Open Journal of Marine Science
Vol.05 No.02(2015), Article ID:55925,9 pages
10.4236/ojms.2015.52019
Sterile Surfaces of Mnemiopsis leidyi (Ctenophora) in Bacterial Suspension― A Key to Invasion Success?
Sven Hammann1,2*, Anthony Moss3, Martin Zimmer1,4
1Zoological Institute, Christian-Albrechts-University Kiel, Kiel, Germany
2Thuenen-Institute for Sea Fisheries, Hamburg, Germany
3Auburn University, Biological Sciences, Auburn University, Auburn, USA
4Leibniz-Center for Tropical Marine Ecology, Bremen, Germany
Email: *sven.hammann@ti.bund.de
Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 26 March 2015; accepted 20 April 2015; published 24 April 2015
ABSTRACT
Seawater is a dense microbial suspension with >106 prokaryotic and >104 eukaryotic propagules per milliliter. Hence, submerged surfaces get immediately covered by biofilm-forming colonizers upon contact with seawater. Since biofilms may reduce individual fitness through decreasing motility and attractiveness or increasing shearing stress by water currents and infection risk by pathogens, marine organisms have evolved countermeasures to regulate the number of surface-co- lonizers; alternatively they tolerate settlement and biofilm-formation. Antimicrobial defense mechanisms co-evolved with potentially colonizing microbes. By contrast, non-native animals (neozoa) are confronted with novel microbial colonizers upon colonizing a new habitat, and are expected to be less well protected against surface-colonization. Here we present results of a thorough screening of the epithelial surface of the ctenophore Mnemiopsis leidyi, being non-native in European marine environments, for epithelial bacteria and archaea. Neither light- and electron- microscopic inspection nor PCR-screening for bacterial and archaeal DNA of 134 adult specimens from different collection sites in the Western Baltic revealed any presence of prokaryotes on the surface epithelium of comb jellies in a recently invaded environment. A limited number of bacterial associates became evident from whole-body extracts of both juvenile and adult comb jellies. Their taxonomic diversity, however, was significantly lower in adult than in juvenile specimens, suggesting a maturation of anti-microbial defense upon ontogenetic development. The mechanisms underlying the effective defense of Mnemiopsis against microbial colonization, however, remain unknown. Based on our findings, we propose 1) to make use of invasion events as natural space-for-time experiments on how symbiotic interactions change upon environmental change; and 2) to study basal metazoan animals, such as ctenophores, to understand the evolutionary basics of symbiont-host interactions.
Keywords:
Ctenophora, Immune Response, Invasive Species, Space-For-Time, Symbiotic Interactions

1. Introduction
The significance of symbiont-host interactions for ecological processes is currently receiving increasing attention with particular focus on how they are shaped by environmental conditions [1] . Symbiotic interactions, irrespective of whether the symbiont exerts negative (parasites and pathogens) or positive (mutualists), or no (commensals) effects on its host, clearly affect the performance of at least one of the partners. To this end, environmental conditions mediate the outcome of symbiotic interactions [2] . Thus, a mutualistic symbiont in a given environment may become a parasite in another environment, or vice versa. Effects of changing environments on ecological interactions and eco-physiological processes can be studied in natural space-for-time experiments that make use of organisms that are currently spreading to colonize previously unexplored habitats (neobiota). In particular, invasive species provide an excellent opportunity for studying the effects of changing environments on symbiont-host interactions. Upon invasion of a new habitat, both partners face changes in their biotic and abiotic environment simultaneously, and symbiont-host interactions and established symbionts may change in quality and exclusiveness. Mnemiopsis leidyi, a common lobate ctenophore native to the Western Atlantic [3] recently invaded the North and Baltic Seas [4] -[7] , after it had previously been reported from the Black Sea and the Caspian Sea [8] -[10] . Dunn et al. [11] and Hejnol et al. [12] provided convincing molecular evidence for Ctenophora to represent the phylogenetic base of metazoans, being the sister group of all other metazoan taxa (see also: [13] ). Hence, M. leidyi may be ideally suited for studying the early evolution of metazoan interactions with symbionts and environmentally-induced changes in host-symbiont interactions in an evolutionary context. In addition, invading Mnemiopsis offer an opportunity to study real-time changes in elemental ecological and physiological interactions between a host and its symbionts in the context of successful adaptation to a changing environment. As it invades new habitats, Mnemiopsis and any associated co-introduced species must face novel environmental conditions and biotic interactions. Ctenophores bear a variety of invertebrate parasites. Amphipods of the suborder Hyperiidea (Crustacea: Amphipoda) are commonly found attached to the external epidermis [14] [15] , while the metacercaria of three digenean trematode families (Trematoda: Digenea: Faustulidae, Lepocreadiidae, Hemiuridae) are commonly found embedded in the mesoglea, often concentrated in regions close to the