American Journal of Plant Sciences
Vol.08 No.06(2017), Article ID:76339,21 pages
10.4236/ajps.2017.86084
Geographical Variation, Distribution and Diversity of Rice Yellow Mottle Virus Phylotypes in Tanzania
Judith Hubert*, Herman J. F. Lyimo, Ashura Luzi-Kihupi
Department of Crop Science and Production, Sokoine University of Agriculture, Morogoro, Tanzania

Copyright © 2017 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: March 31, 2017; Accepted: May 20, 2017; Published: May 23, 2017
ABSTRACT
Rice yellow mottle virus (RYMV) is the most important disease of rice in Africa. The disease was first observed in 1966 in Kenya but has now spread in all rice-growing countries of Sub-Saharan Africa. In Tanzania, its distribution has been restricted to the major rice-growing regions. However, the knowledge on RYMV genetic diversity relies on a limited number of coat protein sequences. Previous studies revealed the presence of the phylotypes S4lv, S4lm and strain S5 in Mwanza, Mbeya and Morogoro regions, respectively, and strain S6 in Kilimanjaro region and Pemba Island. Surveys were conducted during the cropping seasons 2013-2014 in eight rice-growing regions of Tanzania to determine geographical variations and phylotypes of RYMV and the influence of environment factors on its distribution and diversity. A total of 185 rice fields were surveyed. Results indicate that prevalence, severity and phylotypes of RYMV varied significantly with rainfall intensity, temperature and relative humidity (P ≤ 0.01). The highest prevalence was found in Morogoro (82%), Mbeya (80%) and Arusha (67.33%) regions whereas Kigoma (9.33%), Rukwa (11.33%) and Shinyanga (18.67%) had the lowest RYMV prevalence. In each region, RYMV prevalence was higher in 2014 than in 2013. The phylotypes S4lm and new determined phylotypes (S6c and S6w) were highly adapted to low temperature (13.3˚C) and rainfall (13.7 mm) areas. For the first time, strains from the phylotype S4ug were found outside Uganda, in Kilimanjaro region. Strain S4lv (phylotype Lake Victoria) was found for the first time in Arusha region. The strain S4lm was found in Mbeya, Morogoro and Rukwa regions. Strains S4lm and S4lv were also found in Shinyanga and Kigoma regions, respectively. The strain S5 was still restricted to Morogoro but extended to new locations such as Ulanga district. Strain S6 was found in several new areas and new phylotypes of S6 (S6c and S6w) are reported in this study.
Keywords:
Rice Yellow Mottle Virus, Prevalence, Severity, Environmental Factors, Tanzania

1. Introduction
Rice (Oryza sativa L.) is the staple food of more than half of the world’s population [1] . In Africa where consumption has grown faster than production, yield increase in one of the major issue. Indeed, rice cultivation is facing several biotic and abiotic constraints. Rice yellow mottle virus (RYMV) provokes the most important viral disease in most rice-growing countries of Africa [2] [3] . The typical symptoms of the disease are mottle and yellowing of the leaves, stunting of the plants, reduced tillering, poor panicle exertion and sterility. Yield losses ranging from 20% - 100% on susceptible rice cultivars due to variation levels of RYMV disease incidence and severity has been reported [2] [4] . In Tanzania, the second rice producer in Eastern Africa, rice is the second most important staple crop. However, yield remains low, in the range of 1 - 1.5 tons/ha, due to several constrains such as RYMV [4] [5] [6] [7] . First reported in 1993 in Mkindo irrigation project in Morogoro region [8] , RYMV was detected two years later in the major rice producing regions [9] . The virus was found in 100%, 50% and 8% of the fields in Morogoro, Mbeya, Mwanza and Shinyanga regions, respectively. The host range of RYMV is restricted to cultivated rice and few wild grass species [10] . RYMV is transmitted mechanically mainly by sap contact and insect vectors (family Chrysomelidae) [11] . However, the virus is not seed transmitted [12] .
The virus belongs to the genus Sobemovirus and is characterized by an icosahedral particle of 30 nm in diameter that contains one single strand positive sense genomic RNA [13] . RYMV has a high level of genetic diversity, spatially structured and which gradually decrease from West to East [14] . Several serotypes and strains of RYMV have been identified in various geographical locations [10] [15] . Six major strains have been described [9] [14] , three (S4, S5 and S6) were found in East Africa. Tanzania has been reported as one of the biodiversity hotspot of RYMV [15] . RYMV was first observed in 1966 in Kenya [11] and later reported in nearly all rice-growing countries of Sub-Saharan Africa [10] . In Tanzania, the first incidence of RYMV was reported in 1993 in Mkindo irrigation project, Morogoro region [8] . Fields studies in Pemba Island from the Zanzibar archipelago showed RYMV symptomatic plants. Two different serotypes were detected using ELISA assays in various geographical areas: one in Morogoro region and Pemba Island and one in Mbeya, Mwanza and Shinyanga regions, and later named as Ser5 and Ser4, respectively. Genetic analyses of the coat protein (CP) gene confirmed the molecular basis of the two serotypes [9] [16] which also allowed the distinction of two phylotypes in the serotype Ser4: phylotypes S4lv and S4lm [9] . Coat protean sequences from Mwanza region (close to Lake Victoria) clustered together in the phylotype S4lv while the phylotype S4lm gathered sequences from Mbeya region (close to the Lake Malawi). In addition, two different strains were defined from the serotype Ser5: strains S5 and S6 with CP sequences from Morogoro region and Pemba Island, respectively [9] . In 2005, the strains S4 and S6 were first detected in Morogoro and Kilimanjaro regions, respectively, whereas the strain S5 was restricted to Morogoro region [17] . Since this publication, very few data on RYMV in Tanzania have been available.
