Advances in Microbiology
Vol.3 No.6A(2013), Article ID:37949,8 pages DOI:10.4236/aim.2013.36A006
Cyanobacterial Composition of Microbial Mats, Found in Brejo do Espinho and in Artificial Saline of Araruama, RJ, Brazil
1Geology Department, Federal University of Rio de Janeiro, Rio de Janeiro, Brazil
2Botanical Department, Federal University of the State of Rio de Janeiro, Rio de Janeiro, Brazil
Email: fabianefeder@yahoo.com.br, deise.odelfino@gmail.com, dolores@geologia.ufrj.br, loreineh@unirio.br
Copyright © 2013 Fabiane Feder et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received July 25, 2013; revised August 25, 2013; accepted September 3, 2013
Keywords: Cyanobacteria; Microbial Mat; Brejo do Espinho; Araruama Saline
ABSTRACT
Microbial mats are organo-sedimentary ecosystems formed by autotrophic and heterotrophic bacterias. Cyanobacteria contribute to the formation of these structures from the entrapment and engulfment of sediments contributing to the formation of carbonate rocks found in the geological record from the Precambrian to the Recent. In this research we identified the living cyanobacteria mats type 1 (gelatinous) and type 2 (cohesives) present in Brejo do Espinho and Araruama saline, east Fluminense, Rio de Janeiro state. The microbial mats type 1, were found in compartiment A and B of Brejo do Espinho and the Araruama saline in the tank with a depth of 10 cm. The mats type 2 were only found in compartiment A and in tank with 15 cm depth. In the microbial mats of compartiments A and B predominated the spherical forms of genres Chroococcus Nägeli 1948 and Aphanothece Nägeli 1948. In the tanks predominated filamentous forms of the Leptolyngbya Anagnostidis and Komárek 1988 genre. In mats type 1, the diversity indices (H') were 1.95 and 2.24 and in type 2 of 2.16 and 2.14 for the Brejo do Espinho and Araruama saline, respectively. The highest values of diversity indices found in saline tanks occurred by stable salinity conditions.
1. Introduction
The microbial mats are organo-sediment [1] formed by a microbial consortium of autotrophic and heterotrophic bacterias that acquire carbon and energy for the formation of biomass [2].
Cyanobacteria are common in microbial mats that develop in marine and hypersaline ecosystems. In general, they are distributed in the photic zone of these structures and remain abundant in the upper strata [3].
In microbial mats cyanobacteria are major primary producers [4,5] and contribute to the formation and development of these structures from the laminar growth by trapping and aggregation of sediments [6].
In Brazil, the presence of microbial mats in the east Fluminense of the state of Rio de Janeiro, in the Araruama Lagoon Complex, allows the research of the relationship with microbial sedimentation and the formation of carbonate rocks present in the geological record.
Despite the importance of diversity to ecological studies, few surveys have been developed with cyanobacteria [7]. The ecological treatment of this study was a pioneer for the microbial mats of Brejo do Espinho and Araruama saline.
The microbial mats found in Brejo do Espinho and Araruama saline, state of Rio de Janeiro, Brazil, were selected in order to identify the composition and diversity of cyanobacteria in different salinity gradients.
2. Field Site
The Araruama Lagoon Complex consists of a dual system of sand ridges, which serve as a shield against the current ocean beach.
Small and narrow merged lagoons occur in this complex between sea and the Araruama lagoon [8].
This region is characterized by a semiarid microclimate in response to the flashing of a coastal upwelling at Cabo Frio, with annual rainfall around 830 mm and annual average evaporation around 1400 mm [9].
This microclimate results in the formation of hypersaline lagoons and deployment of artificial salts on salt production in the area.
2.1. Brejo do Espinho
In Brejo do Espinho (22˚56'S and 42˚14'W) sampling occurred from September 2007 to October 2008, being selected in eight sites aboard two compartiments marked as A and B (Figure 1) [10]. This low anthropic impact area is characterized by having a length of 1 km2 and the water column that varies from 2 cm to 1.5 m [11-14].
The ongoing action from the frequent winds blowing in the region associated with low depth makes their water show no stratification. The large oscillations of the water surface and salinity have caused this system to be classified as a pond, lagoon [12,14], marsh [10] as well as Sabkha [11].
2.2. Araruama Saline
In the artificial saline of Araruama (22˚51' to 22˚55'S and 42˚20'W) samplings were conducted from August 2008 to July 2009, being selected evaporation tanks with 15 cm and 10 cm deep (Figure 1).
This saline is provided by the Araruama Lagoon, considered as a system suffocated by its close connection with the sea.
Its high degree of evaporation, the strong action of winds, intense insolation, low rainfall, the slow turnover of water and little fluvial inflow remains a hypersaline environment [15] with average salinity of 52 g∙L−1 [16].