subplate canals. Such parasites mature in fish predators of ctenophores and commonly occur with remarkable prevalence and infestation intensity [16] . Mnemiopsis leidyi uniquely harbors the parasitic vermiform larva of the sea anemone Edwardsiella lineata (although it will transfer to the ctenophore predator Beroe ovata via predation [17] ). Little is known about the host-specificity of symbionts and knowledge on how the host is affected by the symbiotic association is essentially lacking. Furthermore, there is virtually no information regarding the geographical distribution of these metazoan symbionts of ctenophores in relation to the distribution of the host. Protists appear to be common epithelial symbionts of ctenophores [18] -[20] . According to Moss et al. [20] , mutualistic, parasitic as well as commensalic protists are associated with Mnemiopsis mccradyi, which is probably a synonym of M. leidyi. Parasitic protists, being taxonomically related to symbiotic or free- living protists in a variety of habitats, appear to inhabit well-defined body regions of their host, i.e., exclusively the surface of the comb plates or limited regions of the ectoderm. Environmental conditions may affect the suitability of ctenophores as potential hosts [21] [22] . In particular, reduced salinity appears to promote infestation by protists. It has been proposed that high infestation rates may affect Mnemiopsis fluid handling performance [20] . As has been shown for numerous invertebrates, close associations with bacteria are common in both aquatic and terrestrial environments (for recent review, see [23] ); in some cases, high specificity by both partners has been explicitly demonstrated (e.g. [24] -[26] ). Clearly, opportunistic and obligate symbionts co-exist and share the same host. So far, only one bacterial symbiont of Mnemiopsis (mccradyi) has been described [27] . Rod- shaped bacteria live inside a ciliary structure within the food groove area; their role is entirely unknown. Thus, little is known about bacterial associations with ctenophores, although preliminary reports by Moss and students (unpubl.) seem to indicate that Mnemiopsis has few to no bacteria on the epidermis while the gut contains bacteria characteristic of the free-living populations (Moss, unpublished results). By contrast Daniels & Breitbart [28] found a temporally variable microbiota that, however, clearly differed from free-living communities in the surrounding water column, and Dinsquet et al. [29] identified specific communities associated with M. leidyi tissue and gut. It is likely that the effects that bacterial symbionts exert on their host are mediated by environmental conditions and the resulting status of the host. Thus, interactions between Mnemiopsis and bacteria that are advantageous for the former within its natural range may become deleterious when conditions in a newly encountered habitat generate stress on the host. In contrast, parasitic bacteria could become beneficial to the host in a new environment, although this latter scenario seems unlikely. More likely, encounters with novel bacteria in an invaded environment could establish entirely new associations. Alternatively, the inability to establish, or the ability to prevent, such novel associations may decrease or improve the chance of successful invasion, respectively. There is precedence for this: Sánchez et al. [30] described enhanced invasive activity of New World Artemia species given their lack of susceptibility to Mediterranean parasites. Diversity of parasitic microsporidia has also been observed to be reduced in invasive amphipods [31] . Based on this, we hypothesize that symbiont-host associations of an invasive host species routinely change in quantity and quality upon invading novel habitats, and that this specifically happens in one of the most invasive coastal marine animals: the lobate Comb Jelly, Mnemiopsis leidyi, native at the coasts of North- and South America. In this first study, we screened individuals of M. leidyi from a recently invaded European region (the Baltic Sea) with respect to epithelial prokaryotic symbionts, and were unable to detect any metazoan symbiont associated with M. leidyi specimens in the Baltic Sea.
2. Materials and Methods
2.1. Molecular Identification of Ctenophore Bacterial Populations
We sampled M. leidyi through bucket-capture (“dipping”) of near-surface specimens and plankton-netting (100 µm) from greater depths during ship cruises at 4 different times during a major bloom throughout 2009 at 5 different Western Baltic sampling sites: Kiel Bight (54˚28'2"N, 10˚23'80"E; 20.07.09, 02.10.09, 30.10.09); Kiel Bight (54˚28'6"N, 10˚13'70"E; 20.07.09, 02.10.09, 30.10.09); Kiel Bight (54˚28'6"N, 10˚11'40"E; 20.07.09, 02.10.09, 30.10.09); Eckernfoerde Bight (54˚50'7"N, 09˚38'06"E; 02.10.09); Fehmarn Island (54˚37'2"N, 10˚41'94"E; 10.08.09). At every sampling site three water controls-1 liter each-were taken. Every water sample was mixed out of three samples from 3 different water depths (surface, near-bottom, and intermediate). Larvae of M. leidyi are <1 cm in oral-aboral lengths and typically bear two tentacles, which were lost during ontogenesis (see Figure 1). Captured adult specimen were typically between 2 and 8 cm oral-aboral lengths. After cap-
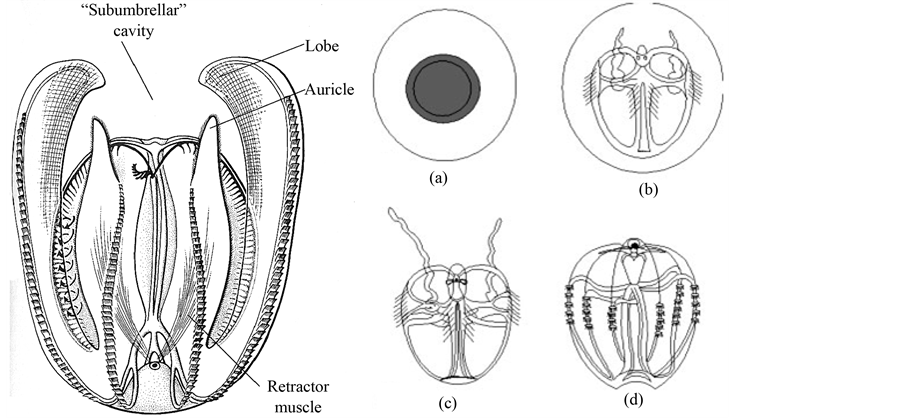
Figure 1. Mnemiopsis leidyi as an adult (left) and on the right side the development from egg (a) to larvae (c) and (d).