Rice yellow mottle virus is widely distributed in all rice growing conditions in Africa but the role of environmental factors on the virus distribution and diversity is poorly investigated. However, the environment and the climate change can influence host plant growth and susceptibility, pathogen and vector reproduction, dispersal, survival and activity as well as host-pathogen interaction [18] . Few data suggested an influence of the environmental factors such as temperature and rainfall on RYMV epidemics. More epidemics occur in irrigated areas and, also to a lesser extent, in lowland rice during the rainy season [19] . RYMV was reported in every region in Madagascar except in the central highlands [20] . In addition, the genetic diversity is spatially structured including strains from West and East Africa which are different, even at a country scale. For instance in Cote d’Ivoire, strains located in savanna and forest areas are different [21] .
Therefore, today while rice intensification is a priority, the geographic diversity and epidemiological surveys of the virus is needed in Tanzania (1) to detect RYMV presence in more rice-growing regions, (2) to precisely quantify the viral prevalence and severity in each agro-ecological zones and (3) to determine the geographical variation and spatial distribution of the RYMV strains and phylotypes in the Tanzanian biodiversity hotspot. Finally, the relationship between RYMV distribution, genetic diversity and environmental variability was investigated.
2. Materials and Methods
2.1. Description of the Study Area
Field surveys were carried out in farmers’ rice fields in selected rice growing areas in the Lake zone (Shinyanga and Kigoma regions), the Southern Highland zone (Rukwa and Mbeya regions), the Eastern zone (Morogoro and Pwani regions) and in the Northern zone (Kilimanjaro and Arusha regions) in Tanzania. The study was carried out in two cropping seasons, March to May 2013 and April to May 2014 to determine the prevalence and severity of RYMV strains on rice. The mean annual rainfall in the study areas ranged from 500 to 1400 mm/year with minimum and maximum temperatures of 13˚C to 23˚C and 25˚C to 32˚C, respectively. The rice fields were located in various altitudes ranging from 25 to 1326 m above sea level.
2.2. Field Surveys
2.2.1. Distribution, Prevalence and Severity of RYMV
The surveyed areas were selected based on their history of rice production, diverse ecological backgrounds and reported frequent occurrence of RYMV. The surveys covered a total of 185 fields. Samples were collected using the W-walk sampling method as described by Ardales et al. [22] to better understand the relationships between strains and environmental factors within geographical areas. Geographical position system (GPS) coordinates, altitude and weather data (rainfall, temperature, RH and wind speed) obtained from Tanzania Meteorological Agency, for the period from 2013 to 2014 were recorded and collected, respectively. Tanzanian administrative boundaries overlaid with collected GPS data using ArcGIS Software Package to display location of the RYMV disease surveyed areas (Figure 1). The GPS points were then interpolated to create a geographical distribution map of Rice yellow mottle virus disease (Figure 6). Altitudes of all locations visited [Mto wa Mbu, Kiwowo, Maweni (Arusha), Kyela, Igurusi, Chunya (Mbeya), Woria, Ndungu, Lower Moshi (Kilimanjaro), Madaganya, Dakawa, Vigoi (Morogoro), Mkuti, Kibondo, Kasulu (Kigoma), Shinyanga, Kahama (Shinyanga), Kibaha, Bagamoyo (Pwani), Mpanda, Sumba-

Figure 1. A geographic map showing locations (colored in green and red circles) of Rice yellow mottle virus field surveyed areas (2013 and 2014 rice growing seasons).
wanga (Rukwa)] were recorded. Three hundred and fourteen isolates of RYMV were obtained from diseased plants showing characteristic leaf mottling symptoms. The diseased rice leaf samples were collected in paper bags labeled and transported to the African Seed Health Center laboratory, Sokoine University of Agriculture (SUA), Tanzania. These samples were kept into the freezer at −20˚C until when used for further analysis.
The percentage of rice plants with RYMV disease symptoms over the total plant area of 1 m x 1 m was determined. In each locality, three counts were taken diagonally per field and 5 to 7 fields per location were assessed at an interval of 10 to 15 km per location following the procedures of Bekeko et al. [23] . Within each quadrat, RYMV prevalence was calculated using the formula described by Nwilene et al. [24] as follows: RYMV prevalence = Number of rice plants with disease/ Total number of rice plants (health and diseased) x 100.
Scores for disease severity were recorded using the 1?9 scale of standard evaluation system for rice [1] : where: 1 = no symptoms, 3 = sparse dots or streaks, 5 = general mottling of the leaves, 7 = yellowing and stunting and 9 = necrosis and sometimes plant death. Disease severity (S) was calculated according to the procedures described by Finninsa [25] as:
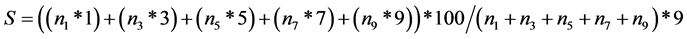 where n1, n3, n5, n7, n9 represent the number of leaves scored 1, 3, 5, 7 and 9, respectively.
where n1, n3, n5, n7, n9 represent the number of leaves scored 1, 3, 5, 7 and 9, respectively.
The RYMV disease prevalence and severity data were analysed as One Way Randomized Block with three replications using GenStat Software Package. The data were subjected to arcsine transformation to normalize the data before analysis [26] . A constant value (0.5) was added to each observation, before taking arcsine transformation [27] . The mean separation test based on the different locations and years tested for disease prevalence and severity were done using the Tukey’s Multiple Range Test at P ≤ 0.05.
2.3. Laboratory Studies
2.3.1. Rice Yellow Mottle Virus Detection and Serotyping
A sample was prepared by grinding 0.1 g leaf in 1 ml of phosphate buffer saline with 0.5% Tween-20 (PBST 1X) in a 2 ml tube at the ratio of 1:10 (w/v) using TissueLyser II. Direct antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA) using polyclonal antibodies raised against a strain from Madagascar were used to detect RYMV in the rice leaf samples collected during the survey [14] . Two polystyrene micro plate wells were used to score the optical density (OD) values at 405 nm using ELISA spectrometer plate reader and the mean values were used for analysis. The virus titer was considered significant when the optical density values were two-fold greater than the mean value of the negative control. Positive leaf samples for RYMV were selected and stored at −20˚C for molecular analyses. Triple-antibody- sandwich (TAS)-ELISA was used to assess the immunological profile of the RYMV isolates as described by Pinel et al. [28] . Two monoclonal antibodies (MAbs) (E and M) were used to distinguish serotypes Ser4 and Ser5 which are wide spread in East Africa. Antibody M was non discriminative to each RYMV strain and was used as a reference to quantify the overall virus titer independently of the serological properties of the isolates.