The artificial saline environments are formed by anthropic intervention for salt exploration (NaCl), and obtaining salt based on the evaporation of brines from the sun and wind energy. Initially, the brine is stored in confined areas of the lagoon. Then, that brine travels to a tank system with continuous evaporation and concomi-
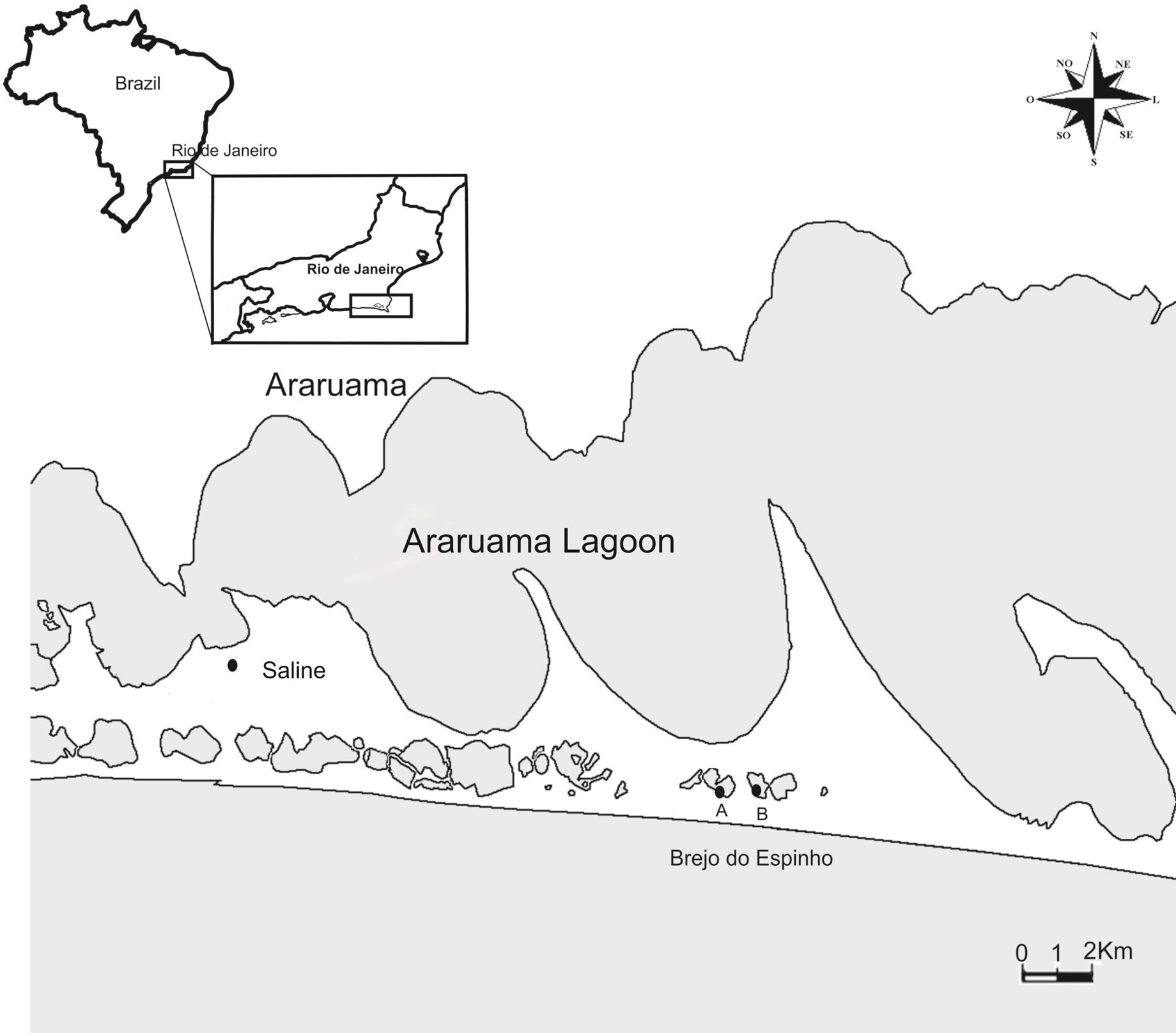
Figure 1. Map showing the location of the research area, pointing the Brejo do Espinho and Araruama saline.
tant increase of salinity to salt precipitation [17].
3. Materials and Methods
In Brejo do Espinho and Araruama saline, during the collection period were measured pH (microprocessor— pH metter GG867) and salinity (portable refractometer) with range 0 - 100‰.
The microbial mats were taken at different stages of maturity, were separated into type 1 (gelatinous) and type 2 (cohesive). The structures type 1 were found in compartiment A and B of Brejo do Espinho and in the salinity tank with 10 cm depth. While the structures type 2 were only in compartiment A and in the tank with 15 cm depth.
These microbial mats were removed from the collection site and placed in sterile plastic containers in the dark with a solution of 4% formalin buffered with borax.
These structures were measured in thickness with the aid of calipers Mitutoyo (JIF B7507). The samples were horizontally sectioned for the initial 5 mm separation of the structure, being withdrawn subsamples from a 2 mm dermatological punch for making permanent blades. Reading of these was performed by using an optical microscope Olympus CX40, increases in the 40 to 100 times, sweeping three random columns per blade.
The identification of cyanobacterial composition followed [18,19].
The methodology for ecological quantification was modified and adapted by [20]. A total number of 1500 individuals was considered to evaluate the representativeness of species by microbial mat. The ecological interpretation occurred from the diversity index proposed by Shannon Weaver (H'), equitability and richness. The Software program used was the Paleontological Statistics (PAST) version 2.17 [21].
4. Results
The pH measured in the field in compartiments A and B of the Brejo do Espinho and tanks of Araruama saline always remained basic. In compartiment B and in salinity tanks obtained the minimum value of 7.0 and a maximum of 10 (Table 1).
Salinity in the compartiments of Brejo do Espinho showed great variation, ranging from a minimum value of 16‰ and above 100‰. In the Araruama saline, the salinity maintained values greater than 100‰ (Table 1).
The microbial mats analyzed for this research were disposed in Brejo do Espinho and Araruama saline as flat, smooth, continuous and uniform structures.