ture, the animals were identified visually rinsed carefully with sterile sea water and either immediately surface- swabbed for screening for epithelial symbionts, or frozen at −20˚C for whole-body extracts. Swabs were stored in 98% ethanol until DNA extraction. Before the animals were frozen they were kept for starving in sterile artificial seawater to empty their stomachs-inspected visually. DNA was extracted from sterile surface swabs, whole-body homogenates, water samples, and five times from the rinsing water following Henne et al. ([32] ; modified by R. Schmitz-Streit; adapted for comb jelly samples by S. Hammann). In brief, 1.35 mL DNA Extraction Buffer (DEB: 100 mM Tris, 100 mM sodium-EDTA, 100 mM sodium phosphate, 1.5 M NaCl, 1% CTAB, pH 8.0) was added to 0.5 to 1.5 mL sample. After adding 5 µl of 20 mg/mL ProteinaseK, samples were rotated for 30 min in a water bath at 37˚C. Afterwards, 1.5 µL of RNase (7 U/µL) were added. After incubation for 30 min at 37˚C, 150 µL of 20% SDS were added, and samples were inverted every 10 - 15 min for 2 h at 65˚C. After centrifugation (12 min; 10,000 g; RT) the supernatant was transferred for further processing, whereas the pellet was treated again with DEB (450 µL) and 20% SDS (50 µL), inverted and incubated for 10 min at 65˚C. Again, samples were centrifuged (12 min; 10,000 g; RT) and the supernatant was merged with the first supernatant. After adding 900 µL chloroform:isoamylalcohol (24:1) and inversion, samples were centrifuged for 20 min at 10,000 g (RT). Aliquots of 750 µL of the supernatant were mixed with the same volume of isopropanol, inverted and incubated overnight at 4˚C. After centrifugation (20 min; 10,000 g; 4˚C), the pellet was resuspended in 200 µL ice-cold 70˚C ethanol, inverted and centrifuged for 2 min at 10,000 g (4˚C). Subsequently, the pellet was air-dried and resuspended in 50 µL TE overnight. Although this method is commonly used for DNA extraction from soil samples it turned out to be the most efficient and stable method for extraction DNA from comb jellies. For cleaning extracted DNA, samples were brought to 200 µL with double-distilled water and mixed with an equal volume of phenol-chloroform. After vortexing and centrifugation (1 min, 10,000 g, RT), the upper phase was mixed with phenol-chloroform again, centrifuged, and the lower phase discarded. Then, 1/10 of the volume of 3 M sodium-acetate and three volumes of absolute ethanol were added and incubated overnight at −20˚C. After centrifugation (12 min; 10,000 g; RT), the pellet was resuspended in 50 µL TE overnight. The additional cleaning step was necessary because of PCR inhibiting effects of either remaining salts from the sea water or remaining mucus of the animals. DNA contents were measured before and after the cleaning steps using the NanoDrop® ND-1000 Universal bacterial PCR primers (27f and 1492r) were used to amplify the region corresponding to positions 27 - 1492 of the E. coli 16S rRNA gene. For archaeal DNA amplification the primers A20f and A958r were used. PCR was performed with the PromegaGoTaq Green Master Mix. In order to detect and amplify small amounts of bacterial DNA, PCR followed a touch-down protocol: 94˚C: 3 min; (94˚C: 30 s; 65˚C: 30 s; (ΔT = −0.5˚C/cycle); 24 cycles with 72˚C, 90 s; 15 cycles with 94˚C: 30 s, 54˚C: 30 s, 72˚C: 90 s; 72˚C: 5 min; hold at 4˚C. The resulting PCR fragments were cloned into pGEMT vector (Promega, Madison, WI) and transformed into DH5a E. coli cells (Invitrogen, Karlsruhe, Germany) according to the manufacturers’ instructions. Sanger-sequencing was done by the Institute for Clinical Molecular Biology (IKMB), Kiel. The sequences were edited and analyzed with the following programs: BioEdit, MEGA, DNAMAN, Pintail, sequin. For the web research and sequence (and therefore species-) identifiction, the BlastSearch of the NCBI was used. For the estimation of bacterial phylotype diversity, we used the Chao1 nonparametric richness estimator [33] implemented in EstimateS (version 8.2, http://purl.oclc.org/estimates), treating each comb jelly specimen as a separate sample [25] .
2.2. Electron Microscopy
Electron-microscopic screening of the epithelial surface was performed at the Zoological Institute of Kiel University, Functional Morphology and Biomechanics. For fixation of comb jelly epithelia for scanning electron microscopy, whole individuals were directly transferred into liquid nitrogen for shock-freezing. After 5 min, the specimens were transferred into 50 mL tubes (Beckton-Dickinson: “Falcon”) and stored at −70˚C to await further processing. For scanning electron microscopy, specimens were fixed onto glass slides, sputter-coated with a 10 nm layer of gold-palladium alloy, and subsequently observed under a cryo-electron-microscope (Hitachi S- 4800 equipped with a GATAN cryo-apparatus).
3. Results
In total, 144 clones were sequenced. Whole-body extracts (N = 14) of juvenile M. leidyi from the Baltic Sea yielded 8 different sequences of amplified bacterial 16S rRNA genes that could be affiliated with diverse bacterial classes (Figure 2). Based on this, we estimated a bacterial diversity (Chao1 estimator) of 20 + 11 (mean + S.D.) in this category. From 30 whole-body extracts of adult Baltic Sea M. leidyi we obtained another 8 different phylotypes that belonged to either α-(3) or γ-(5) Proteobacteria (Figure 2), suggesting specific host-symbiont-associations with a bacterial diversity (Chao1 estimator) of as low as 8 + 3.
Remarkably, at most only a single bacterial phylotype was associated with any given adult individual, and in >85% of the whole-body extracts we could not detect any bacterial DNA signature (Figure 3). None of the phylotypes we found in adults were shared with larval M. leidyi, and only one of them was also found in Baltic water samples; three larva-associated phylotypes were also detected in water samples. As indicated by PCR- analysis of surface swabs, none of the bacteria associated with adult M. leidyi resides on the surface of the umbrella, since neither bacterial nor archaeal rRNA gene sequences could be amplified from the surface of 134 adult specimens from the Western Baltic Sea.