2.3.2. Viral Coat Protein Gene Sequencing
Total RNA of RYMV was extracted from frozen infected rice leaves using the Rneasy Plant Mini Kit (Qiagen) method as described by Pinel et al. [29] . Viral suspension were collected in 2 ml eppendorf tube with sterile steel beads, frozen in liquid nitrogen and ground with high speed TissueLyser II mechanical shaker for 1 min at 30 rpm. The RTL lysis buffer was added, mixed by vortexing then incubated in water bath at 56˚C for 2 minutes and centrifuged at 7000 rpm for 7 minutes. Tissues were separated by 225 μl of 100% ethanol followed by spinning at 10,000 rpm for 1 min, and then the supernatants were transferred into 2 ml Eppendorf tubes. Proteins of RYMV were removed by adding 700 μl RW1 and 500 μl RPE buffer, respectively and separately, centrifuged as above then the supernatant liquid was discarded and transferred into sterile 2 ml tubes. Ribonucleic acid was washed in 500 μl RPE buffer by spinning at 13,000 rpm for two minutes. Nucleic acids were eluted by 30 μl RNase free water directly to the spin column membrane and placed into clean sterile 1.5 ml tubes then centrifuged at 10,000 rpm for 1 min at 25˚C. The obtained RNAs were stored in the freezer at −20˚C for RT-PCR amplification.
A reaction of RT-PCR to transcribe and amplify the coat protein gene consisted of a total volume of 50 μl of a mixture per one sample was done as described by Pinel et al. [29] . For reverse transcriptase (RT)- complementary deoxyribonucleic acid (cDNA) synthesis, the reaction mixture of 7.5 µl of total RNA, 1 µl antisense primer RYMV II at 100 µM and 1.5 µl Rnase free water in a PCR-tube was done and denatured at 70˚C for 5 min. Then a mixture of 6 µl Rnase free water, 2 µl dNTPs 5 mM, 5 µl Buffer RT x5 (promega) x1 final, 1 µl M-MLV-RT (promega) 200U/µl and 1 Rnase inhibitor 20U/µl (promega) per one sample was added into the PCR tube containing 10 µl of first reaction making a total volume of 25 µl. The mixture was incubated at 42˚C for 1 hour. The PCR mixture included 5µl buffer PCR x10, x1 final, 2 µl dNTP 5 mM chacum 2.0 mM final, 1µl antisense primer RYMV II 10 µM, 3 µl sense primer RYMV III 10 µM, and 1 µl dynazyme, 35.5 µl SDW and 2.5 µl RT reaction. The primer set consisted of 5'-CAAAGATGGCCAGGAA-3' (forward primer) and 5'-CTCCCCCACCCATCCCGAGAATT-3' (reverse primer) were used to amplify the coat protean (CP) gene of RYMV. The amplification process involved initial denaturation at 94˚C for 5 minutes followed by 30 cycles (Denaturation at 94˚C for 3 minutes, annealing at 55˚C for 30 seconds, elongation at 72˚C for 1 minute), then final extension and stop at 72˚C for 10 minutes. The products were confirmed using 1% agarose gel [in 0.5x Tris-acetate-Ethylenediaminetetraacetic acid (TAE) buffer] electrophoresis and sequenced by Biomed Company.
2.3.3. Phylogenetic Analysis for Identification of RYMV Strains and Phylotypes
The 48 sequences of the coat protein gene were aligned with 7 reference strains retrieved from the Genbank using CLUSTAL X with default parameters [29] . Neighbour-joining tree was calculated from the pairwise amino acid sequence distances between the aligned coat protein genes of 53 RYMV strains from Tanzania and 2 RYMV strains from Uganda. The phylogenetic tree was constructed using the maximum likelihood method with default parameters in SeaView software [30] [31] .
3. Results
3.1. Variation in RYMV Prevalence and Severity in Tanzania
The surveys covered two cropping seasons, March to May 2013 and April to May 2014, for a total of 185 fields distributed in 8 rice-growing regions of Tanzania. Morogoro region was selected to be more intensively surveyed, as a major rice production area and a RYMV biodiversity hotspot.
Rice yellow mottle virus was found in all surveyed regions including Kigoma, Rukwa, Pwani, Arusha in Western Lake zone, Plateau, Coast and Northern Highland zone of Tanzania. The analysis of variance for disease prevalence for the two rice growing seasons showed statistically highly significant differences (P < 0.001) across locations (Table 1). The mean disease prevalence of RYMV across regions varied from 9.33% in Kigoma region (Western Lake zone) to 82% in Morogoro region (Coastal zone), respectively, over the two rice growing seasons (Table 1). The highest prevalence of RYMV was found in Morogoro, Mbeya and Arusha regions whereas Kigoma, Rukwa and Shinyanga showed the lowest prevalence. The RYMV prevalence in Pwani and Kilimanjaro regions was

Table 1. Disease prevalence and severity of Rice yellow mottle virus for surveyed regions 2013 and 2014 cropping seasons.
*Values are means of three replicates in two separate seasons (2013 and 2014). Values were Arcsine transformed before analysis. Numbers followed by the same letters in a column are not significantly different at P < 0.05, using Tukey’s Multiple Range Test. *** = highly significantly different (P < 0.001).
intermediate. In each region, RYMV prevalence was higher in 2014 than in 2013. The annual variability of the RYMV prevalence in each region explained 90% of the variability (Figure 2). The lowest RYMV variability was found in Shinyanga region whereas Pwani and Morogoro regions showed the highest variability.
