Advances in Microbiology
Vol.3 No.1(2013), Article ID:29122,4 pages DOI:10.4236/aim.2013.31006
Activation of Ethanol Production by Combination of Recombinant Ralstonia eutropha and Electrochemical Reducing Power
1Department of Biological Engineering, Seokyeong University, Seoul, South Korea
2Department of Radiation Biology, Environmental Radiation Research Group, Korea Atomic Energy Research Institute, Daejeon, South Korea
Email: *baakdoo@skuniv.ac.kr
Received November 21, 2012; revised December 23, 2012; accepted January 25, 2013
Keywords: Ralstonia eutropha; Electrochemical Reducing Power; Genetic Recombination; Pyruvate Decarboxylase; Alcohol Dehydrogenase
ABSTRACT
Ralstonia eutropha was genetically modified to induce ethanol production from glucose. An electrochemical bioreactor was prepared to generate electrochemical reducing power coupled to regeneration of NADH. Growing cells of recombinant R. eutropha produced about 29 mM of ethanol in conventional conditions and 56 mM of ethanol in electrochemically reduced conditions from 100 mM glucose. Grown cells of the recombinant produced about 52 mM of ethanol in conventional conditions and 142 mM of ethanol in electrochemically reduced condition from 100 mM glucose. These results are a clue that electrochemical reducing power can induce the recombinant R. eutropha to produce more ethanol coupled to increase of NADH/NAD+ ratio.
1. Introduction
Electrochemical reduction reaction can be separated from the electrochemical oxidation reaction by a specially designed electrochemical bioreactor [1,2]. The electrochemically reduced neutral red catalyzes regeneration of biochemical reducing power (NADH) without enzyme catalysis [3,4]. Increase of the NADH/NAD+ ratio induces an increase of metabolites that are produced coupled to NADH oxidation to NAD+ in fermentative metabolism [5,6]. The electrochemical reducing power can be obtained from electricity generated from solar energy without combustion of fossil fuels. An electrochemical bioreactor is a useful device to accomplish conversion of solar energy to biochemical energy and carbon dioxide to organic compounds [1,7].
The nutritionally versatile Ralstonia eutropha is a facultative autotroph that can assimilate CO2 in coupling with metabolic oxidation of H2 via Calvin-BensonSassham reductive pentose-phosphate cycle [8] and can also heterotrophically grow with glucose [9]. Growth of R. eutropha was known to be less than 1.0 on an optical density basis in the autotrophic condition with H2 and CO2 as a sole energy and carbon source but maximally 6 times increased in the mixotrophic condition with H2, CO2, and amino acids [10-12]. R. eutropha autotrophically grows with CO2 and electrochemical reducing power as a sole carbon and energy source, but doesn’t produce any metabolite that is alcohol and organic acid in both autotrophic and heterotrophic condition [13]. Practically, ethanol production of R. eutropha may be induced by recombination of specific genes encoding pyruvate decarboxylase and alcohol dehydrogenase genes that were obtained from Zymomonas mobilis [14,15]. Ethanol production was proportionally increased to the ratio of NADH/NAD+ in fermentative metabolism [6]. Accordingly, combination of intrinsically autotrophic metabolism, extrinsically genetic recombination, and electrochemically regenerated reducing power may be the best way to induce R. eutropha to produce ethanol from CO2. This study is performed to test the possibility that the metabolic function of the recombinant R. eutropha for ethanol production may be activated by the electrochemical reducing power under heterotrophic condition.
2. Materials and Methods
2.1. Transformation of R. eutropha
The pLOI 3018-10 plasmid containing the genes encoding pyruvate decarboxylase and alcohol dehydrogenase II were kindly offered by Ingram [14,16]. The plasmid was inserted into R. eutropha by using an electroporator (MicroPulser, BioRad Laboratories, Richmond, CA, USA) at 2.5 KV for 2.5 msec according to the user’s manual. All of procedures for preparation of electrocompetent cells of R. eutropha, culture medium, plasmid DNA, buffers, and selection medium were performed according to methods used by Taghavi et al. [17] and transformant capable of producing ethanol was selectively isolated from agar plate medium according to the method used by Conway et al. [18].