In Brejo do Espinho the structures of type 1 were observed in compartment A and B with 16 mm thick, being found in the dry period and in the initial lagoon filled period in the submerged portion of the lagoon. These
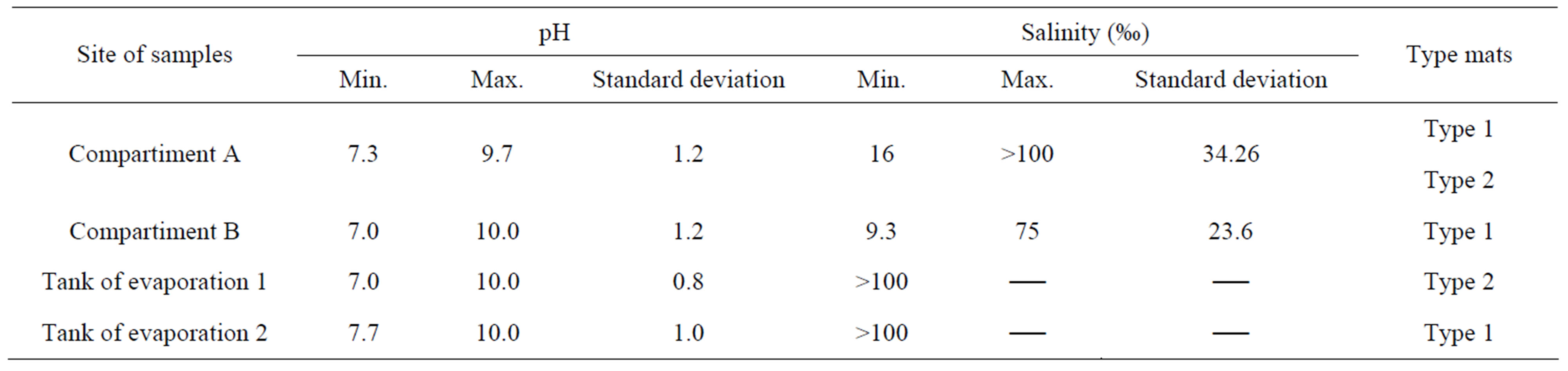
Table 1. Physico-chemical parameters of pH and salinity and the types of microbial mats found in Brejo do Espinho (compartiment A and B) and Araruama saline (evaporation tanks of 10 and 15 cm depth).
mats were observed multicolored strata with yellow, green, red and brown colors.
In saline, these structures showed little cohesive, thin and brittle with a thickness of 5 mm, green color and evaporate minerals on their surface. They were found throughout the collection period in evaporation tanks of 10 cm.
The structures of type 2 of Brejo do Espinho proved to be cohesive with 20 mm thick, made of multicolored layers, with yellowish, green, red and brown that are repeated throughout the structure. They were found only in the dry lagoon period.
In Araruama saline this type of structure was found in the initial evaporation tanks with water layer of 15 cm, it had a thickness of 15 mm. These structures were found throughout the collection period, with upper stratum green, intermediate red and brown bottom.
In mat type 1 the composition cyanobacterial presented a richness of 30 species in Brejo do Espinho and 17 species in Araruama saline. In Brejo do Espinho the genres more representatives in number of species were Chroococcus Nägeli 1849 with 24% Aphanothece Nägeli 1849 and Leptolyngbya Anagnostidis and Komárek 1988 with 11%. In Araruama saline the genres Aphanothece Nägeli 1849 and Chroococcus Nägeli 1849 were the ones who gathered the largest number of species, both with 29% (Figure 2).
In mats type 2 showed a richness of 27 species in Brejo do Espinho and 16 in Araruama saline. In both structures the genres most representative were Aphanothece Nägeli 1849 with 29% and 32% and Chroococcus Nägeli 1849 14% and 20% (Figure 2), respectively.
In relation to gelatinous mat type 1 were common to Brejo do Espinho and the saline species Aphanothece clathrata West and West 1906, Chroococcus minor (Kützing) Nägeli 1849, Chroococcus turgidus (Kützing) Nägeli 1849 (Table 2).
In mat type 2 were common Aphanothece castagnei (Brébisson) Rabenhorst 1865, Aphanothece microscopica Nägeli 1849, Aphanothece stagnina (Sprengel) A. Braun 1863, Microcoleus cthonoplastes (Thuret) Gomont 1892 and Schizothrix friesii (Agardh) Gomont 1892 (Table 2).
In gelatinous mat of Brejo do Espinho was possible to see from the values of the relative abundance the predominance of species Chroococcus sp. (0.42) Leptolyngbya sp. (0.24) (Figure 2).
In the saline the same structure type showed from the relative abundance of predominant species Leptolyngbya hypolimnetica (0.24), Leptolyngbya tenuis (0.19) and Chroococcus minimus (0.13) (Figure 3).
In Brejo do Espinho and saline, gelatinous mats showed diversity index H'1.95 and 2.24 and equitability of 0.57 and 0.79 (Table 3), respectively.
In structure type 2 of Brejo do Espinho, from the values of relative abundance the species that show dominant, were species Aphanothece stagnina (Sprengel) A. Braun 1863 (0.43), Aphanothece clathata (0.16) (Figure 3).
In mat type 2 from saline occurred predominantly three species based on relative abundance, the Leptolyngbya hypolimnetica (0.31), Chroococcus minutus (0.20) and Chroococcus minimus (0.11) (Figure 3).
Type 2 structures found in Brejo do Espinho and saline showed diversity index (H') of 2.16 and 2.14 and equitability of 0.65 and 0.79 (Table 3), respectively.
5. Discussion
Brejo do Espinho is a dynamic system governed by seasonal cycles of drought and flood lagoons, which are determined by regional climatic conditions [10] and cause large fluctuations in salinity as observed during the research period.
The mats type 1 found in Brejo do Espinho during drought and flood lagoon proved well adapted to conditions of low and high salinity. While that type 2 structures, present only in the dry lagoon, highlighted the need for a stable conditions and high salinity for its formation.
Araruama saline despite of having tanks with low depth had found some stability in salinity always above 100‰.
According to [22] the location of saline in dry climates with low rainfall justifies the high salinities found in these salines and the continuous exploitation of salt.