The lack of prokaryotes on the umbrella surface was also corroborated by electron-microscopic inspection of M. leidyi specimen collected in the Baltic Sea (Figure 4).
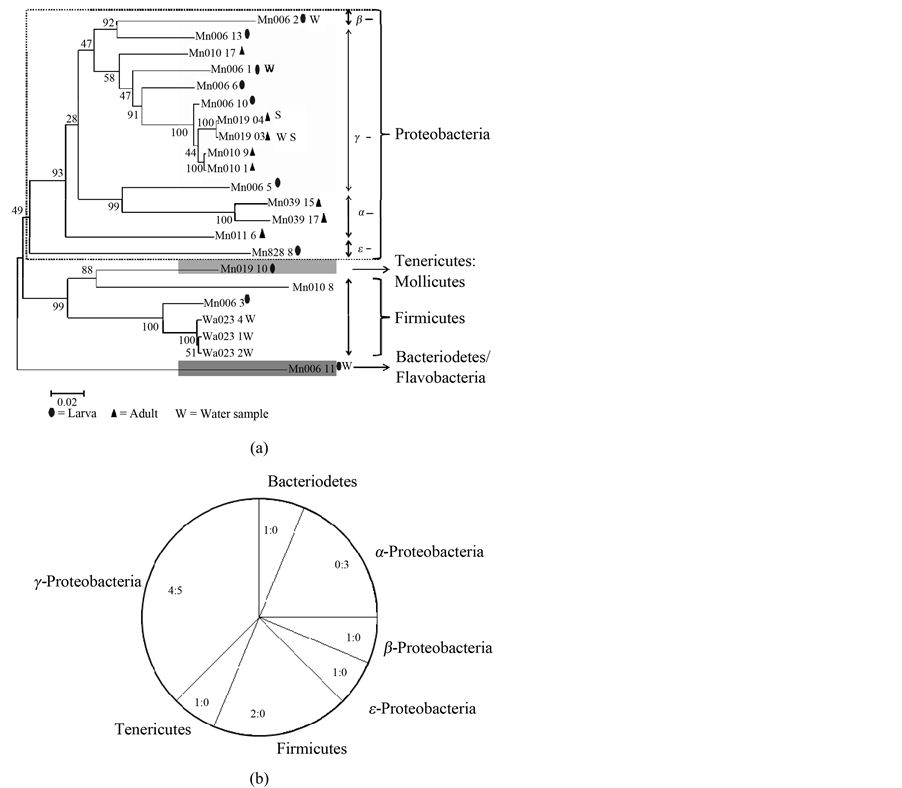
Figure 2. (a) Phylogenetic affiliation of bacterial 16S rRNA gene sequences obtained from whole-body extracts of larval (ellipsoid; N = 14) and adult (triangle; N = 30) Mnemiopsis leidyi, and the surrounding water column (w). Neighbor-Joining-Tree with bootstrap 1000, generated with MEGA 4.1 Beta. (b) Summary of (a), showing the numbers of phylotypes representative of different bacterial classes that were found in whole-body extracts of larval (L) and adult (A) Mnemiopsis leidyi; data are presented as L:A.

Figure 3. Mean (+S.D.) number of bacterial phylotypes (left ordinate) in whole- body extracts of individual larvae (N = 14) and adults (N = 30) of Mnemiopsis leidyi, and proportion of larvae (left) and adults (right) that did (gray area) or did not (white area) harbor prokaryotic symbionts (right ordinate).
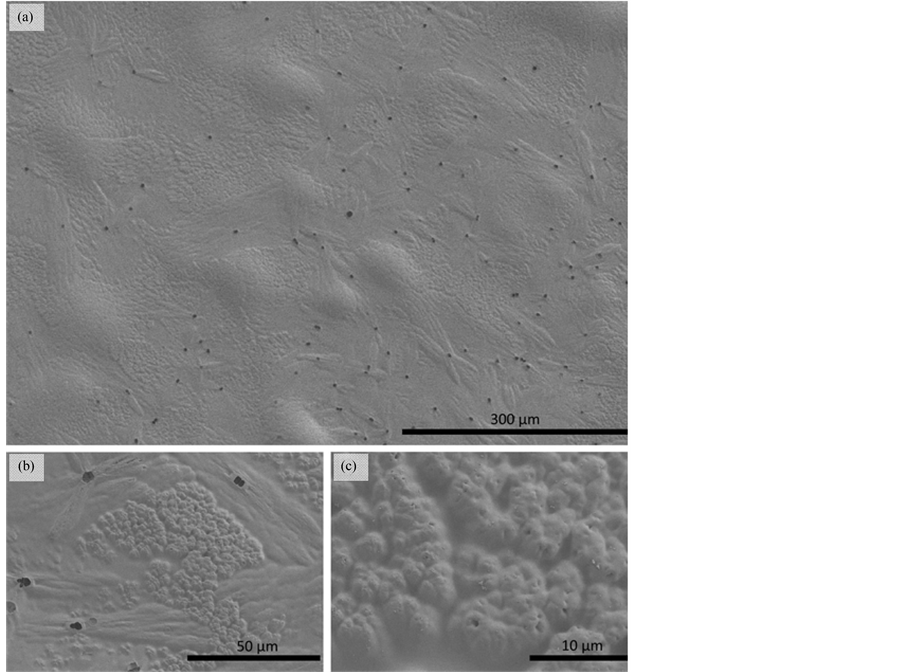
Figure 4. Representative SEM-micrograph of epithelial surfaces of Mnemiopsis leidyi, demonstrating the absence of bacteria and archaea from the jelly’s umbrella with (a) 150×, (b) 1000× and (c) 4500× enlargement, respectively.