2.2. Cultivation of Transformant
An isolate was cultivated in M9 mineral medium (disodium phosphate 6.8 g/L, monosodium phosphate 3 g/L, ammonium chloride 5 g/L, sodium chloride 0.5 g/L, magnesium sulfate 0.246 g/L, calcium chloride 0.0147 g/L) supplemented with 10 mM NaHCO3, 18 g glucose/L, and 3 g yeast extract/L.
2.3. Ethanol Production of Transformant
An electrochemical bioreactor that was designed for continuous culture in previous research was partially modified for batch culture of recombinant R. eutropha, shown in Figure 1. Graphite felt (thickness, 10 mm; height, 200 mm; length, 500 mm; Electrosynthesis, USA) modified with neutral red was used as a cathode and platinum wire (0.3 mm thickness, 400 mm length) was used as a counter electrode [13]. Two thousand ml of the modified M9 medium was prepared in the electrochemical bioreactor (Figure 1) to which O2-free CO2 (50 ml·min−1) was continuously supplied during cultivation. Wild type or recombinant R. eutropha previously cultivated for 24 h was inoculated at the ratio of 5% (v/v). DC −3 V of electricity was charged to the cathode for culture test in the electrochemical reduction condition but not during culture test in the conventional condition. One hundred mM of K2HPO4 was used as anolyte.
2.4. Analysis
Ethanol production was analyzed by GLC (Acme 6000, Younglin Instrument, Seoul, Korea) equipped with a flame-ionized detector and DB-Wax column (30 m × 0.535 mm, Agilent Technology, CA, USA). Oven temperature was initially maintained at 50˚C for 5 min and then gradually increased to 150˚C at the rate of 20˚C a minute. Split ratio of sample and split flow of mobile phase (99.999% helium) was adjusted to 20:1 and 16.1 ml/min, respectively. Flow rate of mobile phase and injection volume was adjusted to 0.8 ml/min and 1 μl, respectively. 20 mM of valeric acid was used as an internal standard. Glucose consumption was analyzed by HPLC equipped with a refractive index detector (Younglin Instrument, Seoul, Korea) and HPX-87H ion exchange column (BioRad Laboratories, Richmond, CA, USA). Temperature of column and detector was adjusted to 35˚C, and flow rate of mobile phase and injection volume of sample was adjusted to 0.6 ml/min and 20 μL, respectively.
3. Results and Discussion
In the time profile of batch culture for recombinant R. eutropha aimed at the production of ethanol by the introduction of genes encoding pyruvate decarboxylase and alcohol dehydrogenase II, glucose consumption and ethanol production was evaluated as shown in Figure 2. The recombinant R. eutropha produced approximate two times of ethanol in the electrochemically reduced condition than conventional condition; however, glucose was mostly consumed and not influenced by the cultivation conditions. Generally, the ethanol production by fermentative microorganisms that are Zymomonas mobilis and Saccharomyces cerevisiae is proportional to glucose consumption, with a ratio of ethanol to glucose of 2 to 1 [19,20]. However, the ratio of ethanol production to glucose consumption by recombinant R. eutropha was about 0.28 to 1.0 in the conventional condition and 0.58 to 1.0 in the electrochemically reduced condition. The metabolic intermediates and reducing power (NADH) generated in coupling with glucose oxidation may be partially
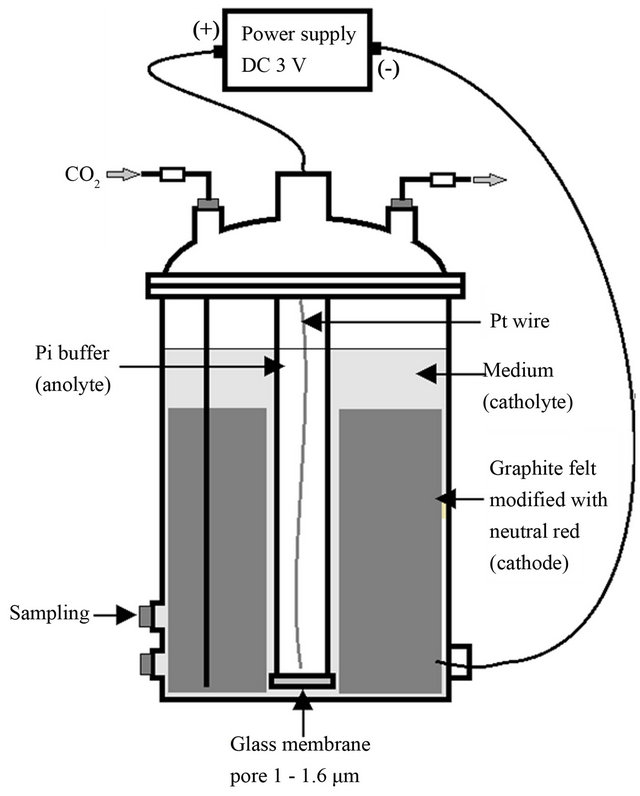
Figure 1. Electrochemical bioreactor to induce biochemical reducing power for bacterial metabolism in coupling with redox reaction of neutral red immobilized in graphite felt.
