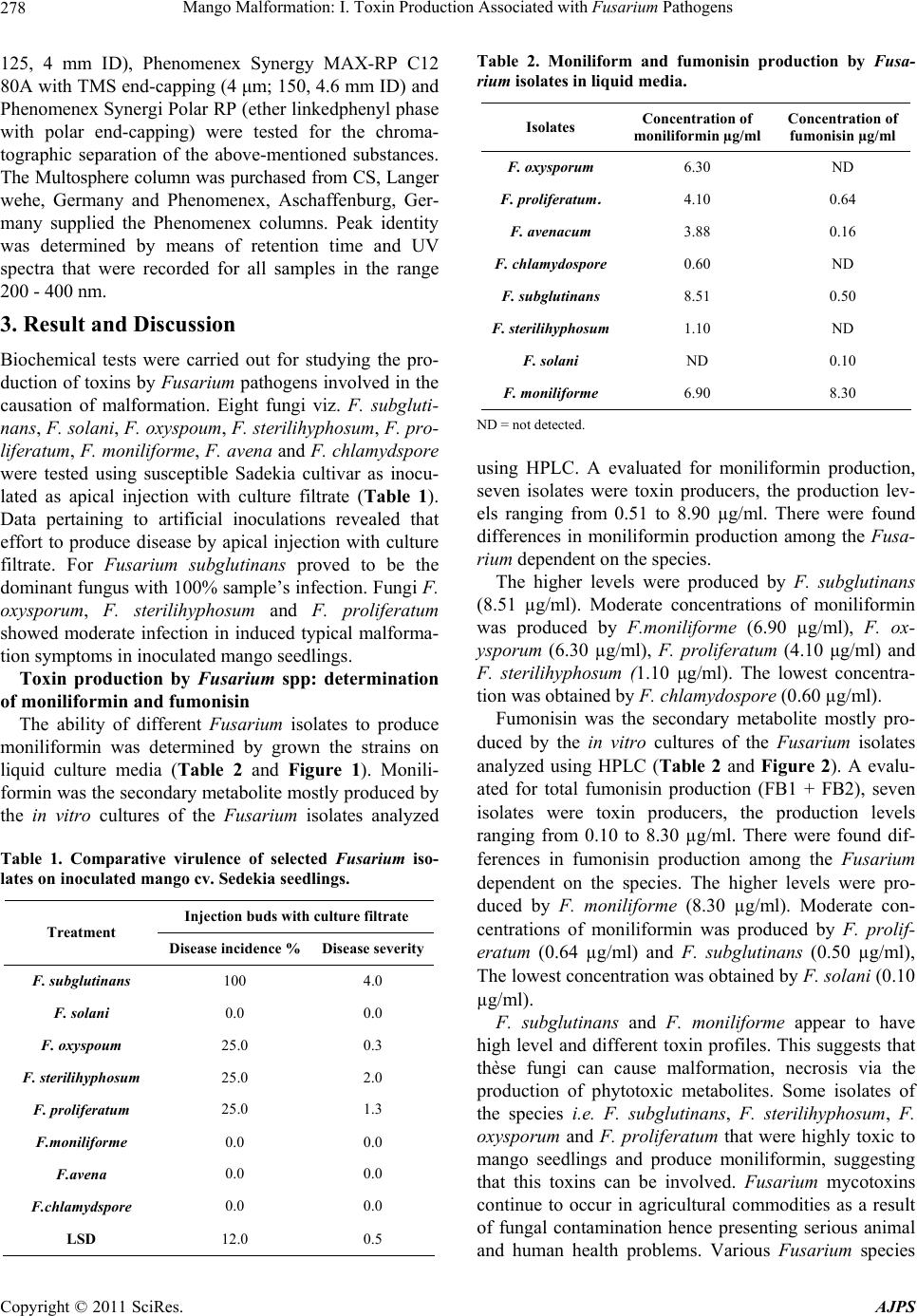
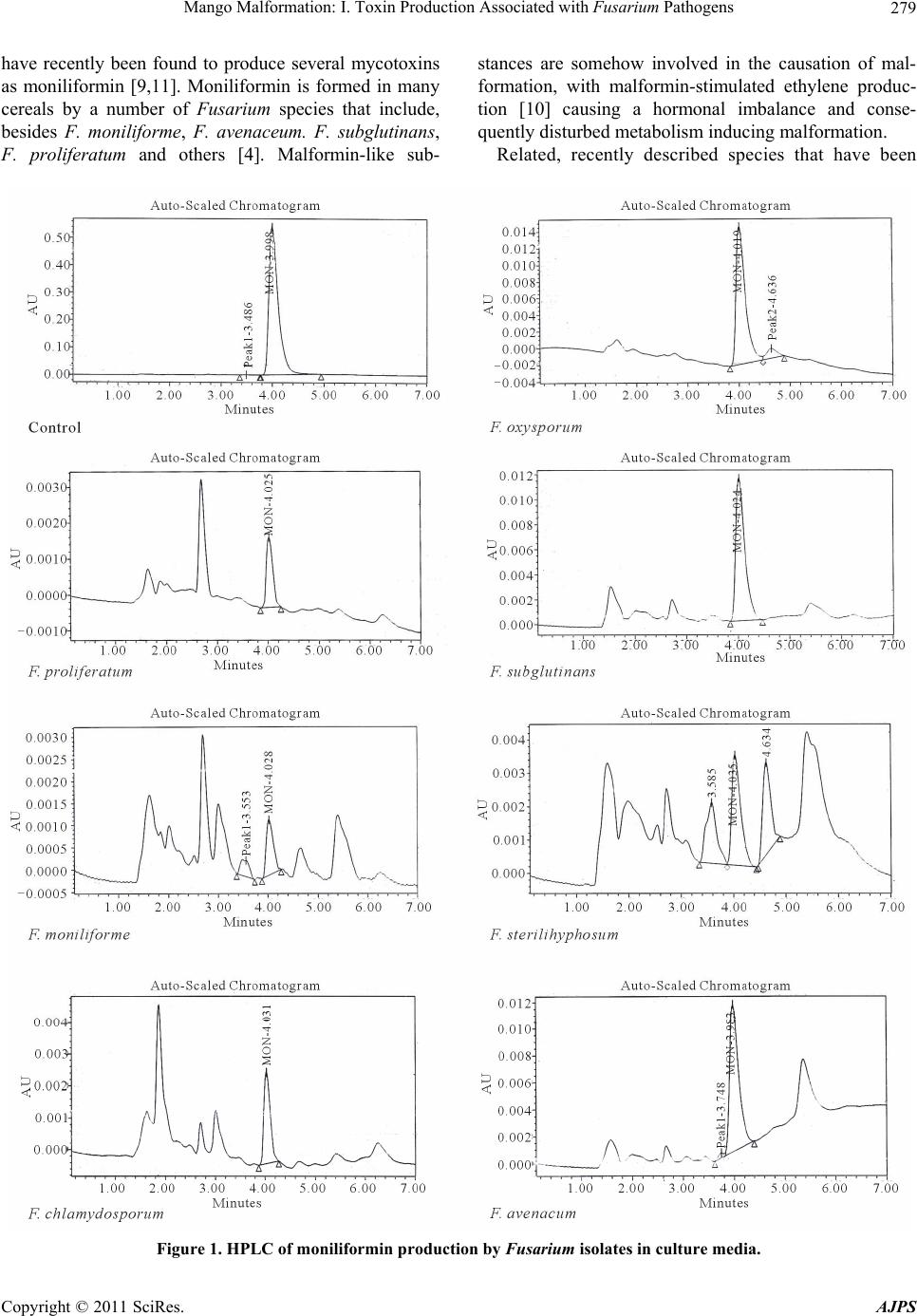
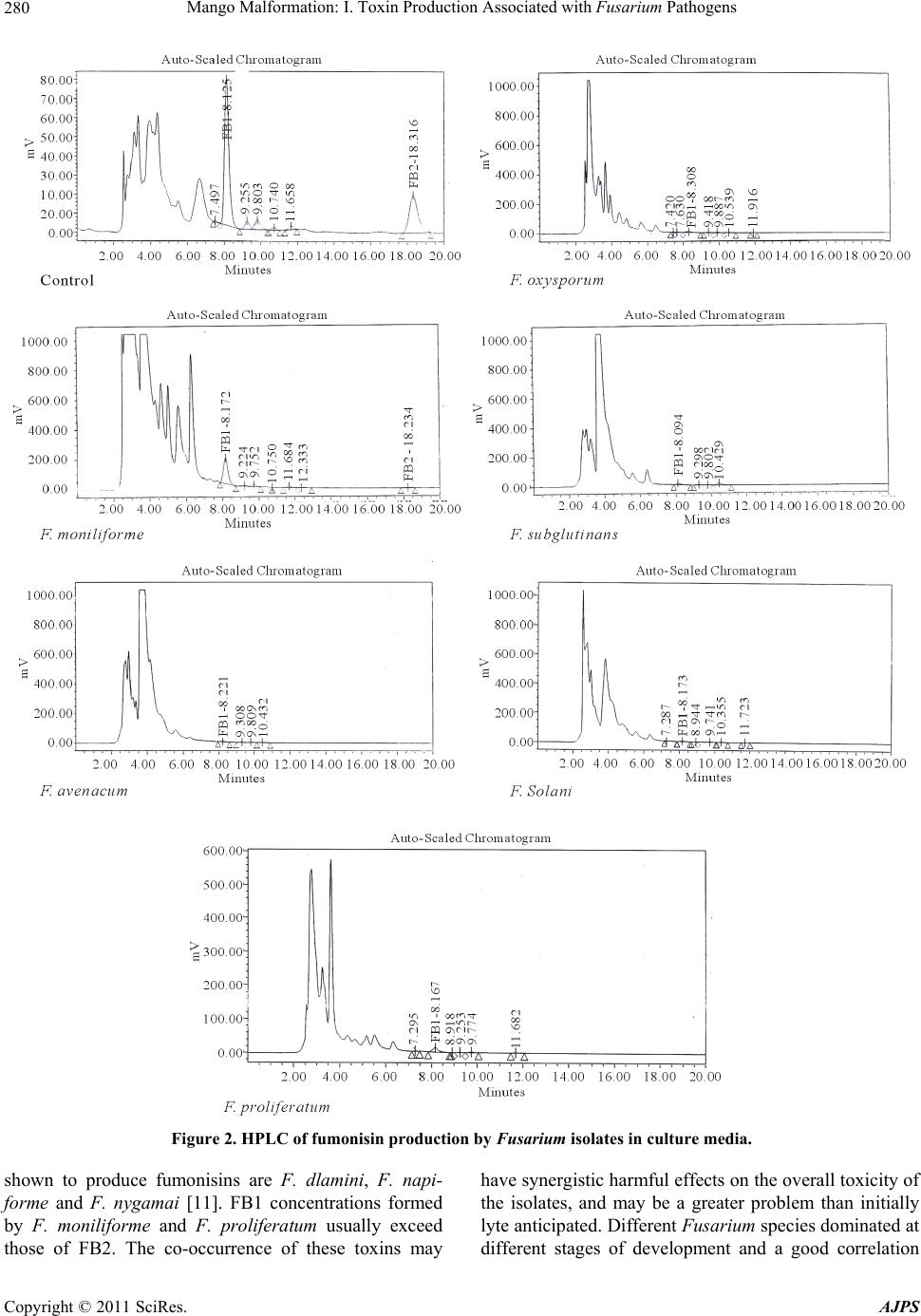
Mango Malformation: I. Toxin Production Associated with Fusarium Pathogens281
was found between fumonisins and the presence of F.
moniliforme and F. proliferatum. The occurrence of very
high levels of fumonisin B# in some samples was corre-
lated with the presence of strains producing abundant
fumonisin B# in the laboratory [16].
4. Acknowledgements
This manuscript funded from the project “New applied
approaches to promote productivity and Quality of some
fruit crops (Mango)” National Research Centre, 2007 to
2010.
REFERENCES
[1] R. C. Ploetz, “Malformation: A Unique and Important
Disease of Mango, Mangifera Indica L,” In: B. A. Sum-
merell, J. F. Leslie, D. Backhouse and W. L. Bryden,
Eds., Fusarium: Paul E. Nelson Memorial Symposium,
American Phytopathological Society (APS) Press, St
Paul, 2001, pp. 233-247.
[2] Z. Singh and B. S. Dhillon, “Comparative Developmental
Morphology of Normal and Malformed Floral Organs of
Mango (Mangifera IndicaL.),” Trop Agriculture, Trini-
dad, Vol. 67, 1990, pp. 143-148.