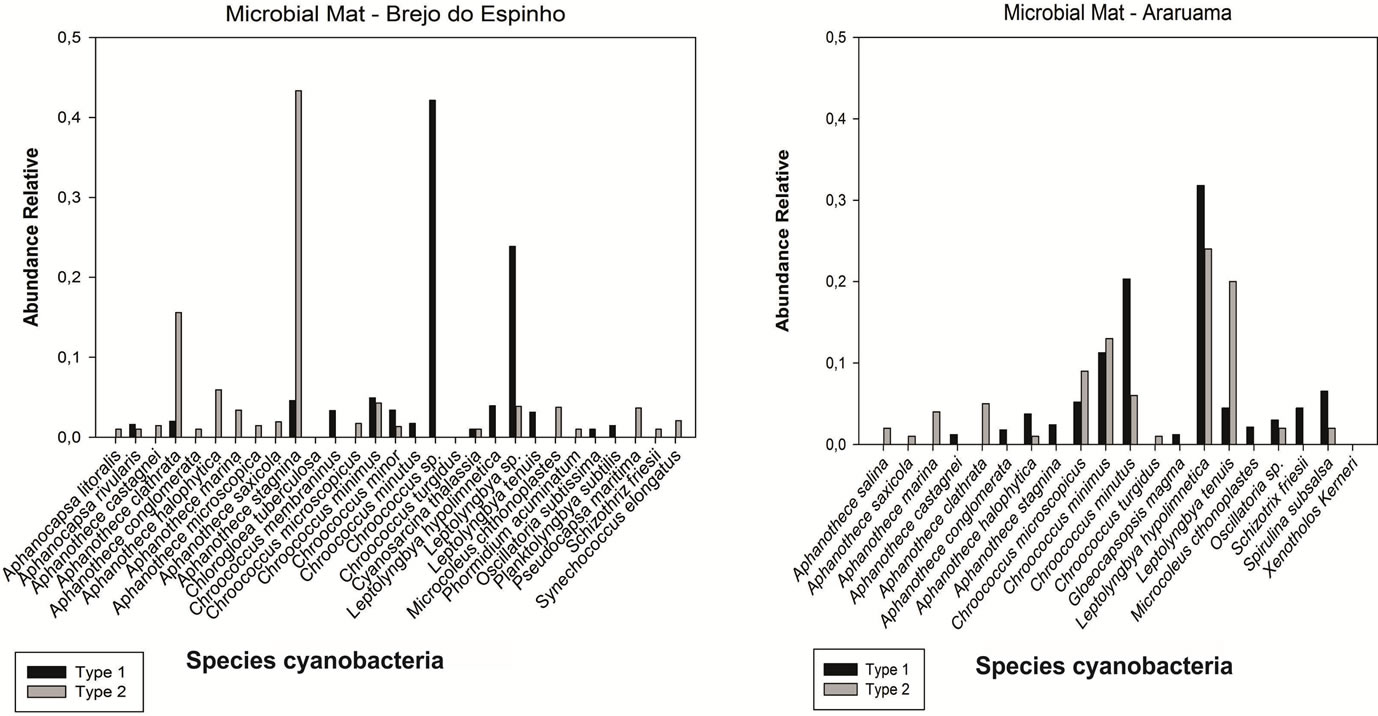
Figure 2. Graphs of the qualitative distribution of genres in mats type 1 and type 2 of Brejo do Espinho and Araruama saline.
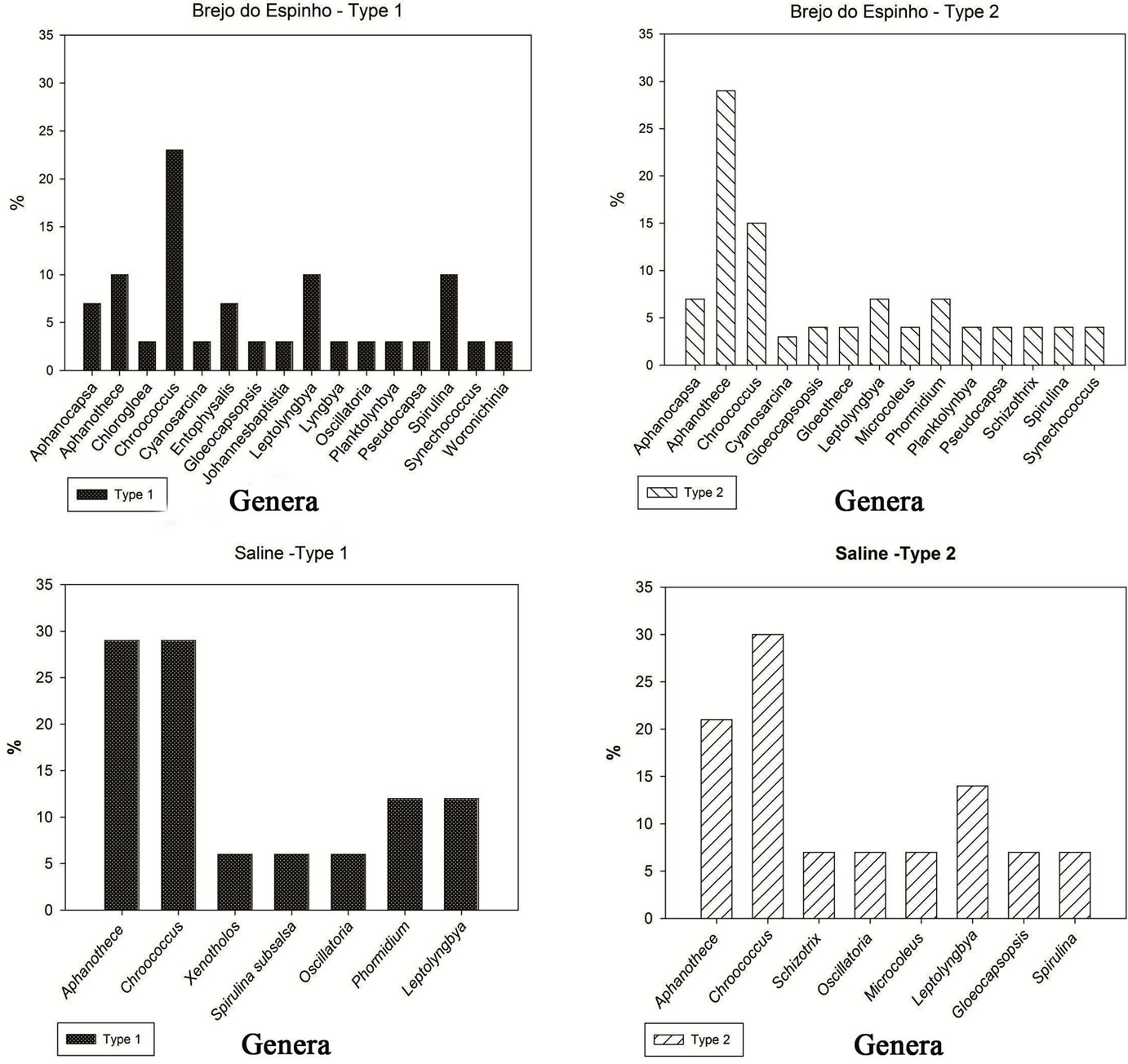
Figure 3. Graphs representing the relative abundance of species in mats type 1 and type 2 of Brejo do Espinho and Araruama saline.