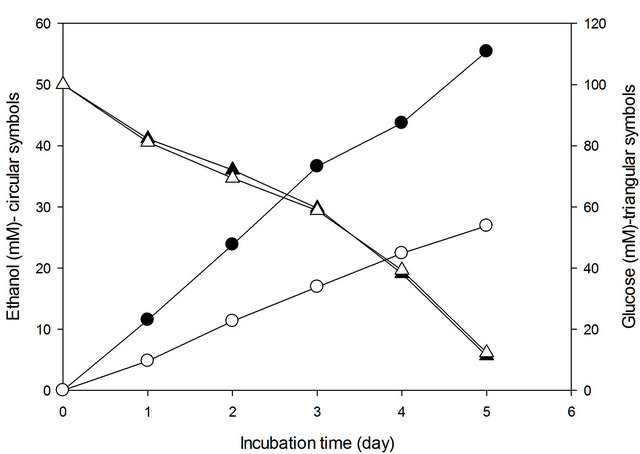
Figure 2. Ethanol production by recombinant Ralstonia eutropha that was cultivated in electrochemical reduction condition (solid symbols) and conventional condition (open symbols).
consumed for ethanol production and biosynthesis of building blocks in heterotrophically growing cells [21, 22]. The ratio of ethanol production to glucose consumption by grown cells of recombinant R. eutropha that was previously cultivated for 48 h and harvested and then suspended in 0.5 volume of fresh medium was increased to be 0.52 to 1.0 in conventional condition and 1.42 to 1.0 in electrochemically reduced condition as shown in Table 1. Theoretically, grown cells of fermentative bacteria may less consume reducing power for biosynthesis of building blocks but more consume reducing power for metabolite production than growing cells. The higher ethanol production by the grown cells compared to the growing cells may be caused by higher production of reducing power in proportion to biomass and higher consumption of reducing power coupled to ethanol production. The higher ethanol production in the electrochemically reduced condition may be caused by increased reducing power (ratio of NADH/NAD+ balance) that is electrochemically reduced coupled to redox reaction of neutral red immobilized in cathode [6,23].
Conversion of electrochemical reducing power to biochemical reducing power (high balance of NADH/NAD+) has been employed to induce increase of specific metabolite production from glucose by fermentative metabolism [4]. Ethanol produced from 1 M glucose by ethanol-fermentation bacteria or yeast has to be less than 2 M because all of NADH can’t be oxidized coupled to reduction of acetaldehyde to ethanol; however, 2 M ethanol can be produced from 1 M glucose by the coupling with the electrochemical redox reaction of neutral red. A system for ethanol production from glucose may not be required to improve on the technical basis but may be required to develop on the material basis.
R. eutropha can autotrophically grow with electrochemical reducing power and CO2 but doesn’t produce
Table 1. Growth, glucose consumption, and ethanol production of R. eutropha and recombinant R. eutropha cultivated in conventional condition (CC) and electrochemically reduced condition (ER) for 120 h.