[3] J. Kumar and S. P. S. Beniwal, “A Method of Estimating
Cultivar Susceptibility against Mango Malformation,”
Tropical Pest Management, Vol. 33, 1987, pp. 208-210.
[4] M. I. Khaskheli, M. Pathan, M. Jiskani, M. H. Soomro
and G. B. Poussio, “First Record of Fusarium Nivale
(FR.) Ces. Associated with Mango Malformation Disease
(MMD) in Pakistan,” Pakistan Journal of Botany, Vol.
40, No. 6, 2008, pp. 2641-2644.
[5] W. M. Haggag and M. E. A. El-Wahab, “First Report of
Fusarium sterilihyphosum and F. Proliferatum-Induced
Malformation Disease of Mango in Egypt,” Journal of
Plant Pathology, Vol. 91, No. 1, 2009, pp. 231-240.
[6] J. Kumar and S. P. S. Beniwal, “Mango Malformation,”
In: J. Kumar, H. S. Chaube, U. S. Singh, A. N. Muk-
hopadhyay, Eds., Plant Diseases of International Impor-
tance, Prentice Hall, New York, Vol. 3, 1992, pp. 357-
393.
[7] Z. Singh and B. S. Dhillon, “Presence of Malformin-Like
Substance in Malformed Floral Tissues of Mango,”
Journal of Phytopathology, Vol. 125, No. 1, 1989a, pp.
17-123.
[8] S. Ram, “Horticultural Aspects of Mango Malformation,”
Acta Horticulture, Vol. 291, 1991, pp. 235-252.
[9] Z. Singh and B. S. Dhillon, “Effect of Naphthalene Acetic
Acid, Ethrel, Dikegulac and Hand Deblossoming on Flo-
ral Malformation, Flowering, Yield and Fruit Quality of
Mango (Mangifera Indica L.),” Journal of Phytopathol-
ogy, Vol. 120, 1986, pp. 245-248.
doi:10.1111/j.1439-0434.1987.tb04438.x
[10] Z. Singh and B. S. Dhillon, “Hormonal Changes Associ-
ated with Vegetative Malformation of Mango (Mangifera
indica L.),” Journal of Phytopathology, Vol. 125, No. 3,
1989b, pp. 193-197.
doi:10.1111/j.1439-0434.1989.tb01060.x
[11] P. E. Nelson, R. D. Plattner, D. D. Shackelford and A. E.
Desjardins, “Fumonisin B1 Production by Fusarium Spe-
cies other than F. Moniliforme in Section Liseola and by
Some Related Species,” Applied and Environmental Mi-
crobiology, Vol. 58, No. 3, 1992, pp. 984-989.
[12] J. D. Miller, M. E. Savard, A. Sibilia, S. Rapior, A. D.
Hocking and J. I. Pitt, “Production of Fumonisins and
Fusarins by Fusarium Moniliforme from Southeast Asia,”
Mycologia, Vol. 85, No. 3, 1993, pp. 385-391.
doi:10.2307/3760700
[13] M. E. Savard and J. D. Miller, “Characterization of Fusa-
rin F, a New Fusarin from Fusarium Moniliforme,” Jour-
nal of Natural Products, Vol. 55, No. 1, 1992, pp. 64-70.
doi:10.1021/np50079a010
[14] P. M. Scott and G. A. Lwrence, “Liquid Chromatographic
Determination and Stability of the Fusarium Mycotoxin
Moniliformin in Cereal Grains,” Journal of the Associa-
tion of Official Analytical Chemists, Vol. 70, No. 5, 1987,
pp. 850-853.
[15] J. D. Miller, “Epidemology of Fusarium Ear Diseases,”
In: J. D. Miller and H. L. Trenholm, Eds., Mycotoxins in
Grain, Eagan Press, St. Paul, 1994, pp. 19-36.
[16] S. N. Chulze, M. L. Ramirez, M. C. Farnochi, M. Pascale,
A. Visconti and G. March, “Fusarium and Fumonisins
Occurrence in Argentinian Corn at Different Ear Maturity
Stages,” Journal of Agricultural and Food Chemistry,
Vol. 44, No. 9, 1996, pp. 2797-2801.
doi:10.1021/jf950381d
Copyright © 2011 SciRes. AJPS