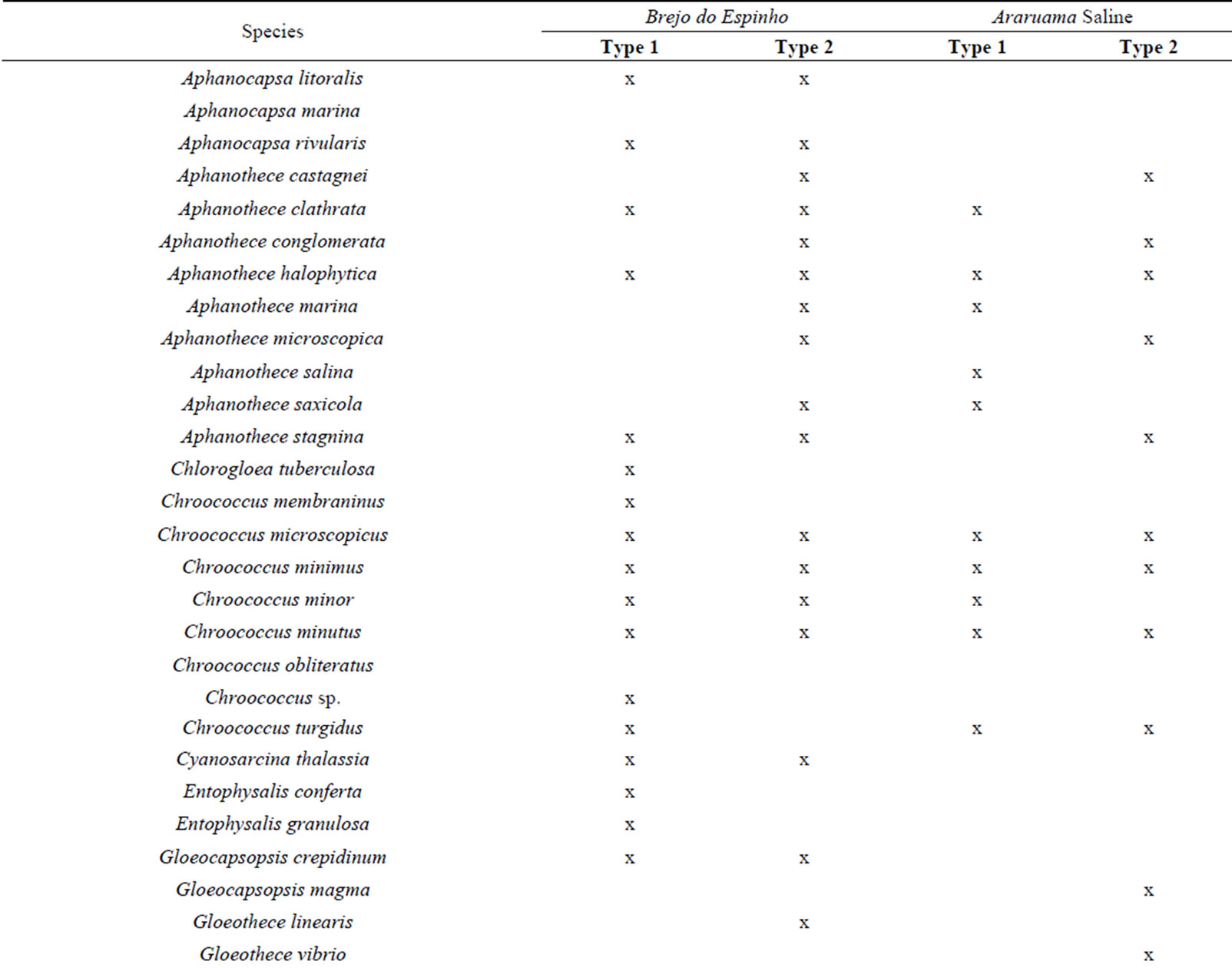
Table 2. Composition of cyanobacterial plain microbial mats type 1 and type 2 found in Brejo do Espinho and Araruama saline.
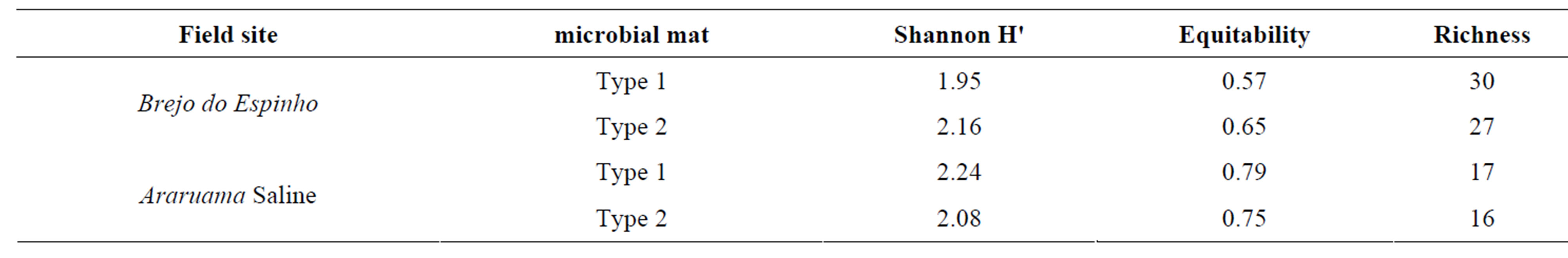
Table 3. Diversity index, equitability and richness to the mats type 1 and type 2 of Brejo do Espinho and Araruama saline.