Disease severity was assessed using the standard evaluation system (SES) for RYMV symptoms. The maximum value of RYMV severity was observed at 55% in Morogoro region in 2014. Rice yellow mottle virus severity varied from 15 to 55% depending on the region and the year (Table 1). The trends of RYMV severity in Pwani and Kilimanjaro regions were the same and intermediary as for RYMV prevalence. Although RYMV severity was globally higher in 2014 than in 2013, exceptions were observed for Arusha and to a lesser extent Shinyanga region. However, the annual variability of the RYMV severity in each region explained 45% of the variability (Figure 2). The lowest RYMV variability was again found in Shinyanga region whereas Morogoro region showed the highest variability.
Weather parameters (temperature, wind speed, rainfall and RH) were related to prevalence, severity and distribution of RYMV phylotypes within the fields and locations (Figure 3). In 2013, the highest RYMV disease prevalence and severity were recorded in Mbeya region (Southern highland zone), receiving a total rainfall of 167 mm and temperature ranging from 16.8 to 27.7˚C (Figure 3(a)). The RH value of 70.4%, wind speed of 4 km/h and temperature range of 20 to 31˚C were associated with high RYMV prevalence and severity in Morogoro region (Coastal zone). A similar trend was observed in 2014 rice cropping season (Figure 3(b)). The S4lm phylotype and S6 strain of RYMV were highly associated to low temperature (13.3˚C) and rainfall (13.7 mm), respectively. The highest RYMV disease prevalence and severity were observed only in areas with strong wind conditions such as mean wind speed of 9.3 and 18.5 km/h recorded

Figure 2. Rice yellow mottle virus prevalence and severity in Tanzania for 2013 and 2014 cropping seasons. Linear regressions were calculated and determination coefficients were indicated when p-value < 0. 05.

Figure 3. Relationship between weather and Rice yellow mottle virus disease parameters in studied regions in Tanzania (3A) 2013 cropping season (3B) 2014 cropping season. The RYMV strains and phylotypes detected in each region were indicated at the center. Phylotypes: S4lv = Strain 4-Lake Victoria, S4ug = Strain 4-Uganda, S4lm = Strain 4-Lake Malawi, S6c = Strain 6-coast area, S6w = Strain 6-wide.
in Pwani and Arusha, respectively (Figure 3(b)). The high prevalence and severity in Mbeya, Morogoro and Pwani regions as shown in Figure 3, were associated with high rainfall, but in Arusha region were probably associated by wind speed and temperature.
3.2. The Effect of Altitude on the Prevalence and Severity of RYMV Strains and Phylotypes
The rice fields in this study were located in various altitudes ranging from 25 m to 1326 m above sea level. The results have shown that the prevalence and severity of RYMV varied with altitudes (P ≤ 0.05) (Figure 4). The results also showed that, RYMV prevalence and severities were slightly influenced by altitude. This is because they were high in Arusha and Mbeya regions where altitudes are high and also in Morogoro and Pwani regions where the altitude is low (Figure 4).

Figure 4. The effect of altitude on the prevalence and severity of Rice yellow mottle virus disease in the studied regions in Tanzania.
The highest disease prevalence (81.3%, 88.9%, 90.3% and 95%) were recorded in Chunya (Mbeya-Highland), Madaganya and Vigoi (Morogoro-Coast belt) and Kyela (Mbeya-Highlands) located at altitudes of 524 m, 505 m, 480 m and 529 m above sea level, respectively (Figure 4). The highest RYMV disease severity was also recorded in the same regions. However, rice fields in Kasulu, Kigoma (Lake Zone), located at 1319 m, had the lowest RYMV prevalence (15%) and severity (13%) of RYMV.
3.3. Serological Characterization of RYMV Isolates
A total of 193 samples collected from surveyed areas in Tanzania were positive for RYMV in Double Antibody Sandwich Enzyme-Linked immunosorbent Assay (DAS ELISA). RYMV isolates were serotyped using two monoclonal antibodies (MAbs M and E) in TAS ELISA (Table 2). The isolates belonged to two serotypes, Ser 4 and Ser 5 which have already been described in Tanzania. The serotype Ser 4 was detected in all regions except in Pwani (Coast). Ser 4 of RYMV was detected for the first time in this study in Arusha, Kilimanjaro, Kigoma and Rukwa regions. The second serotype (Ser5) was also found in each surveyed region except in Rukwa (Plateau). These isolates showed negative reactions with all MAbs (M and E) and were designated as Ser 5. Several variants of the two serological profiles (Ser 4 and Ser 5) were distinguished by their unusual reactions with both MAbs. Serotype Ser 5 was recorded for the first time for one isolate from Kigoma (Lake Zone) and two isolates from Mbeya (Southern Highlands) where S6c and S6w phylotypes were found.
3.4. Identification of RYMV Strains and Phylotypes
To characterize the RYMV diversity, the coat protein genes of 48 strains, 21 from the serotype Ser4 and 27 strains of the serotype Ser5, were sequenced and
Table 2. Serotypes of Rice yellow mottle virus isolates from the study locations.
