longum] and A. dahurica had the strongest and almost equally feeding deterrence to H. armigera larvae.
4. Discussion
The use of insecticides is a double-bladed sword in crop protection. Although they no doubt play an important role in protecting the crops from damage, adverse effect on natural enemies also can not be ignored. Feeding deterrents may be an alternative to resolute this technical contradiction since they may protect crops from damage meanwhile offered the natural enemies with “non-toxic” prey. To date, the most widely used and most successful feeding deterrent is azadirachtin, whose anti-feeding effect is highly variable among pest species, and even those species initially deterred are often capable of rapid desensitization [2] , additionally, its noxious bitter taste further limited its application in some frequently harvested crops such as vegetables. From a prospect of practice, the edible spices would be a promising resource of feeding deterrents.
With this regard, we tested the feeding deterrence of some common edible spices against H. armigera. These includes four species from Apiaceae, three species each from Rutaceae and Zingiberaceae, two species from Lauraceae, and one species each from Brassicaceae, Illiciaceae, Myristicaceae, Myrtaceae, Solanaceae, and Valerianaceae. Among these, all the Piper materials exhibited strongest and equal feeding deterrence. Similarly, Piper cenocladum and P. imperiale were deterrent to Spodoptera frugiperda, another noctuid generalist [4] , and other herbivores such as two Hymenoptera species (Paraponera clavata and Atta cephalotes) and two Orthoptera species (Microtylopteryx hebardi and Homeomastax robertsi) [16] [17] . However, to the best of our knowledge, this is the first report on the feeding deterrence activity of P. nigrum and P. longum towards H. armigera. This study and the previous reports suggest that the potent feeding deterrent activity may be a common characteristic at genus level. In contrast, Z. bungeanum always evoked fairly strong feeding deterrence within each group, while Z. schinifolium did not, although they belong to the same genus, indicating that even phylogenetically closely related plants did not necessarily share common defense traits.
The phytochemistry of the family Piperaceae and the genus Piper is well documented [18] [19] , many species contain insecticidal secondary metabolites, providing a broad array of defenses against arthropod herbivores, the predominant ingredient is piperine [16] . Furthermore, the extract of another related species, Piper guineense, could significantly inhibit the hatching of the newly-deposited eggs of Maruca vitrata and Clavigralla tomentosicollis [20] , and suggesting that the action mode of Piper species is more complex than had been realized, although the active compounds inhibiting egg hatching have not been characterized yet. A. dahurica was the only one plant species exhibiting potent feeding deterrent activity among the four Apiaceae plants, which mainly contained a variety of furanocoumarin derivates [21] [22] . Unfortunately, these compounds are not available in our laboratory presently.
A potential feeding deterrent may act via the following mechanisms: 1) olfactory repellent effect, in this case, the larvae will reject the treated feeding substrate without contacting, 2) true taste deterrence, then the larvae will cause some small biting points on the substrate, 3) physical obstacle via oral mechanoreceptors, and/or 4) toxicity via post-ingestive response, if this is true, the larvae will consume relatively much food before they reject them [23] . In our bioassay, the first and the second possible mechanisms could not be discriminate yet, but the fourth mechanism cannot explain our results since all of the test subjects were living after bioassay.
5. Conclusion
In conclusion, only four materials were finally determined as the effective feeding deterrent resources of H. armigera larvae, except for Z. bungeanum, which was determined in a previous study. The isolation and purification of crude extract of Piper species and A. dahurica, as well as their long-term effect and possible mechanism involved in feeding deterrence, deserve to be explored further.
Acknowledgements
This work was granted by Natural Science Foundation of China (31471772) and The National Special Research Fund for Non-Profit Sector (Agriculture) (201203036). Special thanks are given to the following undergraduate trainees, Ruihong Wang, Li Zhuang, Ruilin Wang, Liwei Han, Jinlei Li.
References
- Ulrichs, C., Mewis, I., Adhikary, S., Bhattacharyya, A. and Goswami, A. (2008) Antifeedant Activity and Toxicity of Leaf Extracts from Porteresia coarctata Takeoka and Their Effects on the Physiology of Spodoptera litura (F.). Journal of Pest Science, 81, 79-84. http://dx.doi.org/10.1007/s10340-007-0187-4
- Isman, M.B. (2006) Botanical Insecticides, Deterrents and Repellents in Modern Agricultural and an Increasingly Regulated World. Annual Review of Entomology, 51, 45-56. http://dx.doi.org/10.1146/annurev.ento.51.110104.151146
- Bernays, E.A., Oppenheim, S., Chapman, R.F., Kwon, H. and Gould, F. (2000) Taste Sensitivity of Insect Herbivores to Deterrents is Greater in Specialists than in Generalists: A Behavioral Test of the Hypothesis with Two Closely Related Caterpillars. Journal of Chemical Ecology, 26, 547-563. http://dx.doi.org/10.1023/A:1005430010314
- Richards, L.A., Dyer, L.A., Smilanich, A.M. and Dodson, C.D. (2010) Synergistic Effects of Amides from Two Piper Species on Generalist and Specialist Herbivores. Journal of Chemical Ecology, 36, 1105-1113. http://dx.doi.org/10.1007/s10886-010-9852-9
- Jallow, M.F.A., Cunningham, J.P. and Zalucki, M.P. (2004) Intra-Specific Nariation for Host Plant Use in Helicoverpa armigera: Implications for Management. Crop Protection, 23, 955-964. http://dx.doi.org/10.1016/j.cropro.2004.02.008
- Koul, O., Jain, M.P. and Sharma, V.K. (2000) Growth Inhibitory and Antifeedant Activity of Extracts from Melia dubia to Spodoptera litura and Helicoverpa armigera Larvae. Indian Journal of Experimental Biology, 38, 63-68.