Locations where samples were collected are considered as intermediate salinity tanks (above 100‰ and below 150‰), having structures in development, as noted by [21] in saline for most authors, by [23,24]. In saline of Cabo Frio, Rio de Janeiro.
Microbial mats type 1, probably are found in evaporation reservoirs with higher salinity for presenting on its surface evaporate precipitation.
Microbial mats analyzed showed distinct cyanobacterial composition in different salinity regimes which explains the low number of species found in common in Brejo do Espinho and Araruama saline corroborating [25] in microbial mats of Arabian Gulf.
Individual filamentous Microcoleus chthonoplastes common in microbial mat type 2 are considered as prominent phototrophic organisms composing these structures. This species was not considered abundant as observed in saline of Guerrero Negro, Mexico [26], the saline of Girauld in southern France [27] and Puerto Rico [28].
The apparent abundance of this species is cited by several authors for their biovolume occupying an area greater microbial mat when compared with other species of cyanobacteria [29].
The spherical forms of cyanobacteria were the major constituents in microbial mats of the Brejo do Espinho, being also abundant in the structures of Araruama saline.
The predominance of spherical forms were also observed by [30] from microbial mats Salar Llámara (Atacama Desert, Chile) and [31] for microbial mats of Saline Cabo de Gata that corroborate by mentioning spherical species that are adapted to hypersaline environments.
The greater abundance of Aphanothece stagnina in microbial mats of Brejo do Espinho in mats type 2 indicates that this species is hardy and well adapted to conditions of high salinity [32]. In tanks, this species showed no significant relative abundance, being found only in the microbial mat type 2.
[23] considered this species as resistant to high salinity, being found in Salines of Cabo Frio, Rio de Janeiro, Brazil.
The genre Aphanothece is reported as a major photosynthetic groups in hypersaline mats of Guerrero Negro (Mexico), Solar Lake (Sinai) and Salines of Giraud (France) by [33]. This genre was also present in microbial mats of brackish environments in the Caribbean according [34].
The high values of relative abundance of Chroococcus sp. in mats type 1 of Brejo do Espinho is corroborated by the presence of the genre Chroococcus in microbial mats of several lagoons in Belize (Caribbean) [34], in Camargue (Iberia) [35] and Ebro Delta (Iberia) [36].
The high relative abundance of filamentous cyanobacteria of the genre Leptolyngbya found on the surface of microbial mats in Araruama saline contributes to the growth of the structure, as well as colony formation by spherical shapes genre Aphanothece and Chroococcus found abundantly in mats of Brejo do Espinho, that showed thicker than those found in saline. In distinct environments, different cyanobacteria can occupy similar ecological niches contributing to the formation of microbial mats [37].
Spherical and filamentous species of cyanobacteria promote the formation of upper layers from the trapping of sediment particles that are deposited on the structure.
The spherical species that dominate the surface are organized in colonies, producing in conjunction with the filamentous species relevant quantities of EPS which assist in capturing and trapping of sediment according to [38].
In addition, [39] noted that the arrangement of the spherical cells in colonies serves to provide protection in environments of intense solar radiation.
The predominant species in the spherical surface of microbial mats of the Brejo do Espinho possibly resulted in yellowing of its upper stratum evidenced by high carotenoid production for protection of hydrous and luminous stress [30,33,40].
[41] mentions that microbial mats show high microbial diversity, agreeing with the high values of diversity and equitability found in microbial mats type 1 and 2 of Brejo do Espinho and Araruama saline.
Biodiversity depends on the salinity gradient, having a reduction of microbial diversity with its increase. However, in microbial mats of hypersaline environments the diversity for phototrophic oxygenic organisms is high with the occurrence of Microcoleus chthonoplastes and the genera Phormidium according to [27].
In this research, mats type 2 is an example of diversity and the presence of these groups.
Cyanobacterial diversity and equitability were higher in saline than in the Brejo do Espinho, which according to [25] may be related to environmental conditions more stable in saline.
In Brejo do Espinho was found in mats type 1 the lowest values of diversity and equitability. These structures were observed under conditions of high salinity variation with predominance of species Chroococcus sp. and Leptolyngbya sp., which are probably the most favored and adapted to these environmental conditions.
Brejo do Espinho got diversity values similar to [26] in natural saline of Guerrero Negro, Mexico.
The ecological study of the composition of cyanobacteria based on classical taxonomy presented limitations. Groups of prokaryotes with similar morphologies may represent distinct phylogenetic entities [26]. However, cyanobacteria are a special case, as its botanical name is based on morphological preceding bacterial identification. In fact, the diversity and richness of cyanobacterial populations are estimated by morphology in ecological studies of phytoplankton [28].
6. Conclusions
In microbial mats type 1 and 2 of the Brejo do Espinho and Araruama saline, few species of cyanobacteria were common indicating relationship between the distribution of species and different salinity gradients.
The floatability environmental of Brejo do Espinho favored spherical forms with a predominance of genres Chroococcus Nageli 1948 and Aphanothece 1948.
The stability in salinity of Araruama saline set higher values of diversity and dominance of filamentous species of the genre Leptolyngbya Anagnostidis and Komárek 1988 in the formation of these structures.
REFERENCES
- H. Van Germanden, “Microbial Mats: A Joint Venture,” Marine Geology, Vol. 113, No. 1-2, 2003, pp. 3-25. http://dx.doi.org/10.1016/0025-3227(93)90146-M
- L. K. Baumgartner, R. P. Reid, C. Dupraz, A. W. Decho, D. H. Buckley, J. R. Spear, K. M. Przekop and P. T. Visscher, “Sulphate Reducing Bacteria in Microbial Mat: chaging Paradigms, New Discoveries,” Sediment Geology, Vol. 185, No. 3-4, 2006, pp. 131-145. http://dx.doi.org/10.1016/j.sedgeo.2005.12.008
- D. J. Des Marais, “Biogeochemistry of Hypersaline Microbial Mats Illustrates the Dynamics of Modern Microbial Ecosystems and the Early Evolution of the Biosphere, Biology,” Biology Bulletin, Vol. 204, No. 2, 2003, pp. 160-163.
