Identification of EDTA-Soluble Polysaccharides from Pea Epicotyl Cell Walls and
154
Their Interaction with Xyloglucan
studies indicated that a similar binding pattern occurs
between nascent GAX and pectin to xyloglucan extracted
from the third internodes of pea epicotyls, and that the
binding requires the presence of proteins named assem-
blins [5]. The association between the polymers would
probably not occur in Golgi vesicles prior to their arrival
at the plasma membrane, since the pH within Golgi vesi-
cles is thought to be close to neutrality. The present in-
vestigation aimed at investigating the interaction between
xyloglucan and polymers that have already been depos-
ited into the wall. The results obtained confirm that the
binding pattern between xyloglucan and EDTA-soluble
polysaccharides that have been extracted from the wall,
is similar to that reported between xyloglucan and nas-
cent EDTA-soluble polysaccharides obtained from Golgi
membranes [5]. To identify these polysaccharides, dif-
ferent enzyme treatments followed by gel filtration were
performed, in addition to acid hydrolysis followed by
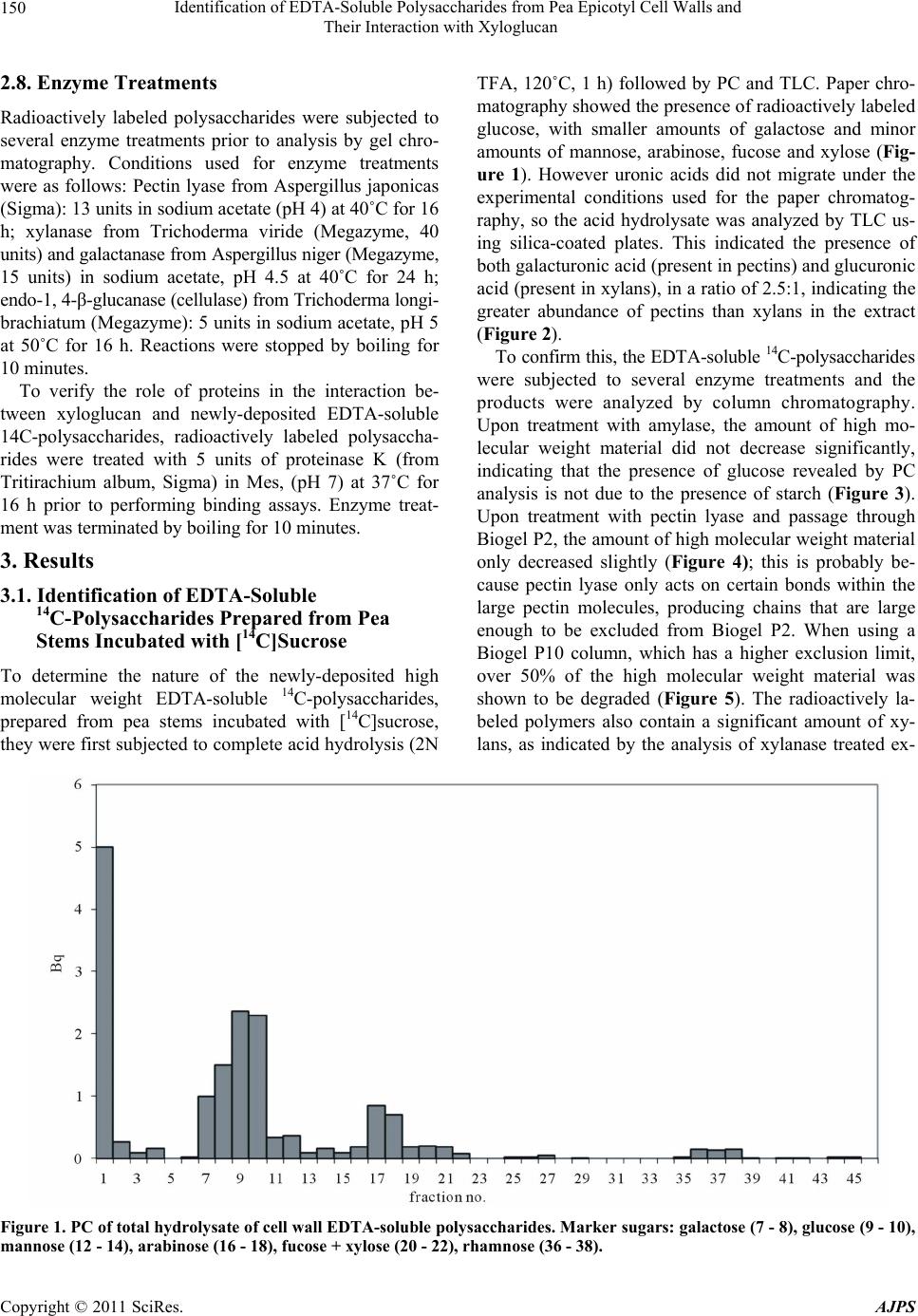
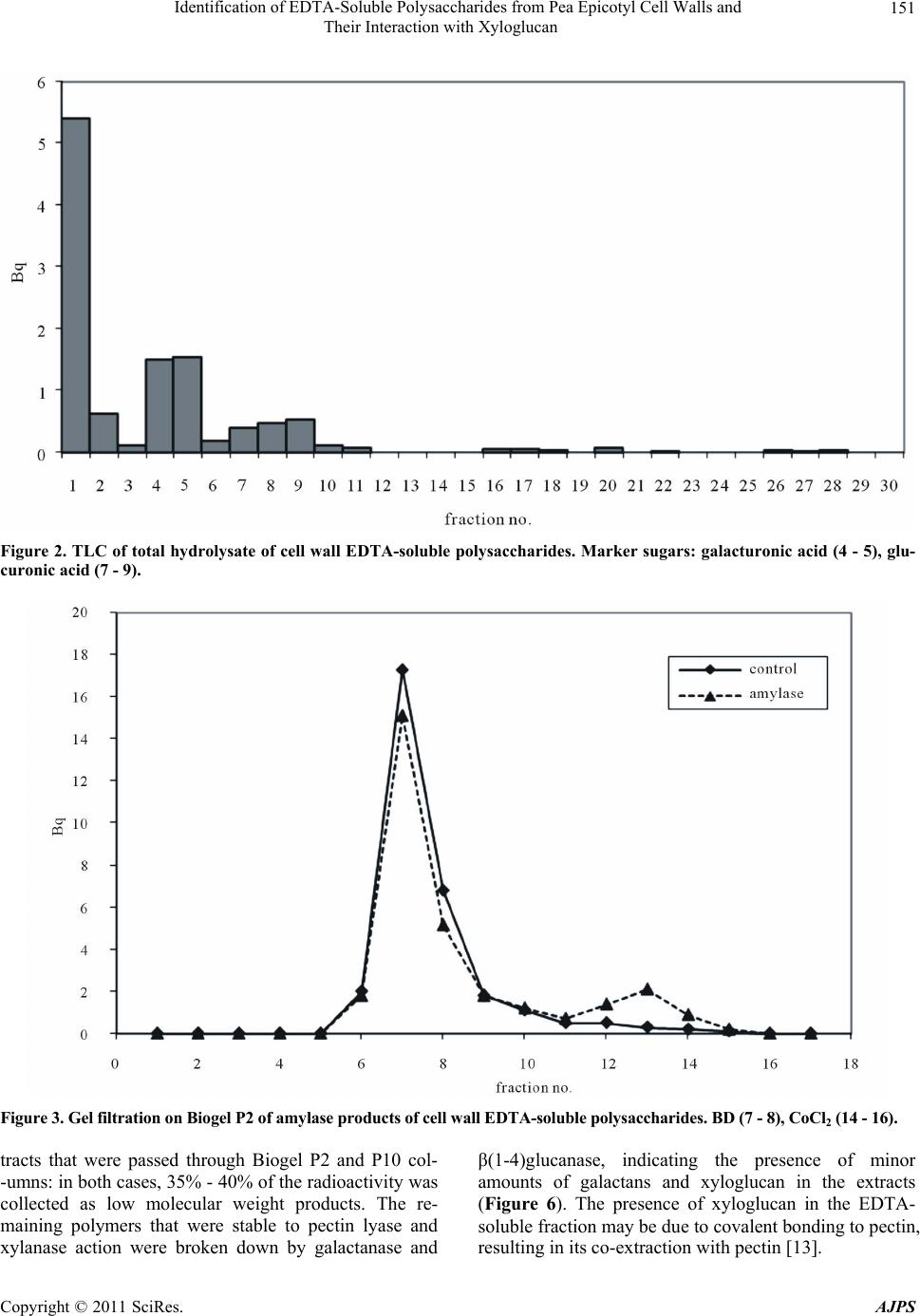
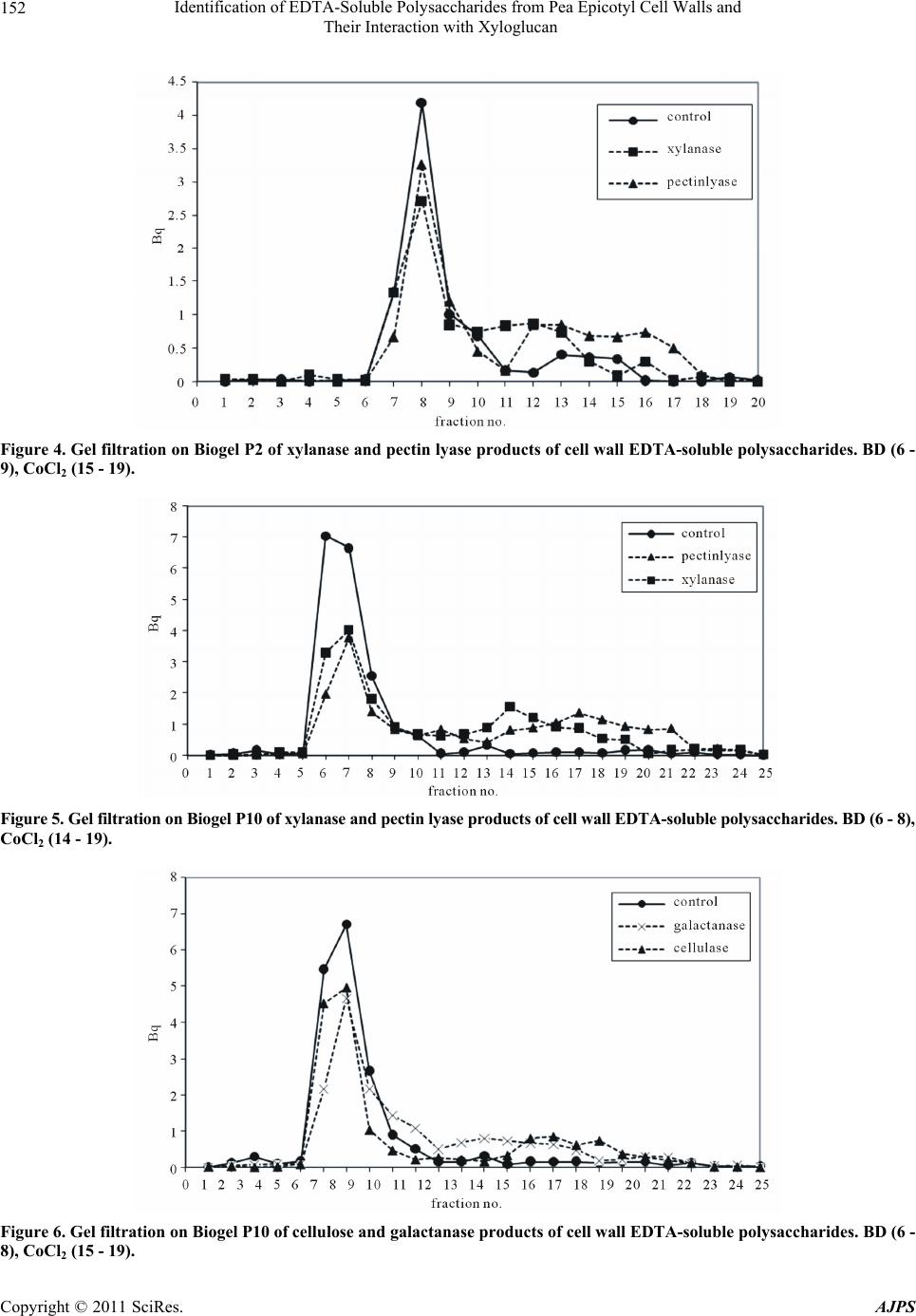
chromatography of the hydrlolysate. The results indi-
cated the newly-deposited polysaccharides consisted of