4. Discussion
Essentially every submerged surface is immediately covered by biofilm-forming bacterial, archaeal or protist colonizers upon contact with seawater. According to our careful screening of the umbrella surface of 134 adult specimens by means of PCR and electron microscopy, we conclude that the epithelial umbrella surface of Mnemiopsis leidyi is void of both bacteria and archaea in the recently invaded Western Baltic. Irrespective of this finding of the present study, this species appears to harbor a specific microbiota that clearly differs from the bacterial community of the water column [28] [29] . Mnemiopsis leidyi of the east coast of southern US have been described to harbor specific epithelial protists. The mobiline peritrich, Trichodina ctenophorii [19] and small Flabellula-like gymnamoebae inhabit the surface of the comb plates, whereas Vexillifera-like gymnamoebae and Protoodinium-like dinoflagellates are common all over the umbrella epithelium [20] . Except for the gymnamoebae, we did not find these protists upon microscopic inspection of individual from the Baltic Sea (authors’ unpubl. observation) and tentatively suggest that these protist symbionts have been lost at some stage of invading Baltic water bodies. Reasons for, and consequences of, this change in symbiotic associations remain to be studied in detail. However, it is obvious from our findings that M. leidyi exhibits a highly effective defense, be it chemically or mechanically, that prevents its epithelial surface from being colonized by microbes from the surrounding sea water. Invasion events may be considered natural space-for-time experiments on changes in biotic interactions under changing environmental conditions. Along this line, we propose that the ability to maintain a symbiont-free epithelial surface is a key to invasion success [30] . We view this hypothesis as extension of the enemy-release hypothesis that had been put forward to explain why invasive species may be superior to native species [34] and has subsequently been tested repeatedly [35] -[38] . Here, we argue that mechanisms that keep epithelial surfaces void of symbionts help protect invaders against potential parasitic or pathogenic symbionts in their novel environment. Williamson and Fitter’s [39] Tens Rule for invasive species estimates that on average only 10 % of potentially invasive species make it to the next step of successful invasion [40] , even though conditions appear appropriate for neobionts. To this end, it is tempting to conclude that by changing environment, the host introduces the usual community of associated symbionts to such harsh conditions that it loses them. In addition, because the host enters an environment with an assemblage of naive parasites, the host does not present a significant, easily colonized surface. In circumstances where the symbiont is supportive of the host, loss of the symbiont upon invasion would be expected to negatively regulate the invading host. In contrast to the above extended enemy-release hypothesis, Mnemiopsis leidyi may be an exceptionally successful invader because it already lacks any prokaryotic epithelial symbionts upon which it depends in its native range. Preliminary results indicate a complete lack of surface-attached prokaryotes in New World M. leidyi [41] . These two hypotheses are not mutually exclusive, and further studies are needed to decide upon this issue. Our present data on whole-body extracts clearly indicate that there are indeed bacteria associated with M. leidyi, but at low diversity. Corroborating our present findings, Daniels and Breitbart [28] and Dinasquet et al. [29] presented evidence for a much lower bacterial richness and diversity in Mnemiopsis whole-body extracts than in the surrounding sea water. According to the present findings, adults are associated with a limited diversity of bacteria (estimated 8 + 3), whereas the diversity of bacteria associated with juveniles is about 2.5-times higher. Daniels and Breitbart [28] observed seasonal variation in the diversity of bacterial communities associated with Mnemiopsis but did not distinguish between adults and juveniles. Dinasquet et al. [29] found specific bacterial communities in the gut and tissue of M. leidyi. Again there is no differentiation between adult and larval specimen. In addition to Daniels and Breitbart [28] , they sampled surface tissue, but in contrast to our study were the surface was swapped they dissected the animals tissues and included epithelium and parts of the mesogloea as well. Our findings suggest that larval Mnemiopsis are less well defended against microbial colonization than adults, but defense mechanisms mature during ontogenetic development. At current, we cannot specify where the bacterial symbionts we detected are situated. However, the obvious specificity suggests that they are either harbored inside the tissue or are specifically associated with the (gut) epithelium. We consider it unlikely that such low bacterial diversity reflects a bacterial community that is only loosely associated with the digestive tract and would be representative of the surrounding water column. The findings of Daniels and Breitbart [28] and Dinasquet et al. [29] support this hypothesis. Obviously, none of the bacterial symbionts, nor any archaea or protist is harbored on the epithelial surface of adult Mnemiopsis in the invaded Baltic Sea. The mechanism of keeping the epidermal surface void of prokaryotes remains unclear: neither polar (methanol) nor apolar (hexane) whole-body extracts exhibited any antibacterial activity on settlement or growth (M. Wahl and M. Zimmer, unpubl.). Thus, it