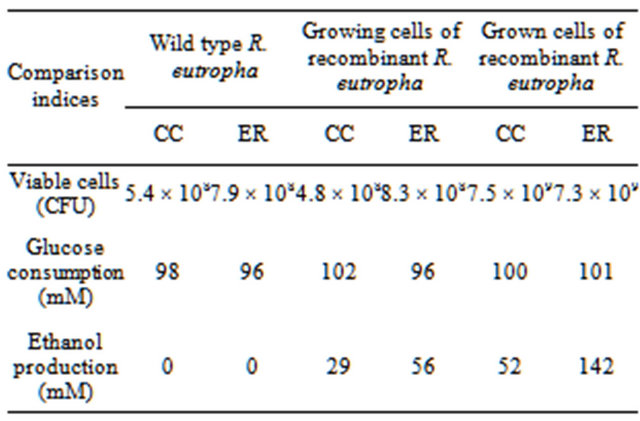
any metabolite from CO2 except biomass [1,13]. In this research, R. eutropha was genetically modified with pyruvate decarboxylase and alcohol dehydrogenase to obtain a recombinant strain capable of producing ethanol from glucose. The recombinant R. eutropha produced ethanol from glucose, about two times increased in the electrochemically reduced culture condition. The recombinant R. eutropha, the autotrophic CO2 fixation, and the electrochemical bioreactor will be combined to produce ethanol from CO2 in near future.
4. Conclusion
R. eutropha autotrophically grew with the electrochemical reducing power and CO2, and ethanol production of the recombinant from glucose was activated by the electrochemical reducing power. On the basis of this result, it is possible that the recombinant R. eutropha may produce ethanol from CO2 with electrochemical reducing power in medium without glucose by application of a specific signal for induction of pyruvate decarboxylase and alcohol dehydrogenase genes.
5. Acknowledgements
This work was supported by the New & Renewable Energy of the Korea Institute of Energy Technology Evaluation and Planning (KETEP) grant funded by the Korea governmental Ministry of Knowledge Economy (2012T1001100334).
REFERENCES
- B. Y. Jeon, I. L. Jung and D. H. Park, “Enrichment and Isolation of CO2-Fixing Bacteria with Electrochemical Reducing Power as a Sole Energy Source,” Journal of Environmental Protection, Vol. 3, No. 1, 2012, pp. 55- 60.
- B. Y. Jeon, S. Y. Kim, Y. K. Park and D. H. Park, “Enrichment of Hydrogenotrophic Methanogens in Coupling with Methane Production Using Electrochemical Bioreactor,” Journal of Microbiology and Biotechnology, Vol. 19, No. 12, 2009, pp. 485-493.
- D. H. Park and J. G. Zeikus. “Utilization of Electrically Reduced Neutral Red by Actinobacillus succinogenes: Physiological Function of Neutral Red in Membrane-Driven Fumarate Reduction and Energy Conservation,” Journal of Bacteriology, Vol. 181, No. 8, 1999, pp. 2403-2410.
- D. H. Park, M. Laivenieks, M. V. Guettler, M. K. Jain and J. G. Zeikus, “Microbial Utilization of Electrically Reduced Neutral Red as the Sole Electron Donor for Growth and Metabolite Production,” Applied Environmental Microbiology, Vol. 65, No. 7, 1999, pp. 2912-2917.
- H. S. Kang, B. K. Na and D. H. Park, “Oxidation of Butane to Butanol Coupled to Electrochemical Redox Reaction of NAD+/NADH,” Biotechnogical Letters, Vol. 29, No. 8, 2007, pp. 1277-1280. doi:10.1007/s10529-007-9385-7
- B. Y. Jeon and D. H. Park, “Improvement of Ethanol Production by Electrochemical Redox Combination of Zymomonas mobilis and Saccharomyces cerevisiae,” Journal of Microbiology and Biotechnology, Vol. 20, No. 1, 2010, pp. 94-100.
- W. J. Lee and D. H. Park, “Electrochemical Activation of Nitrate Reduction to Nitrogen by Ochrobactrum sp. G3-1 Using a Noncompartmented Electrochemical Bioreactor,” Journal of Microbiology and Biotechnology, Vol. 19, No. 8, 2009, pp. 836-844.