pectin, probably with some xyloglucan attached, and
smaller amounts of glucuronoxylan. These polysaccha-
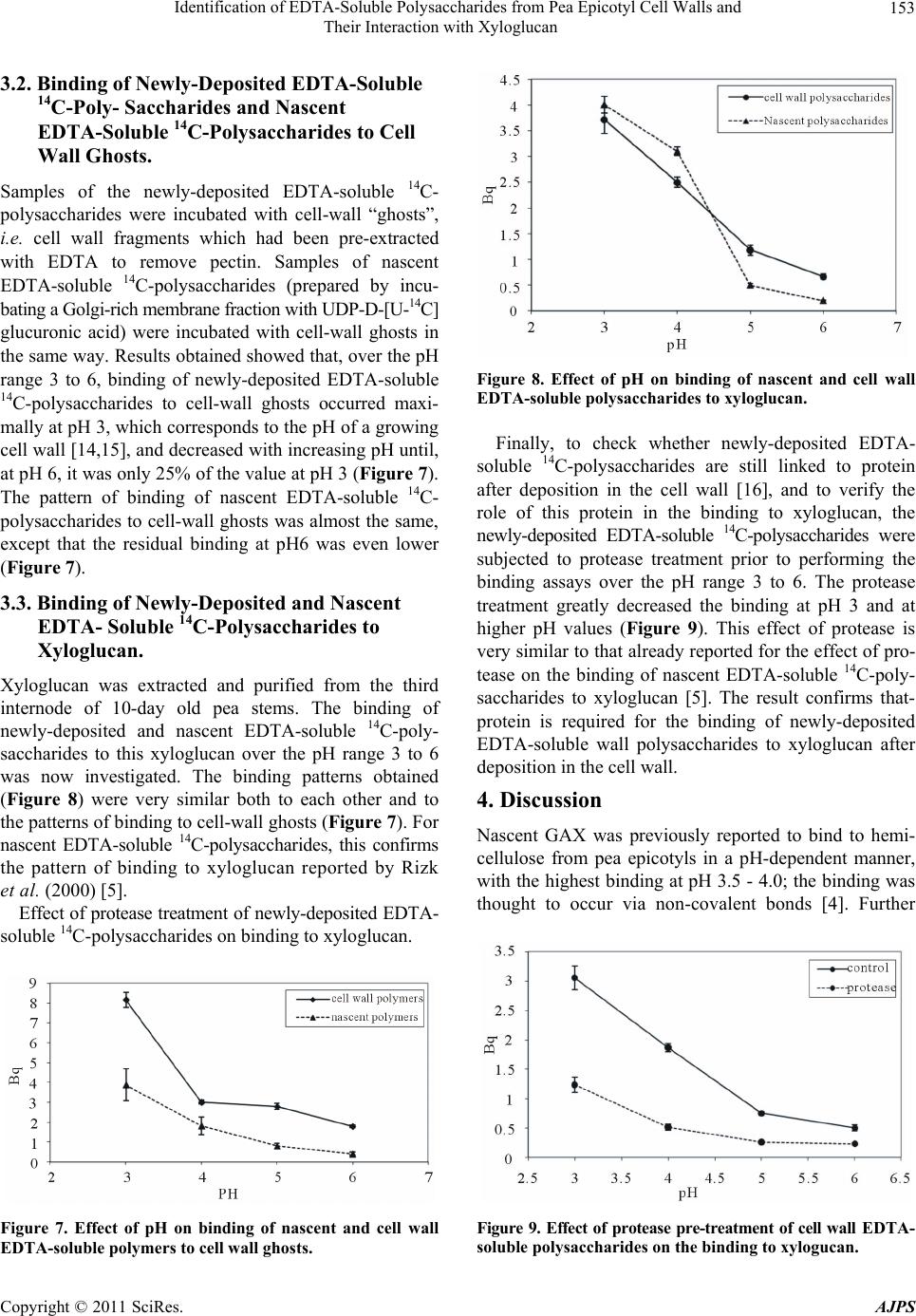
rides were found to bind to xyloglucan in a pH-depend-
ent manner, whereby pH 3 gave the highest binding over
the pH range tested, which is similar to that reported by
Rizk et al., 2000. This pH corresponds to that of a cell
wall undergoing growth, and lower pH values were not
tested because they would be of doubtful physiological
significance. The binding greatly decreased when the
polysaccharides were subjected to protease treatment
prior to performing the binding assay, confirming the
presence of proteins associated with these polymers after
their deposition into the wall.
This protein- and pH-dependent binding suggests a
functional interaction with the mechanisms that control
growth. In pea epicotyls, the pH of the wall decreases to
3 when growth is initiated [14]. Rapid growth may re-
quire strong interactions between matrix polymers in
order to maintain the cohesion of the wall. In addition,
pectin and GAX molecules present in the cell wall may
compete with cellulose for binding to xyloglucan at an
acidic pH, interfering with the strong xylogluca-
n-cellulose binding, and hence rendering the cell wall
more extensible to allow rapid growth.