remains to be clarified which surface characteristics of the comb jelly render the umbrella void of bacteria. We can at present only speculate on evolutionary and ecological consequences of a sterile epithelial surface on ctenophores. The phylum Ctenophora is currently considered an evolutionary basal group within the Metazoa, and so a sister taxon of all other Metazoa [11] [12] . We propose to study Mnemiopsis more closely to understand better the evolutionary basis of metazoan/microbe-interactions and the evolution of the metazoan immune system. Our results here provoke the question: If basal Metazoa (e.g., Ctenophora) are capable of maintaining their epithelium void of bacterial (and archaeal) colonizers, why is it then advantageous for all other Metazoa to host epithelial symbionts? Alternatively, the ability to prevent epithelial colonization may have evolved in Ctenophora after phylogenetic separation of all other Metazoa. Preventing colonization may have arisen for very simple reasons, related to the microanatomy of this group. Preventing colonization will eliminate the ready attachment of degradative microbes as well as any activities arising via symbioses, which may be very important in Mnemiopsis in particular, because it samples benthic sediments [42] . Any colonization that could accelerate microbe-induced degradation would be extremely dangerous for these organisms, which fundamentally consist of a thin monolayer epidermal “skin” surrounding a thick, nutrient rich mesoglea [43] , [44] and Moss, unpubl. ultrastructural studies. It is well-established that symbiosis is a significant driver of animal evolution and diversification [45] -[48] . Considering that symbioses have led to ontogenetic innovations in many organisms, having led to all modern phyla, we propose that preventing bacterial colonization is very probably a novel evolutionary pathway, and yet one that obviously has proved to be fundamentally evolutionarily advantageous. It may be that by adopting this mechanism, perhaps as a means of self-protection, ctenophores have undergone much less change with time. This strategy may have led to these ancient organisms persisting from deep time, with few changes, to the present day. By contrast, the Cnidaria, which are commonly improperly grouped with the Ctenophora, demonstrate many thousands of species, unlike the very limited diversity of species seen in the Ctenophora [49] . They also display enormous diversity within each subgroup, and are famous for their extensive associations with microbes. For instance corals are reliant upon both eukaryotic and prokaryotic symbionts and are more correctly today referred to as a holobiont―i.e., a complex multispecies super-organism― and are greatly affected by changes in those relationships, which are themselves exceptionally sensitive to photic, thermal, chemical, genetic and mechanically-based environmental changes [1] [50] -[52] . By completely avoiding prokaryotic interactions, ctenophores may have gone the evolutionarily safe way but may have inadvertently slowed their evolutionary progress. Future work will be directed at elucidating the evolution of ancient cellular and immunological response mechanisms, thereby taking advantage of the unique characteristics of this enigmatic and fascinating group of animals.
Acknowledgements
Stanislav N. Gorb and Marie-Christin Klein, University of Kiel (Germany) provided lab and office space, along with invaluable support, to S.H. for electron microscopy; Martin Wahl, GEOMAR (Kiel, Germany) provided office and lab space to M.Z. We are grateful for initial funding of our project through grants from the Cluster of Excellence Future Ocean (Kiel, Germany) to A.G.M. (for a start-up visit) and M.Z. The authors are indebted to the Thuenen Institute for Sea Fisheries for financial support.
References
- Weis, V.M. (2008) Cellular Mechanisms of Cnidarian Bleaching: Stress Causes the Collapse of Symbiosis. Journal of Experimental Biology, 211, 3059-3066.
- Steinert, M., Hentschel, U. and Hacker, J. (2000) Symbiosis and Parthogenesis: Evolution of the Microbe-Host Interaction. Naturwissenschaften, 87, 1-11. http://dx.doi.org/10.1007/s001140050001
- Mayer, A.G. (1912) Ctenophores of the Atlantic Coast of North America. Carnegie Institution of Washington, (Publ. 162), 58 pp. http://dx.doi.org/10.5962/bhl.title.5968
- Faasse, M.A. and Bayha, K.M. (2006) The Ctenophore Mnemiopsis leidyi A. Agassiz 1865 in Coastal Waters of the Netherlands: An Unrecognized Invasion? Aquatic Invasions, 1, 270-277. http://dx.doi.org/10.3391/ai.2006.1.4.9
- Hansson, H.G. (2006) Ctenophores of the Baltic and Adjacent Seas―The Invader Mnemiopsis Is Here! Aquatic Invasion, 1, 295-298. http://dx.doi.org/10.3391/ai.2006.1.4.16
- Javidpour, J., Sommer, U. and Shiganova, T. (2006) First Record of Mnemiopsis leidyi A. Agassiz 1865 in the Baltic Sea. Aquatic Invasions, 1, 299-302. http://dx.doi.org/10.3391/ai.2006.1.4.17
- Boersma, M., Malzahn, A., Greve, W. and Javidpour, J. (2007) The First Occurrence of the Ctenophore Mnemiopsis leidyi in the North Sea. Helgoland Marine Research, 61, 153-155. http://dx.doi.org/10.1007/s10152-006-0055-2
- Vinogradov, M.E., Shushkina, E.A., Musayeva, E.I. and Sorokin, P.Y. (1989) A New Exotic Species in the Black Sea: The Ctenophore Mnemiopsis leidyi (Ctenophora: Lobata). Oceanology, 29, 220-224.
- Studenikina, Y.I., Volovik, S.P., Mirzoyan, I.A. and Luts, G.I. (1991) The Ctenophore Mnemiopsis leidyi in the Sea of Azov. Oceanology, 31, 722-725.
- Shiganova, T.A. (1993) Ctenophore Mnemiopsis leidyi and Ichthyoplankton in the Sea of Marmara in October of 1992. Oceanology, 33, 900-903.