Absorbance values in ELISA were coded as follows: ‘0’ ≤ 0.30, 0 ≤ 31‘1’0 ≤ 60, ‘0 ≤ 61‘2’1 ≤ 20, 1 ≤ 21‘3’ ≤ 1.80, ‘4’ ≥ 1.81.
analyzed by the maximum likelihood method (Figure 5 Figure 6 and Supplement Table 1). Sequences of the strains from the serotype Ser4 clustered in the strain S4 which is divided into three phylotypes (S4ug, S4lv and S4lm). For the first time, strains from the phylotype S4ug were found outside Uganda, in Kilimanjaro region (Northern Highlands of Tanzania). Strain S4lv (phylotype Lake Victoria) was found for the first time in Arusha region. The strain S4lm (first reported near Lake Malawi) was found in Mbeya, Morogoro and Rukwa regions. Surprisingly, the strains S4lm and S4lv were detected in Shinyanga and Kigoma regions, respectively. The strain S5 is still restricted to Morogoro but extended to new locations such as Ulanga district, where it had not been reported previously. Rice yellow mottle virus strains which belonged to S6 were found in new areas, in Woria division-Kilimanjaro, Kigoma and Mbeya regions where they have not been reported previously. They were also found in Pwani, Morogoro and Arusha regions. Furthermore, group S6 was made up of twenty two sequences of strains from Ser5 clustered in the strain S6 group which divided into two new phylotypes (S6w-wide distributed and S6c-coast area). The environmental factors such as temperature, rainfall and RH may play role and trigger the geographic distribution of phylotype variation within the genetic structure of the RYMV populations.
Phylotypes of RYMV were S4lm (strain Tz408), S6w (strain Tz303) and S6c (Tz305) in Mbeya region (Southern Highlands), while S4lm (strains Tz441, Tz483, Tz526, Tz554), S5 (Tz416, Tz429, Tz445, Tz449, Tz450, Tz454, Tz460, Tz461), S6c (strains Tz452, Tz523, Tz510, Tz539, Tz608, Tz619, Tz651, Tz801), S6w (strains Tz463, Tz486) were identified in Morogoro region (Coastal zone). Phylotype S4lv (strains Tz421, Tz507, Tz516, Tz520) and S6c (strains Tz504, Tz512, Tz515) were identified in Arusha region (Northern Highland zone)

Figure 5. Phylogeny of Rice yellow mottle virus in Tanzania. Phylogenetic tree reconstructed by the maximum likelihood method from the ORF4 sequences of the 54 strains including five reference strains. The strains are indicated by vertical bars.
(Figure 5). Rice yellow mottle virus phylotypes varied across geographical areas in Morogoro region (Coastal zone) compared to other regions and these similar trends prevailed in both seasons (Figure 5).
Furthermore, results from this study have indicated that the coat protein gene of the 28 strains sequenced had the same length of 720 bp, except 6 strains (Tz429, Tz460, Tz461, Tz416, Tz445, and Tz446), which had a long sequence of

Figure 6. Location of Rice yellow mottle virus strains in Tanzania indicating the surveyed regions and detected strains.
723 bp and one isolate Tz554 had a short sequence of 718 bp. The long sequence was due to extra nucleotides (AGA for Tz554, Tz460, Tz461 and AGG for Tz416, Tz445 and AAG for Tz446) which coded for an extra amino acid arginine at position 56.
4. Discussion
In 2013 and 2014, rice in Tanzania was seriously affected by RYMV with 40% mean prevalence, although spatio-temporal variations were observed. The RYMV prevalence data found in this study confirmed the previous report that defined Morogoro and Mbeya regions as hotspot areas of the disease compared to Shinyanga region [9] . Nevertheless, in Mbeya region the number of infected fields has been found to be lower than in Morogoro region (50% vs 100% in 2005). The RYMV prevalence was measured as percentage of symptomatic plants onto total number of plants in the studied quadrats while severity was assessed on the symptomatic plants using a standard scale. In Shinyanga, the number of symptomatic plants was low, may be due to unfavorable conditions for RYMV in this region. Symptoms variability in the field would be associated with several factors such as changing of rice cultivars, poor agricultural practices, phenological and physiological status at the survey period, inoculum doses and transmission frequency caused by vector population. These factors may also contribute to the annual variability of RYMV disease in each surveyed region that statistically indicated only 45% of the total variability.
However, the rice fields surveyed in the current study were surrounded by Mountains and forest with thick grass vegetations that may have been the source of RYMV insect vectors and created conditions favorable for spread of RYMV. This observation is consistent with earlier and recent reports that natural sources of RYMV infection and spread of RYMV to rice crops were present in a few water-dependent wild grasses, such as Echinochloa colona and Penicum repens [24] [36] . The spread of RYMV and its diversity in Tanzania may be associated with an agro-ecological change, extension of rice cultivation to new areas and modification of landscape ecology [37] . Phylogeographic structure has been reported to be highly dependent on landscape ecology [38] . However, geographical adaptation of RYMV strains found in this study may be caused by climatic factors including temperature and RH. The RYMV prevalence and severity variability were observed between the surveyed years. However, the highest RYMV prevalence and severity of RYMV were recorded in Morogoro region. In Arusha, compared to other regions, RYMV severity was high in 2013 than in 2014.
Variation of RYMV disease prevalence and severity could be attributed to changes in epidemiological factors (environmental conditions), rice plant resistance and strain aggressiveness. However, rainfall and temperature variations might have influenced RYMV insect vector population for the virus distribution in Tanzania. The RYMV disease incidence has been reported to vary between rain and dry seasons [19] . Traoré et al. [39] reported that the epidemics of RYMV were influenced by rice growing environments. However, RYMV is vector-transmitted disease and weather conditions may affect dynamics and population of vectors transmitting the disease [40] . Bakker [11] reported several beetles with potential of transmitting RYMV. Emerging of new strains and variants has been reported to increase RYMV disease incidence and severity [33] .