REFERENCES
[1] A. Staehelin and I. Moore, “The Plant Golgi Apparatus:
Structure, Functional Organization and Trafficking Mecha-
nisms,” Annual Review of Plant Physiology and Plant
Molecular Bi ology, Vol. 46, 1995, pp. 261-288.
doi:10.1146/annurev.pp.46.060195.001401
[2] S. C. Fry, “Cellulases, Hemicelluloses and Auxinstimu-
lated Growth: A Possible Relationship,” Plant Physiology,
Vol. 75, No. 4, 1989, pp. 532-536.
doi:10.1111/j.1399-3054.1989.tb05620.x
[3] T. Hayashi, K. Ogawa and Y. Mitsuishi, “Characteriza-
tion of the Adsorption of Xyloglucan to Cellulose,” Plant
and Cell Physiology, Vol. 35, No. 8, 1994, pp. 1199-
1205.
[4] C. T. Brett, S. A. Healy, M. S. McDonald, C. Macgregor
and E. Baydoun, “Binding of Nascent Glucuronoxylan to
the Cell Walls of Pea Seedlings,” International Journal of
Biological Macromolecules, Vol. 21, No. 1-2, 1997, pp.
169-173. doi:10.1016/S0141-8130(97)00057-3
[5] S. Rizk, M. R. Abdel, E. Baydoun and C. T. Brett, “Pro-
tein- and pH-Dependent Binding of Nascent Glucurona-
rabinoxylan to Xyloglucan in Pea,” Planta, Vol. 211,
2000, pp. 423-429. doi:10.1007/s004250000303
[6] W. D. Reiter, “Biosynthesis and Properties of the Plant
Cell Wall,” Current Opinion in Plant Biology, Vol. 5, No.
6, 2002, pp. 536-542.
doi:10.1016/S1369-5266(02)00306-0
[7] T. Hayashi and G. Maclachlan, “Pea Xyloglucan and
Cellulose. I. Macromolecular Organization,” Plant Physi-
ology, Vol. 75, 1984, pp. 596-604.
doi:10.1104/pp.75.3.596
[8] K. Ogawa, T. Hayashi and K. Okamura, “Conformational
Analysis of Xyloglucan,” International Journal of Bio-
logical Macromolecules, Vol. 12, No. 3, 1990, pp. 218-
222. doi:10.1016/0141-8130(90)90036-A
[9] P. Kooiman, “A Method for Determination of Amyloid in
Plant Seeds,” Recueil des Travaux Chimiques des
Pays-Bas et de la Belgique, Vol. 79, 1960, pp. 675-678.
[10] T. Hayashi, Y. Kato and K. Matsuda, “Xyloglucan from
Suspension-Cultured Soybean Cells,” Plant and Cell
Physiology, Vol. 21, No. 8, 1980, pp. 1405-1418.
[11] K. W. Waldron and C. T. Brett, “Subcellular Localization
of a Glucuronyltransferase Involved in Glucuronoxylan
Biosynthesis in Pea (Pisum Sativum) Epicotyls,” Plant
Science, Vol. 49, No. 1, 1986, pp. 1-8.
doi:10.1016/0168-9452(87)90013-6
[12] S. C. Fry, “The Growing Plant Cell Wall: Chemical and
Metabolic Analysis,” John Wiley and Sons, New York,
1988.
[13] C. M. Cumming, H. D. Rizkallah, K. McKendrick, R. M.
Abdel-Massih, E. Baydoun and C. T. Brett, “Biosynthesis
and Cell-Wall Deposition of a Pectin-Xyloglucan Com-
plex in Pea,” Planta, Vol. 222, 2005, pp. 546-555.
doi:10.1007/s00425-005-1560-2
[14] R. E. Cleland, G. Buckley, S. Nowbar, N. M. Lew, C.
Stienmetz, M. L. Evans and D. L. Rayle, “The pH Profile
for Acid-Induced Elongation of Coleoptile and Epicotyl
Sections Is Consistent with the Acid-Growth Theory,”
Planta, Vol. 186, No. 1, 1991, pp. 70-74.
doi:10.1007/BF00201499
[15] U. Kutschera, “The Current Status of the Acid Growth
Theory,” New Phytologist, Vol. 126, No. 4, 1994, pp.
549-569. doi:10.1111/j.1469-8137.1994.tb02951.x
Copyright © 2011 SciRes. AJPS