- Dunn, C.W., Hejnol, A., Matus, D.Q., Pang, K., Browne, W.E., Smith, S.A., et al. (2008) Broad Phylogenomic Sampling Improves Resolution of the Animal Tree of Life. Nature, 452, 745-749. http://dx.doi.org/10.1038/nature06614
- Hejnol, A., Obst, M., Stamatakis, A., Ott, M., Rouse, G.W., Edgecombe, G.D., et al. (2009) Assessing the Root of Bilaterian Animals with Scalable Phylogenomic Methods. Proceedings of the Royal Society B, 276, 4261-4270. http://dx.doi.org/10.1098/rspb.2009.0896
- Wallberg, A., Thollesson, M., Farris, J.S. and Jondelius, U. (2004) The Phylogenetic Position of the Comb Jellies (Ctenophora) and the Importance of Taxonomic Sampling. Cladistics, 20, 558-578. http://dx.doi.org/10.1111/j.1096-0031.2004.00041.x
- Sorarrain, D.R., Ramirez, F. and Mianzan, H. (2001) Hyperoche medusarum (Krøyer, 1838) (Amphipoda, Hyperiidae) and Mnemiopsis mccradyi (Mayer, 1910) (Ctenophora): A New Host and First Record of This Association for the Southwestern Atlantic. Crustaceana, 74, 407-410. http://dx.doi.org/10.1163/156854001300104499
- Gasca, R. and Haddock, H.D. (2004) Associations between Gelatinous Zooplankton and Hyperiid Amphipods (Crustacea: Peracarida) in the Gulf of California. Hydrobiologia, 530-531, 529-535. http://dx.doi.org/10.1007/s10750-004-2657-5
- Martorelli, S.R. (2001) Digenea Parasites of Jellyfish and Ctenophores of the Southern Atlantic. Hydrobiologia, 451, 305-310. http://dx.doi.org/10.1023/A:1011862406670
- Reitzel, A.M., Sullivan, J.C., Brown, B.K., Chin, D.W., Cira, E.K., Edquist, S.K., Genco, B.K., Joseph, O.C., Kaufman, C.A., Kovitvongsa, K., Mu-oz, M.M., Negri, T.L., Taffel, J.R., Zuehlke, R.T. and Finnerty, J.T. (2007) Ecological and Developmental Dynamics of a Host-Parasite System Involving a Sea Anemone and Two Ctenophores. Journal of Parasitology, 93, 1392-1402. http://dx.doi.org/10.1645/GE-1250.1
- Mills, C.A. and McLean, N. (1991) Ectoparasitism by a Dinoflagellate (Dinoflagellata: Oodinidae) on 5 Ctenophores (Ctenophora) and a Hydromedusa (Cnidaria). Diseases of Aquatic Organisms, 10, 211-216. http://dx.doi.org/10.3354/dao010211
- Estes, A.M., Reynolds, B.S. and Moss, A.G. (1997) Trichodina ctenophorii N. Sp., a Novel Symbiont of Ctenophores of the Northern Coast of the Gulf of Mexico. Journal of Eukaryotic Microbiology, 44, 420-426. http://dx.doi.org/10.1111/j.1550-7408.1997.tb05718.x
- Moss, A.G., Estes, A.M., Muellner, L.A. and Morgan, D.D. (2001) Protistan Epibionts of the Ctenophore Mnemiopsis mccradyi Mayer. Hydrobiologia, 451, 295-304. http://dx.doi.org/10.1023/A:1011846022599
- Khan, R.A. (1990) Parasitism in Marine Fish after Chronic Exposure to Petroleum Hydrocarbons in the Laboratory and to the Exxon Valdez Oil Spill. Bulletin of Environmental Contamination and Toxicology, 44, 759-763. http://dx.doi.org/10.1007/BF01701799
- Khan, R.A., Berker, D.E., Williams-Ryan, K. and Hooper, R.G. (1993) Influence of Crude Oil and Pulp and Paper Mill Effluent on Mixed Infections of Trichodina cottidarium and T. saintjohnsi (Ciliophora) Parasitizing Myoxiocephalus octodecemspinosus and M. scorpius. Canadian Journal of Zoology, 72, 247-251. http://dx.doi.org/10.1139/z94-033
- Dale, C. and Moran, N.A. (2006) Molecular Interactions between Bacterial Symbionts and Their Hosts. Cell, 126, 453- 465. http://dx.doi.org/10.1016/j.cell.2006.07.014
- Moran, N. and Baumann, P. (2000) Bacterial Endosymbionts in Animals. Current Opinion in Microbiology, 3, 270- 275. http://dx.doi.org/10.1016/S1369-5274(00)00088-6
- Fraune, S. and Bosch, T.C.G. (2007) Long-Term Maintenance of Species-Specific Bacterial Microbiota in the Basal Metazoan Hydra. Proceedings of the National Academy of Sciences of the USA, 104, 13146-13151. http://dx.doi.org/10.1073/pnas.0703375104
- Fraune, S. and Zimmer, M. (2008) Host-Specificity of Environmentally Transmitted Mycoplasma-Like Isopod Symbionts. Environmental Microbiology, 10, 2497-2504. http://dx.doi.org/10.1111/j.1462-2920.2008.01672.x
- Moss, A.G., Rapoza, R.C. and Muellner, L. (2001) A Novel Cilia-Based Feature within the Food Grooves of the Ctenophore Mnemiopsis mccradyi Mayer. Hydrobiologia, 451, 287-294. http://dx.doi.org/10.1023/A:1011833618965
- Daniels, C. and Breitbart, M. (2012) Bacterial Communities Associated with the Ctenophores Mnemiopsis leidyi and Beroe ovata. FEMS Microbiology Ecology, 82, 90-101. http://dx.doi.org/10.1111/j.1574-6941.2012.01409.x
- Dinasquet, J., Granhag, L. and Riemann, L. (2012) Stimulated Bacterioplankton Growth and Selection for Certain Bacterial Taxa in the Vicinity of the Ctenophore Mnemiopsis leidyi. Frontiers in Microbiology, 3, 302. http://dx.doi.org/10.3389/fmicb.2012.00302
- Sánchez, M.I., Rode, N.O., Flaven, E., Redón, S., Amat, F., Vasileva, G.P. and Lenormand, T. (2012) Differential Susceptibility to Parasites of Invasive and Native Species of Artemia Living in Sympatry: Consequences for the Invasion of A. franciscana in the Mediterranean Region. Biological Invasions, 14, 1819-1829. http://dx.doi.org/10.1007/s10530-012-0192-2
- Slothouber, J.G.M., Smith, G.J.E., Becnel, J.J., Butlin, R.K. and Dunn, A.M. (2010) Reduction in Post-Invasion Genetic Diversity in Crangonyx pseudogracilis (Amphipoda: Crustacea): A Genetic Bottleneck or the Work of Hitchhiking Vertically Transmitted Microparasites? Biological Invasions, 12, 191-209. http://dx.doi.org/10.1007/s10530-009-9442-3
- Henne, A., Daniel, R., Schmitz, R.A. and Gottschalk, G. (1999) Construction of Environmental DNA Libraries in Escherichia coli and Screening for the Presence of Genes Conferring Utilization of 4-Hydroxybutyrate. Applied and Environmental Microbiology, 65, 3901-3907.