Serological characterization of RYMV strains confirmed the presence of two serotypes in Tanzania. The serotype Ser4 showed the widest distribution and serotype Ser5 was mainly detected in regions where the serotype Ser4 was present showing spatial overlaps between the two serotypes. Molecular typing of RYMV strains suggested other overlaps between the phylotypes S4lm and S4lv due to the emergence of these phylotypes into new areas. Rice yellow mottle virus strains and phylotypes were invaded to Southern and Northern Tanzania from the Lake Zone and to the Lake Zone from Southern highland in Tanzania. These results are consistent with those of Traore´ et al. [41] in Cote d’Ivoire where the competition between strain S1 and S2 has been occurred. Fitness differences have been reported to create changes in the RYMV epidemiological dynamics [41] . The first report in Tanzania (Kilimanjaro region) of the strain S4ug, a new invading strain in Uganda is an evidence of such spatial evolution. The transmission of RYMV phylotypes from their geographical origin may probably be influenced by both geographical and seasonal variations. However, new surveys are needed in the neighboring regions to follow the overlaps with the strain S4lv and to determine the circulating strains in Kenya. Morogoro region had a large RYMV diversity with three different strains, followed by Arusha, Kilimanjaro, Pwani and Shinyanga, both with two different strains. Several distinct RYMV strains assessed using the sequences of their CP gene showed variability in their geographical origins [28] . Serological differences have been reported between the RYMV strains from Ivory Coast and Kenya [41] . Furthermore, Kanyeka et al. [17] reported long sequence length of isolates from Tanzania with extra nucleotides (CGC) which code for an extra amino acid arginine at position 60. A shorter sequence of an isolate Tz554 in this study was due to missing nucleotides for an amino acid arginine at different positions 397, 409, 422, 428, 442 and 468. This feature has been reported to be typical of S5 strain [14] .
Strain S5 isolates collected from Morogoro region showed variation according to the area of its origin. Strain S5 was restricted only in the Kilombero Valley and widely spread in Kilombero and Ulanga districts. The strain S5 was reported by Kanyeka et al. [17] to be restricted in a small area (Ifakara ward) of Kilombero district in Kilombero Valley but it has now spread to new areas. Abubakar et al. [9] reported S5 strain to be found in areas with high disease incidence (hotspot areas). The presence of three East-African strains in Kilombero Valley, Morogoro region, a high diversity hotspot area of RYMV in Tanzania, offered the possibility to study the micro geographical variation of RYMV in order to better understand the relationship between strains and compare to what is found in other area. Targeting a hot spot area may allow to clearly know the structure of the RYMV population in terms of strains and resistant-breaking (RB) strains and the relationship between the strains and pathological variants. Previous studies reported the predominantly occurrence of phylotype S4lv in regions around Lake Victoria [17] , but in this study, S4lv was found in Northern zone of Tanzania, Arusha region. Rice yellow mottle virus strain S6 was found in several new areas and new phylotypes of S6 (S6c and S6w) were determined in this study. The emergence of RYMV phylotypes in the surveyed geographic regions of Tanzania may increase and spread to other rice growing areas. This may also support significant populations of RYMV insect vectors within the country.
The surveyed environmental conditions favour existence of RYMV and support a large number of vectors and RYMV reservoirs, thus resulting in high prevalence of the RYMV. Rice yellow mottle virus was introduced into Madagascar by long distance spread (>400 km) with confirmation of a strong bottleneck effect [37] . The insect vectors may provide long distance transmission of RYMV [38] through existence of continuous natural vegetation regional-wise. Phylotype S4lm has been reported to spread from Lake Malawi regions into eastern Tanzania, despite the mountain chain which separates the two regions [17] . However, temperature, relative humidity, rainfall and wind speed are considered as the most important factors that favor development and spread of plant diseases [18] and completion of vector life cycles [37] . Rice yellow mottle virus is transmitted by wind mediated leaf contact [42] and through guttation fluid [10] and irrigation water [36] [43] that could explain high prevalence observed in high wind speed areas. These new results obtained in this study, call for further studies that should take into account intra-regional variability of rice agro-ecosystems, other factors from host (cultivar, physiology, reservoirs), vector (insect population, transmission level) and environment (as direct or indirect factor). The spatio-temporal model recently built by Trovao et al. [44] may be applied to determine the relationship between the RYMV hyper-variable epidemics and environmental factors.
Acknowledgements
The authors acknowledge financial and material support from the Innovative Agricultural Research Initiative (iAGRI) and the Institut de Recherche pour le Developement (IRD) and Professor Robert B. Mabagala, the Sokoine University of Agriculture, Tanzania and Denis Fargette and Eugénie Hébrard, IRD, France, for helpful discussions. The assistance from Agnes Pinel-Galzi, IRD, France laboratory technician is acknowledged.
Cite this paper
Hubert, J., Lyimo, H.J.F. and Luzi-Kihupi, A. (2017) Geographical Variation, Distribution and Diversity of Rice Yellow Mottle Virus Phylotypes in Tanzania. American Journal of Plant Sciences, 8, 1264-1284. https://doi.org/10.4236/ajps.2017.86084
References
- 1. International Rice Research Institute, Eds. (2002) Standard Evaluation System for Rice. 5th Edition, Inger. Genetic Resources Center, IRRI, Manila, Philippines, 27 p.
- 2. Kouassi, N.K., N’Guessan, P., Albar, L., Fauquet, C.M. and Brugidou, C. (2005) Distribution and Characterization of Rice Yellow Mottle Virus: A Threat to African Farmers. Plant Disease, 89, 124-133.
https://doi.org/10.1094/PD-89-0124 - 3. Matsumura, K., Hijmans, R.J., Chemin, Y., Elvidge, C.D., Sugimoto, K., Wu, W., Lee, Y. and Shibasaki, R. (2009) Mapping the Global Supply and Demand Structure of Rice. Sustainability Science, 4, 301.
https://doi.org/10.1007/s11625-009-0077-1 - 4. Luzi-Kihupi, A., Zakayo, J.A., Tusekelege, H., Mkuya, M., Kibanda, N.J.M., Khatib, K.J. and Maerere, A. (2009) Mutation Breeding for Rice Improvement in Tanzania. In: Induced Plant Mutations in the Genomics Era, Food and Agriculture Organization of the United Nations, Rome, 385-387.