- A. W. Decho, “Microbial Exopolymer Secretion in Ocean Environments: Their Role(s) in Food Webs and Marine Processes,” Oceanography and Marine Biology—Annual Review, Vol. 28, 1990, pp. 73-154.
- M. I. Kühl, T. Fenchel and J. Kasmierczak, “Growth, Structure and Calcification Potential of an Artificial Cyanobacterial Mat,” In: W. E. Krumbein, D. M. Paterson and G. Zavarzin, Eds., Fossil and Recent Biofilms the Natural History of the Impact of Life on Planet Earth, Kluwer Academic Publisher, Dordrecht, 2003, pp. 77- 102.
- N. Noffke, G. Gerdes and T. Klenke, “Benthic Cyanobacteria and Their Influence on the Sedimentary Dynamics of Peritidal Depositional Systems (Siliciclastic, Evaporitic Salty, and Evaporitic Carbonatic),” Earth-Science Reviews, Vol. 62, No. 1-2, 2003, pp. 163-176. http://dx.doi.org/10.1016/S0012-8252(02)00158-7
- F. Garcia-Pichel, U. Nubel, G Muyzer and M. Kuhl, “On Cyanobacterial Community Diversity and Its Quantifications” Cyanobacterial Diversity & Adaptation, Vol. 65, 1999, pp. 422-430.
- P. B. S. Primo and C. R. S. F. Bizerril, “Lagoa de Araruama. Perfil Ambiental do Maior Ecossistema Lagunar Hipersalino do Mundo,” SEMADS, Rio de Janeiro, 2002, pp. 33-35.
- E. Barbiére and R. Coe Neto, “Spatial and Temporal Variation of Rainfall of the East Fluminense Coast and Atlantic Serra do Mar, State of Rio de Janeiro, Brazil,” In: B. Knoppers, E. D. Bidone and J. J. Abrão, Eds, Environmental Geochemistry of Coastal Lagoon Systems of Rio de Janeiro, Brazil, FINEP (Série Geoquímica Ambiental 6), Rio de Janeiro, 1999, pp. 25-46.
- D. O. Delfino, M. D. Wanderley, L. H. Silva e Silva, F. Feder, F. A. S. Lopes, “Sedimentology and Temporal Distribution of Microbial Mats from Brejo do Espinho, Rio de Janeiro, Brazil,” Sedimentary Geology, Vol. 263- 264, 2012, pp. 85-95. http://dx.doi.org/10.1016/j.sedgeo.2011.08.009
- C. F. Barbosa, “Reconstituição Paleoambiental de Fácies Lagunares com Base em Foraminíferos: O Nível do mar no Quaternário Superior na Área de Cabo Frio, RJ,” Ph.D. thesis, University of São Paulo, São Paulo, 1997.
- A. P. A. Anjos, “Caracterização da Sedimentação Atual de Lagoas Costeiras do Litoral do Estado do Rio de Janeiro para Fins de Calibração de Acordo com as Variações Paleoambientais e Paleoclimáticas,” Ph.D. Dissertation, Federal University Fluminens, Niterói, 1999.
- Y. Van Lith, C. Vasconcelos, R. Warthmann, J. C. F. Martines and J. A. Mckenzie, “Bacterial Sulfate Reduction and Salinity: Two Controls on Dolomite Precipitation in Lagoa Vermelha and Brejo do Espinho (Brazil),” Hydrobiologia, Vol. 485, No. 1-3, 2002, pp. 35-49. http://dx.doi.org/10.1023/A:1021323425591
- A. P. A. Anjos, “Processo de Precipitação de Dolomita na Lagoa Brejo do Espinho: Uma Contribuição para Reconstrução Ambiental,” Ph.D. Thesis, Federal University Fluminens, Niterói, 2004.
- B. Kjerfve, “Coastal Lagoon Processes,” Elsevier, Amsterdan, 1994.
- P. F. M. Turcq, “Impact Low Salinity Year on the Metabolism of a Hipersalina Coastal Lagoon (Brazil),” Hidrobiologia, Vol. 429, No. 1-3, 2000, pp. 133-140. http://dx.doi.org/10.1023/A:1004037624787
- R. Wit and J. O. Grimalt, 1992 “Microbial Ecosystems in Spanish Coastal Salinas; An Ecological and Geochemical Research of Biomarkers,” Limnetica, Vol. 8, 1992, pp. 205-212.
- K. Anagnostidis and J. Komárek, “Modern Approach to the Classification System of Cyanophytes. Archiv für Hidrobiologie,” Algological Studies, Vol. 80, No. 1-4, 1988, pp. 327-472.
- J. Komárek and K. Anagnostidis, “Süβwasserflora von Mitteleuropa Band 19/1: Cyanophyta 1. Teil: Chroococcales,” 2nd Edition, Gustav Fischer, Stuttgart, 1999.
- U. Nübel, F. Garcia-Pichel, M. Kühl and G. Myzer, “Quantifying Microbial Diversity: Morphotypes, 16 rRNA Genes, and Carotenoids of Oxygenic Phototrophs in Microbial Mats,” Applied and Environmental Microbiology, Vol. 65, No. 2, 1999, pp. 422-430.
- Ø. Hammer, D. A. T. Harper and P. D. Ryan, “Past: Paleontological Statistics Software Package for Education and Data Analysis,” Palaeontologia Electronica, Vol. 4, No. 1, 2001, p. 9.
- J. S. Davis, “Structure, Function, and Management of the Biological System for Seasonal Solar Saltwork,” Global Nest: The International Journal, Vol. 2, No. 3, 2001, pp. 217-226.