- Sundararajan, G. and Kumuthakalavalli, R. (2001) Antifeedant Activity of Aqueous Extract of Gnidia glauca Gilg. and Toddalia asiatica Lam. on the Gram Pod Borer, Helicoverpa armigera (Hübner). Journal of Environmental Biology, 22, 11-14.
- Ramya, S., Rajasekaran, C., Sundararajan, G., Alaguchamy, N. and Jayakumararaj, R. (2008) Antifeedant Activity of Leaf Aqueous Extracts of Selected Medicinal Plants on VI Instar Larva of Helicoverpa armigera (Hübner). Ethno- Botanical Leaflets, 12, 938-943.
- Kamaraj, C., Rahuman, A.A. and Bagavan, A. (2008) Screening for Antifeedant and Larvicidal Activity of Plant Extracts against Helicoverpa armigera (Hübner), Sylepta derogata (F.) and Anopheles stephensi (Liston). Parasitology Research, 103, 1361-1368. http://dx.doi.org/10.1007/s00436-008-1142-x
- Li, W.Z., Fu, G.X., Wang, Y.H., Yuan, G.H., Zhang, Y.C. and Chai, X.L. (2010) Feeding Responses of Helicoverpa armigera Larvae to Taste Compounds of Humans. Acta Ecologica Sinica, 30, 5709-5715.
- Yang, J., Li, W.Z., Chai, X.L., Yuan, G.H., Fu, G.X., Wang, Y.H., Guo, X.R. and Luo, M.H. (2013) Antifeedant Activity of Numb and Salty Taste Compounds against the Larvae of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Acta Ecologica Sinica, 33, 7-11. http://dx.doi.org/10.1016/j.chnaes.2012.12.002
- Akhtar, Y. and Isman, M.B. (2004) Generalization of a Habituated Feeding Deterrent Response to Unrelated Antifeedants Following Prolonged Exposure in a Generalist Herbivore, Trichoplusia ni. Journal of Chemical Ecology, 30, 1349-1362. http://dx.doi.org/10.1023/B:JOEC.0000037744.73291.b6
- Akhtar, Y., Pages, E., Stevens, A., Bradbury, R., da Camara, C.A.G. and Isman, M.B. (2012) Effect of Chemical Complexity of Essential Oils on Feeding Deterrence in Larvae of the Cabbage Looper. Physiological Entomology, 37, 81-91. http://dx.doi.org/10.1111/j.1365-3032.2011.00824.x
- Roa, R. (1992) Design and Analysis of Multiple-Choice Feeding-Preference Experiments. Oecologia, 89, 509-515.
- Lockwood III, J.R. (1998) On the Statistical Analysis of Multiple-Choice Feeding Preference Experiments. Oecologia, 116, 475-481. http://dx.doi.org/10.1007/s004420050612
- Dyer, L.A., Dodson, C.D., Beihoffer, J. and Letourneau, D.K. (2001) Trade-Offs in Antiherbivore Defenses in Piper cenocladum: Ant Mutualists versus Plant Secondary Metabolites. Journal of Chemical Ecology, 27, 581-592. http://dx.doi.org/10.1023/A:1010345123670
- Dyer, L.A., Dodson, C.D., Stireman, J.O., Tobler, M.A., Smilanich, A.M., Fincher, R.M. and Letourneau, D.K. (2003) Synergistic Effects of Three Piper Amides on Generalist and Specialist Herbivores. Journal of Chemical Ecology, 29, 2499-2514. http://dx.doi.org/10.1023/A:1026310001958
- Parmar, V.S., Jain, S.C., Bisht, K.S., Jain, R., Taneja, P., Jha, A., Tyagi, O.D., Prasad, A.K., Wengel, J., Olsen, C.E. and Boll, P.M. (1997) Phytochemistry of the Genus Piper. Phytochemistry, 46, 597-673. http://dx.doi.org/10.1016/S0031-9422(97)00328-2
- Dyer, L.A. and Palmer, A.D.N. (2004) Piper: A Model Genus for Studies of Phytochemistry, Ecology, and Evolution. Kluwer Academic/Plenum Publishers, New York.
- Ekesi, S. (2000) Effect of Volatiles and Crude Extracts of Different Plant Materials on Egg Viability of Maruca vitrata and Clavigralla tomentosicollis. Phytoparasitica, 28, 305-310. http://dx.doi.org/10.1007/BF02981825
- Baek, N.I., Ahn, E.M., Kim, H.Y. and Park, Y.D. (2000) Furanocoumarins from the Root of Angelica dahurica. Archives of Pharmacal Research, 23, 467-470. http://dx.doi.org/10.1007/BF02976574
- Wang, T.T., Jin, H., Li, Q., Cheng, W.M., Hu, Q.Q., Chen, X.H. and Bi, K.S. (2007) Isolation and Simultaneous Determination of Coumarin Compounds in Radix Angelica dahurica. Chromatographia, 65, 477-481. http://dx.doi.org/10.1365/s10337-007-0185-y
- Glendinning, J.I. (2008) Insect Gustatory Systems. In: Basbaum, A.I., Kaneko, A., Shepherd, G.M., Westheimer, G., Firestein, S. and Beauchamp, G.K., Eds., The Senses: A Comprehensive Reference, Academic Press, San Diego, 75-96.
NOTES
![]()
*Corresponding author.