- 5. Awoderu, Y.A. (1991) Rice Yellow Mottle Virus Situation in West Africa. Journal of Basic Microbiology, 31, 91-99.
https://doi.org/10.1002/jobm.3620310204 - 6. Mghase, J.J., Shiwachi, H., Nakasone, K. and Takahashi, H. (2010) Agronomic and Socio-Economic Constraints to High Yield of Upland Rice in Tanzania. African Journal of Agriculture Research, 5, 150-158.
- 7. Taylor, D.R., Fofie, A.S. and Suma, M. (1990) Natural Infection of Rice Yellow Mottle Virus Disease (RYMV) on Rice in Sierra Leone. International Rice Research Newsletter, 15, 5-19.
- 8. Kanyeka, Z.L., Kibanda, J.M. and Mbapila, J. (1996) Rice Yellow Mottle Virus in Tanzania. Proceedings of the Potential and Constraints for Improvement of Rice Cultivation Seminar, Kilimanjaro Agricultural Training Center, Moshi (KATC), Tanzania, 11-15 March 1996, 17-22.
- 9. Abubakar, Z., Ali, F., Pinel, A., Traoré, O., N’Guessan, P., Notteghem, J., Kimmins, F., Konaté, G. and Fargette, D. (2003) Phylogeography of Rice Yellow Mottle Virus in Africa. Journal of General Virology, 84, 733-743.
https://doi.org/10.1099/vir.0.18759-0 - 10. Abo, M.E., Sy, A.A. and Alegbejo, S.M. (1998) Rice Yellow Mottle Virus (RYMV) in Africa: Evolution, Distribution, Economic Significance and Sustainable Rice Production and Management Strategies. Journal of Sustainable Agriculture, 11, 85-111.
https://doi.org/10.1300/J064v11n02_08 - 11. Bakker, W. (1970) Rice Yellow Mottle, a Mechanically Transmissible Virus Disease of Rice in Kenya. Netherlands Journal of Plant Pathology, 76, 53-63.
https://doi.org/10.1007/BF01974433 - 12. Konaté, G., Sarra, S. and Traoré, O. (2001) Rice Yellow Mottle Virus Is Seedborne but Not Seed Transmitted in Rice. European Journal of Plant Pathology, 107, 361-364.
https://doi.org/10.1023/A:1011295709393 - 13. Tamm, T. and Truve, E. (2000) Sobemovirus (Minireview). Journal of Virology, 74, 6231-6231.
https://doi.org/10.1128/JVI.74.14.6231-6241.2000 - 14. Fargette, D., Pinel, A., Halimi, H., Brugidou, C., Fauquet, C. and Van Regenmortel, M. (2002) Comparison of Molecular and Immunological Typing of Isolates of Rice Yellow Mottle Virus. Archives of Virology, 147, 583-596.
https://doi.org/10.1007/s007050200008 - 15. Mpunami, A., Ndikumana, I., Hubert, J., Pinel-Galzi, A., Kibanda, N., Mwalyego, F., Tembo, P., Kola, B., Mkuya, M., Kanyeka, Z., Mutegi, R., N’chimbi Msolla, S., Njau, P., Séré, Y., Fargette, D. and Hébrard, E. (2012) Tanzania, Biodiversity Hotspot of Rice Yellow Mottle Virus. Arusha, Tanzania, 97-101.
- 16. Banwo, O.O., Alegbejo, M.D. and Abo, M.E. (2004) Rice Yellow Mottle Virus Genus Sobemovirus: A Continental Problem in Africa. Journal of Plant Protection Science, 40, 26-36.
- 17. Kanyeka, Z.L., Sangu, E., Fargette, D., Pinel-Galzi, A. and Hébrard, E. (2007) Distribution and Diversity of Local Strains of Rice Yellow Mottle Virus in Tanzania. African Journal of Crop Science, 15, 201-209.
- 18. Jones, R.A.C. (2016) Future Scenarios for Plant Virus Pathogens as Climate Change Progresses. Advances in Virus Research, 95, 87-147.
- 19. Traoré, O., Galzi-Pinel, A., Poulicard, N., Hébrard, E., Konaté, G. and Fargette, D. (2008) Rice Yellow Mottle Virus Diversification Impact on the Genetic Control of RYMV. Plant Disease Journal, 5, 1-4.
- 20. Rakotomalala, M., Pinel-Galzi, A., Albar, L., Ghesquière, A., Rabenantoandro, Y., Ramavovololona, P. and Fargette, D. (2008) Resistance to Rice Yellow Mottle Virus in Rice Germplasm in Madagascar. European Journal of Plant Pathology, 122, 277-286.
https://doi.org/10.1007/s10658-008-9282-5 - 21. N’Guessan, P., Pinel, A., Caruana, M.L., Frutos, R., Sy, A., Ghesquière, A. and Fargette, D. (2000) Evidence of the Presence of Two Serotypes of Rice Yellow Mottle Sobemovirus in Côte d’Ivoire. European Journal of Plant Pathology, 106, 167-178.
https://doi.org/10.1023/A:1008792109954 - 22. Ardales, E.Y., Leung, H., Vera Cruz, C.M., Mew, T.W., Leach, J.E. and Nelson, R.J. (1996) Hierarchical Analysis of Spatial Variation of the Rice Bacterial Blight Pathogen across Diverse Agroecosystems in the Philippines. Phytopathology, 86, 241-252.
https://doi.org/10.1094/Phyto-86-241 - 23. Bekeko, Z., Hussien, T. and Tessema, T. (2012) Distribution, Incidence, Severity and Effect of the Rust (Puccinia abrupta var. partheniicola) on Parthenium hysterophorus L. in Western Hararghe Zone, Ethiopia. African Journal of Plant Science, 6, 337-345.
https://doi.org/10.5897/AJPS12.040 - 24. Nwilene, F.E., Traore, A.K., Asidi, A.N., Sere, Y., Onasanya, A. and Abo, M.E. (2009) New Records of Insect Vectors of Rice Yellow Mottle Virus (RYMV) in Cote d’Ivoire, West Africa. Journal of Entomology, 6, 198-206.
https://doi.org/10.3923/je.2009.198.206 - 25. Finninsa, C. (2003) Relationship between Common Bacterial Blight Severity and Bean Yield Loss in Pure Stand and Bean-Maize Intercropping Systems. International Journal of Pest Management, 49, 177-185.
https://doi.org/10.1080/0967087021000049269 - 26. William, H.A., Darrell, J.C. and Girish, B. (1990) Use of the Arcsine and Square Root Transformations for Subjectively Determined Percentage Data. Journal of Weed Science, 38, 452-458.