- B. Bowien and B. Kusian, “Genetics and Control of CO2 Assimilation in the Chemoautotroph Ralstonia eutropha,” Archives of Microbiology, Vol. 17, No. 2, 2002, pp. 85-93. doi:10.1007/s00203-002-0441-3
- S. Sichwart, S. Hetzler, D. Bröker and A. Steinbüchel, “Extension of the Substrate Utilization Range of Ralstonia eutropha Strain H16 by Metabolic Engineering to Include Mannose and Glucose,” Applied Environmental Micryobiology, Vol. 77, No. 4, 2011, pp. 1325-1334. doi:10.1128/AEM.01977-10
- R. Repaske, “Nutritional Requirement for Hydrogenomonas eutropha,” Journal of Bacteriology, Vol. 83, No. 2, 1962, pp. 418-422.
- P. E. Stukus and B. T. Becicco, “Autotrophic and Heterotrophic Metabolism of Hydrogenomonas,” Journal of Bacteriology, Vol. 95, No. 4, 1968, pp. 1469-1475.
- B. T. Decicco and P. E. Stukus, “Autotrophic and Heterotrophic Metabolism of Hydrogenomonas: Regulation of Autotrophic Growth by Organic Substrates,” Journal of Bacteriology, Vol. 101, No. 2, 1970, pp. 339-345.
- B. Y. Jeon, I. L. Jung and D. H. Park, “Enrichment of CO2-Fixing Bacteria in Cylinder-Type Electrochemical Bioreactor with Built-In Anode Compartment,” Journal of Microbiology and Biotechnology, Vol. 21, No. 6, 2011, pp. 590-598.
- L. O. Ingram and T. Conway, “Expression of Different Levels of Ethanologenic Enzymes from Zymomonas mobilis in Recombinant Strains of Escherichia coli,” Applied Environmental Microbiology, Vol. 54, No. 2, 1988, pp. 397-404.
- H. J. Tsai, K. L. Lin, J. C. Kuo and S. W. Chen, “Highly Efficient Expression of Fish Growth Hormone by Escherichia coli Cells,” Applied Environmental Microbiology, Vol. 61, No. 11, 1995, pp. 4116-4119.
- L. O. Ingram, T. Conway, D. P. Clark, G. W. Sewell and J. F. Preston, “Genetic Engineering of Ethanol Production in Escherichia coli,” Applied Environmental Microbiology, Vol. 53, No. 10, 1987, pp. 2420-2425.
- S. T. Taghavi, D. Van der Lelie and M. Mergeay, “Electroporation of Alcaligenes eutropha with (Mega) Plasmids and Genomic DNA Fragments,” Applied Environmental Microbiology, Vol. 60, No. 10, 1994, pp. 3685- 3591.
- T. Conway, G. W. Sewell, Y. A. Osman and L. O. Ingram, “Cloning and Sequencing of the Alcohol Dehydrogenase П Gene from Zymomonas mobilis,” Journal of Bacteriology, Vol. 169, No. 6, 1987, pp. 2591-2597.
- J. Swings and J. De Ley, “The Biology of Zymomonas,” Bacteriological Review, Vol. 41, No. 1, 1977, pp. 1-46.
- J. E. McGhee, G. St. Julian and R. W. Detroy, “Continuous and Static Fermentation of Glucose to Ethanol by Immobilized Saccharomyces cerevisiae Cells of Different Ages,” Applied Environmental Micryobiology, Vol. 44, No. 1, 1982, pp. 19-22.
- S. Bringer-Meyer and H. Sahm, “Metabolic Shifts in Zymomonas mobilis in Response to Growth Conditions,” FEMS Microbiological Review, Vol. 54, No. 2, 1988, pp. 131-142. doi:10.1111/j.1574-6968.1988.tb02739.x
- S. Bringer, R. K. Finn and H. Sahm. “Effect of Oxygen on the Metabolism of Zymomonas mobilis,” Archives of Microbiology, Vol. 139, No. 4, 1984, pp. 376-381. doi:10.1007/BF00408383
- D. H. Park and J. G. Zeikus, “Electricity Generation in Microbial Fuel Cells Using Neutral Red as an Electronophore,” Applied Environmental Microbiology, Vol. 66, No. 4, 2000, pp. 1292-1297. doi:10.1128/AEM.66.4.1292-1297.2000
NOTES
*Corresponding author.