- M. H. C. Baeta Neves and A. J. Casarin, “As Cianofíceas das Salinas de Cabo Frio—Brasil,” Acta Biológia Leopoldinense, Vol. 12, No. 1, 1990, pp. 99-123.
- M. A. M. Silva and C. L. Santos, “Halitas das Salinas de Cabo Frio: Reconhecimento das Morfologias como Subsídios para o Entendimento das Halitas Pretéritas,” Boletim de Geociência da Petrobrás, Vol. 11, No. 1-2, 1997, pp. 74-83.
- R. M. M. Abed, K. Kohls and D. de Beer, “Effect of Salinity Changes on the Bacterial Diversity, Photosynthesis and Oxygen Consumption of Cyanobacterial Mats from an Intertidal Flat of the Arabian Gulf,” Environmental Microbiology, Vol. 9, No. 6, 2007, pp. 1384-1392. http://dx.doi.org/10.1111/j.1462-2920.2007.01254.x
- U. Nübel, M. M. Bateson, M. K. Madigan and M. D. Ward, “Diversity and Distribution in Hypersaline Microbial Mat of Bacteria Related to Chloroflexus spp.,” Applied and Enviroment Microbiology, Vol. 67, No. 9, 2001, pp. 4361-4365. http://dx.doi.org/10.1128/AEM.67.9.4365-4371.2001
- A. Fourçans, T. G. Oteyza, A. Wieland and A. Solé, “Characterization of Functional Bacterial Groups in a Hypersaline Microbial Mat Community (Saline-de-Giraud, Camargue, France),” FEMS Microbiology Ecology, Vol. 51, No. 1, 2004, pp. 55-70. http://dx.doi.org/10.1016/j.femsec.2004.07.012
- L. Casillas-Martínez, M. L. Gonzalez, Z. Fuentes-Figuerosa, C. M. Castro, W. Ramirez, R. E. Systema, J. PerezJimenez and P. T. Visscher, “Community Structure, Geochemical Characteristics and Mineralogy of a Hypersaline Microbial Mat, Cabo Rojo, PR,” Geomicrobiology Journal, Vol. 22, No. 6, 2005, pp. 269-281. http://dx.doi.org/10.1080/01490450500182672
- C. L. Ortiz, “Species Richness of Cyanobacterial, Diatoms, and Ciliates in Microbial Mats of the Cabo Rojo Salterns, Puerto Rico,” Ph.D. Dissertation, Universidade de Porto Rico, 2008.
- C. Dermegasso, G. Chong, P. Galleguillos, L. Escudero, M. Martínez-Alonso and I. Esteves, “Tapestes Microbianos del Salar de Ilamará, Norte del Chile,” Revista Chilena de Historia Natural, Vol. 76, No. 3, 2003, pp. 485- 499.
- I. Esteve, M. Martínez-Alonso, J. Mir and R. Guerrero, “Distribuition, Typology and Structure of Microbial Mat Communities in Spain: A Preliminary Research,” Limnetica, Vol. 8, 1992, pp. 185-195.
- G. Hällfors, “Checklist of Baltic Sea Phytoplankton Species (Including Some Heterotrophic Protistan Groups),” Helsinki Commission, Baltic Marine Environment Protection Commission, Filand, 2004.
- M. Martínez-Alonso, J. Mir, P. Caumette, N. Gaju, R. Guerrero and I. Esteve, “Distribution of Phototrophic Populations and Primary Prodution in a Microbial Mat from the Ebro Delta, Spain,” International Microbiology, Vol. 7, 2004, pp. 19-25.
- E. Rejmánková and J. Komárková, “A Function of Cyanobacterial Mats in Phosphorus-Limited Tropical Wetlands,” Hydrobiology, Vol. 431, No. 2-3, 2000, pp. 135- 153. http://dx.doi.org/10.1023/A:1004011318643
- P. Caumette, R. Matheron, N. Raymond and J. C. Relexans, “Microbial Mats in the Hypersaline Ponds of Mediterranean Salterns (Salins-de-Giraud, France),” FEMS Microbiology Ecology, Vol. 13, No. 4, 1994, pp. 273-286. http://dx.doi.org/10.1111/j.1574-6941.1994.tb00074.x
- J. Urmeneta and A. Navarrete, “Mineralogical Composition and Biomass Studies of the Microbial Mats Sediments from the Ebro Delta, Spain,” International Microbiology, Vol. 3, No. 2, 2000, pp. 97-101.
- U. Nubel, F. Garcia-Pichel, M. Kuhl and G. Muyzer, “Quantifying Microbial Diversity: Morphotypes, 16S rRNA Genes, and Carotenoids of Oxygenic Phototrophs in Microbial Mats,” Applied and Enviromental Microbiology, Vol. 65, No. 2, 1999, pp. 422-430.
- L. J. Stal, “Microphytobenthons as Biogeomorphological Force in Intertidal Sediment Stabilization,” Ecological Engineering, 2009.
- F. Garcia-Pichel and R. W. Castenholz, “Comparative Anoxygenic Photosynthetic Capacity in 7 Strain of a Thermophilic Cyanobacterium,” Archives of Microbiology, Vol. 153, No. 4, 1990, pp. 344-351. http://dx.doi.org/10.1007/BF00249003
- A. Oren, “Saltern Evaporation Ponds as Model Systems for the Research of Primary Production Processes under Hypersaline Conditions,” Aquatic Microbial Ecology, Vol. 56, No. 2-3, 2009, pp. 193-204. http://dx.doi.org/10.3354/ame01297
- R. E. Ley, J. K. Harris, J. Wilcox, J. R. Spear, S. R. Miller, B. M. Bebout, J. A. Maresca, D. A. Bryant, M. L. Sogin and N. R. Pace, “Unexpected Diversity and Complexity of the Guerrero Negro Hypersaline Microbial Mat,” Applied and Environmental Microbiology, Vol. 72, No. 5, 2006, pp. 3685-3695. http://dx.doi.org/10.1128/AEM.72.5.3685-3695.2006