- 27. McDonald, J.H. (2014) Biological Statistics: Data Transformation.
http://www.biostathandbook.com/transformation.html - 28. Pinel, A., N’Guessan, P., Bousalem, M. and Fargette, D. (2000) Molecular Variability of Geographically Distinct Isolates of Rice Yellow Mottle Virus in Africa. Archives Virology, 145, 1621-1638.
https://doi.org/10.1007/s007050070080 - 29. Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position Specific Gap Penalties and Weight Matrix Choice. Journal of Nucleic Acids Research, 22, 4673-4680.
https://doi.org/10.1093/nar/22.22.4673 - 30. Gouy, M., Guindon, S. and Gascuel, O. (2010) SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Molecular Biology and Evolution, 27, 221-224.
https://doi.org/10.1093/molbev/msp259 - 31. Guindon, S., Dufayard, J.F., Lefort, V., Anisimova, M., Hordijk, W. and Gascuel, O. (2010) New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Systematic Biology, 59, 307-321.
https://doi.org/10.1093/sysbio/syq010 - 32. Pinel, A. and Fargette, D. (2006) First Report of Rice Yellow Mottle Virus in Rice in Uganda. Plant Disease Journal, 90, 683.2-683.2.
- 33. Ochola, D., Issaka, S., Rakotomalala, M., Pinel-Galzi, A., Ndikumana, I., Hubert, J., Hébrard, E., Séré, Y., Tusiime G. and Fargette, D. (2015) Emergence of Rice Yellow Mottle Virus in Eastern Uganda: Recent and Singular Interplay Between Strains in East Africa and in Madagascar. Virus Research, 195, 64-72.
- 34. Fargette, D., Pinel, A., Abubakar, Z., Traoré, O., Brugidou, C., Fatogoma, S., Hébrard, E., Choisy, M., Séré, Y., Fauquet, C. and Konaté, G. (2004) Inferring the Evolutionary History of Rice Yellow Mottle Virus from Genomic, Phylogenetic, and Phylogeographic Studies. Journal of Virology, 78, 3252-3261.
https://doi.org/10.1128/JVI.78.7.3252-3261.2004 - 35. Rakotomalala, M., Pinel-Galzi, A., Mpunami, A., Randrianasolo, A., Ramavovololona, P., Rabenantoandro, Y. and Fargette, D. (2013) Rice Yellow Mottle Virus in Madagascar and in the Zanzibar Archipelago; Island Systems and Evolutionary Time Scale to Study Virus Emergence. Virus Research, 171, 71-79.
- 36. Abo, M.E., Alegbejo, M.D., Sy, A.A. and Misari, S.M. (2000) An Overview of the Mode of Transmission, Host Plants and Methods of Detection of Rice Yellow Mottle Virus. Journal of Sustainable Agriculture, 17, 19-36.
https://doi.org/10.1300/J064v17n01_04 - 37. Traoré, O., Pinel-Galzi, A., Sorho, F., Sarra, S., Rakotomalala, M., Sangu, E., Kanyeka, Z., Séré, Y., Konaté, G., Fargette, D. (2009) A Reassessment of the Epidemiology of Rice Yellow Mottle Virus Following Recent Advances in Field and Molecular Studies. Journal of Virus Research, 141, 258-267.
- 38. Fargette, D., Konate, G., Fauquet, C., Muller, E., Peterschmitt, M., Thresh, J.M. (2006) Molecular Ecology and Emergence of Tropical Plant Viruses. Annual Review of Phytopathology, 44, 235-260.
https://doi.org/10.1146/annurev.phyto.44.120705.104644 - 39. Traoré, M.D., Traoré, V.S.E., Galzi-Pinel, A., Fargette, D., Konate, G., Traore, A.S. and Traoré, O. (2008) Abiotic Transmission of Rice Yellow Mottle Virus through Soil and Contact between Plants. Pakistan Journal of Biological Sciences, 11, 900-904.
https://doi.org/10.3923/pjbs.2008.900.904 - 40. Harrington, R.B. (2002) The Heat Is on Barley Yellow Dwarf Disease: Recent Advances and Future Strategies, CIMMYT, México, D.F., 34-39.
- 41. Traoré, O., Pinel, A., Fargette, D. and Konate, G. (2001) First Report and Characterization of Rice Yellow Mottle Virus in Central Africa. Plant Disease Journal, 85, 920.1.
- 42. Sarra, S., Oevering, P., Guindo, S. and Peters, D. (2004) Wind-Mediated Spread of Rice Yellow Mottle Virus (RYMV) in Irrigated Rice Crops. Plant Pathology, 53, 148-153.
https://doi.org/10.1111/j.0032-0862.2004.00981.x - 43. Uke, A., Tibanyendela, N., Ikeda, R., Fujiie, A. and Natsuaki, K.T. (2014) Modes of Transmission and Stability of Rice Yellow Mottle Virus. Journal of Plant Protection Research, 54, 363-366.
https://doi.org/10.2478/jppr-2014-0054 - 44. Trovao, N.S., Baele, G., Vrancken, B., Bielejec, F., Suchard, M.A., Fargette, D. and Lemey, P. (2015) Host Ecology Determines the Dispersal Patterns of a Plant Virus. Virus Evolution Journal, 1, vev016.
https://doi.org/10.1093/ve/vev016