- Chao, A. (1984) Nonparametric Estimation of the Number of Classes in a Population. Scandinavian Journal of Statistics, 11, 265-270.
- Elton, C.S. (1958) The Ecology of Invasions by Animals and Plants. Methuen, London. http://dx.doi.org/10.1007/978-1-4899-7214-9
- Keane, R.M. and Crawley, M.J. (2002) Exotic Plant Invasions and the Enemy Release Hypothesis. Trends in Ecology and Evolution, 17, 164-170. http://dx.doi.org/10.1016/S0169-5347(02)02499-0
- Mitchell, C.E. and Power, A.G. (2003) Release of Invasive Plants from Fungal and Viral Pathogens. Nature, 421, 625- 627. http://dx.doi.org/10.1038/nature01317
- De Walt, S.J., Denslow, J.S. and Ickes, K. (2004) Natural-Enemy Release Facilitates Habitat Expansion of the Invasive Tropical Shrub Clidemia hirta. Ecology, 85, 471-483. http://dx.doi.org/10.1890/02-0728
- Blumenthal, D.M. (2006) Interactions between Resource Availability and Enemy Release in Plant Invasion. Ecology Letters, 9, 887-895. http://dx.doi.org/10.1111/j.1461-0248.2006.00934.x
- Williamson, M. and Fitter, A. (1996) The Varying Success of Invaders. Ecology, 77, 1661-1666. http://dx.doi.org/10.2307/2265769
- Lockwood, J.L., Hoopes, M. and Marchetti, M. (2007) Invasion Ecology. Blackwell Publishers, Oxford.
- Donovan, E.W., Dodson, M.B. and Moss, A.G. (2010) Bacterial Assemblages of the Lobate Ctenophore, Mnemiopsis. Proceedings of the Annual Meeting American Society for Limnology and Oceanography, Santa Fe, 6-11 June 2010.
- Taylor, J.C., Odom, D.C. and Moss, A.G. (2006) Benthic Interactions in Mnemiopsis. Proceedings of the Annual Meeting of the American Society of Limnology and Oceanography, Victoria, 4-9 June 2006.
- Hernandez-Nicaise, M.-L. (1973) The Nervous System of Ctenophores III. Ultrastructure of Synapses. Journal of Neurocytology, 2, 249-263. http://dx.doi.org/10.1007/BF01104029
- Hernandez-Nicaise, M.-L. (1974) Ultrastructural Evidence for a Sensory-Motor Neuron in Ctenophora. Tissue and Cell, 6, 43-47. http://dx.doi.org/10.1016/0040-8166(74)90021-4
- Doebeli, M. and Dieckmann, U. (2000) Evolutionary Branching and Sympatric Speciation Caused by Different Types of Ecological Interactions. The American Naturalist, 156, 77-101. http://dx.doi.org/10.1086/303417
- Buckling, A. and Rainey, P.B. (2002) The Role of Parasites in Sympatric and Allopatric Host Diversification. Nature, 420, 496-499. http://dx.doi.org/10.1038/nature01164
- Leonardo, T.E. and Muiru, G.T. (2003) Facultative Symbionts Are Associated with Host Plant Specialization in Pea Aphid Populations. Proceedings of the Royal Society of London B, 270, S209-S212. http://dx.doi.org/10.1098/rsbl.2003.0064
- Watson, R.A. and Pollack, J.B. (2003) A Computational Model of Symbiotic Composition in Evolutionary Transitions. Biosystems, 69, 187-209. http://dx.doi.org/10.1016/S0303-2647(02)00135-1
- Mills, C.E. (1998) Phylum Ctenophora: List of all Valid Species Names. http://faculty.washington.edu/cemills/Ctenolist.html
- Morrow, K.M. and Chadwick, N. (2010) Competition among Sessile Organisms on Coral Reefs. In: Dubinsky, Z. and Stambler, N., Eds., Coral Reefs: An Ecosystem in Transition, Springer, New York.
- Morrow, K.M., Moss, A.G., Chadwick, N. and Liles, M. (2012) Bacterial Associates of Two Caribbean Coral Species Reveal Species-Specific Distribution and Geographic Variability. Applied and Environmental Microbiology, 78, 6438- 6449. http://dx.doi.org/10.1128/AEM.01162-12
- Morrow, K.M., Liles, M.R., Paul, V.J., Moss, A.G. and Chadwick, N.E. (2013) Bacterial Shifts Associated with Coral-Macroalgal Competition at Three Caribbean Reef Locations. Marine Ecology Progress Series, 488, 103-117.
NOTES

*Corresponding author.