Plant Regeneration through Indirect Somatic Embryogenesis in Coelogyne Cristata Orchid263
mination for 3 months. The germinated plantlets about
1.0 cm in height were transferred to Murashige and
Skoog (MS) basal media supplemented with NAA 1.0
mg·L–1 and BA 2.0 mg·L–1 for plant growth. After 2
months of culture, leaves of the in vitro plantlets become
the explants source to conduct further experiments.
2.1. Effect of PGRs and Coconut Water on
Callus Induction from Leaf Segments
Leaf segments of about 3 - 5 mm in length were used as
explants. For investigations on the effects of coconut
water (5% v/v) (MB cell, Seoul, Korea) and plant growth
regulators (PGRs) on callus induction, various combina-
tions of 2,4-dichlorophenoxyacetic acid (2,4-D; 0, 1.0,
2.0 and 3.0 mg·L–1) and benzyladenine (BA; 0, 1.0, 2.0
and 3.0 mg·L–1) were added to MS basal media. Each
experiment consisted of 10 explants with three replica-
tions. The culture vessels were placed in dark conditions
for 12 weeks and forming calli were recorded.
2.2. Effect of Media and AC on Somatic
Embryogenesis
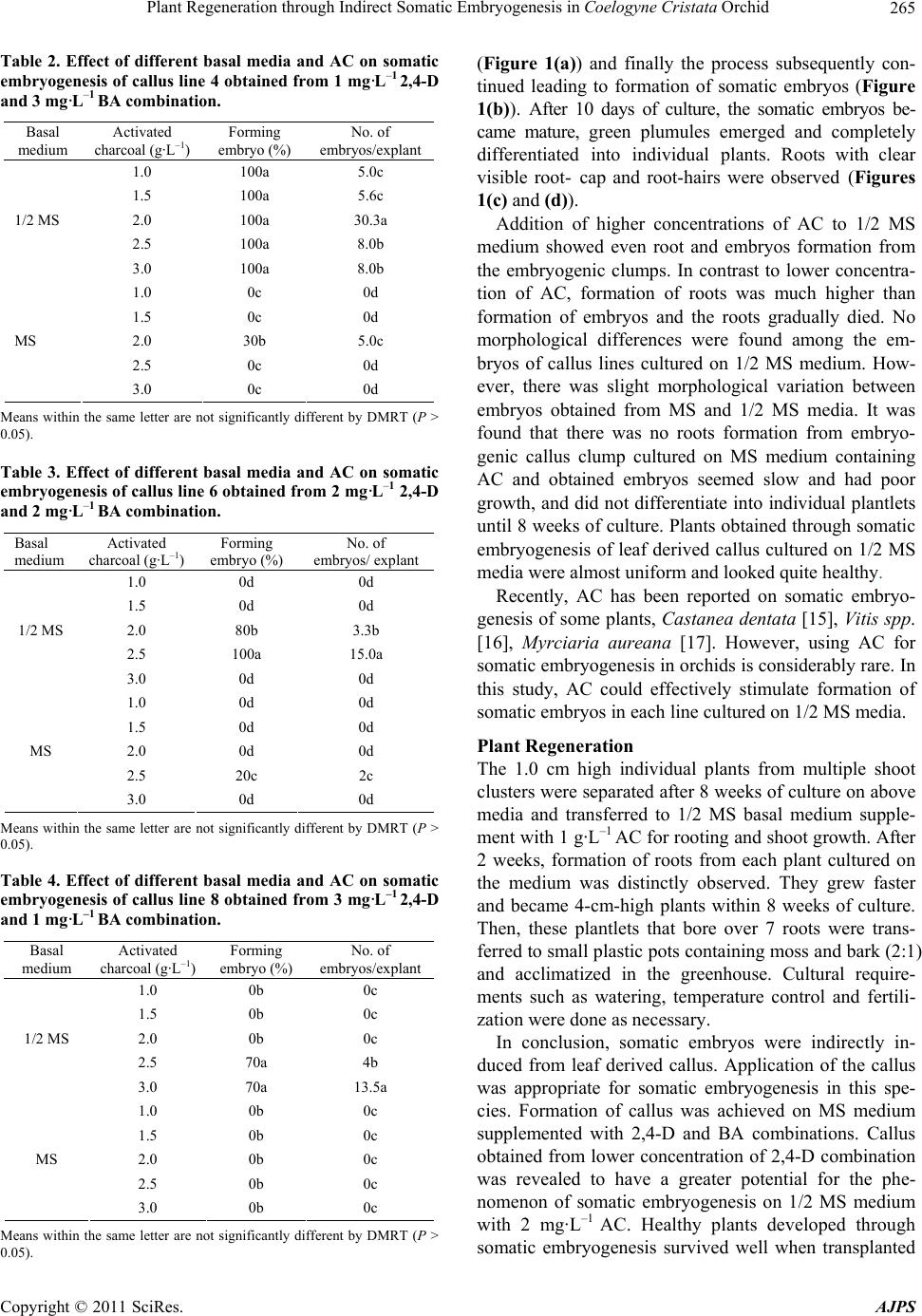
To examine the effect of media and AC on somatic em-
bryogenesis, the above leaves derived callus lines [line
No. 4 (26.7%), line No. 6 (40.0%), line No. 8 (36.7%)]
without CW, which showed better callus formation and
originated from the same tissue but cultured on the dif-
ferent media compositions, were chosen. And, each cal-
lus line was dissected into 2 - 3 mm sized callus pieces
and cultured on the different kinds of basal media (MS
and 1/2 MS) containing (1 - 3 g·L–1) activated charcoal
(AC). Each experiment consisted of 10 explants with
three replications. The culture vessels were placed at
room temperature under 16/8 hr day/night photoperiod at
a 20 - 50 μmolm–²s–¹ photon flux density. Data were
recorded after 8 weeks of culture.
2.3. Plant Regeneration
Single plants derived from the above experiments were
transferred to half-Murashige and Skoog 1/2 MS basal
medium supplemented with 1 g·L–1 AC for plant regen-
eration. Regenerated plants were transplanted and accli-
matized in the greenhouse.
For all cultures, percentage of sucrose and agar was
3.0 and 0.8% respectively. The pH of medium was ad-
justed to 5.8 before adding agar. Data were analyzed by
DMRT (P > 0.05).
3. Results and Discussion
Developing somatic embryogenic culture systems with
reliable regeneration capacity from ornamental plants is a
prerequisite for mass propagation and their genetic im-
provement. Studies were being intensively made to de-
velop a protocol for somatic embryogenesis. The process
can be induced in tissue cultures of orchids either directly
from the epidermal cells of explants [1-3] or indirectly via
intervening callus [4 ]. In this study, somatic embryo-
genesis was induced indirectly through callus culture.
In general, formation of callus in orchids is rather dif-
ficult due to its slow growth and necrotic tendency, espe-
cially, success of callus induction from leaf segments is
the rarest. In this study, when leaf segments were cul-
tured on the hormone free medium they became swollen,
and followed by initiation of a callus mass was visible
from the wound edges of leaf segment after 4 weeks of
culture. The calli were granular, whitish to light yellow
and comprised of isodiametric cells. Higher frequencies
of callus induction from explants were obtained on the
basal media supplemented with various concentrations of
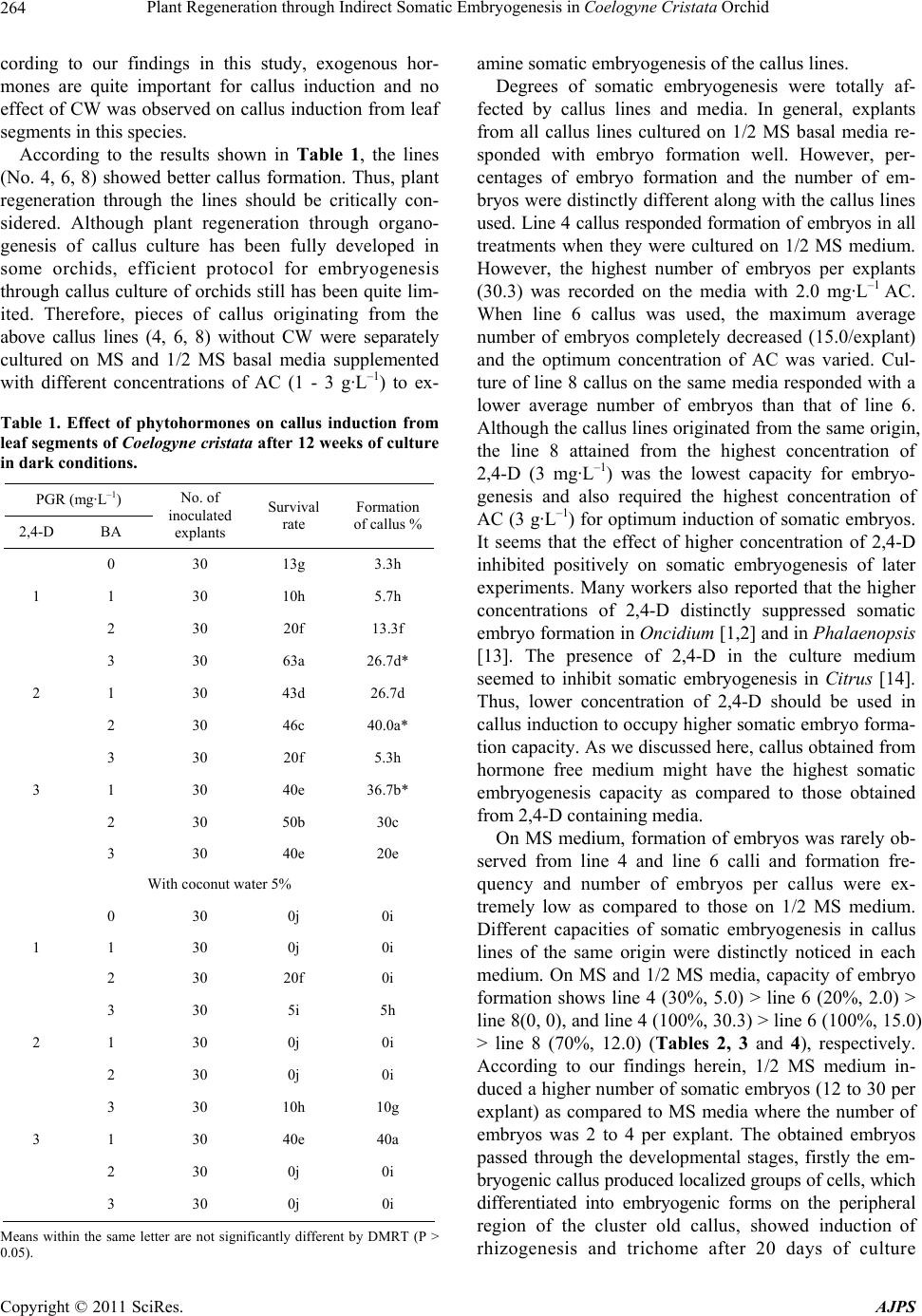
2,4-D and BA (Table 1). However, ratios between con-
centrations of 2,4-D and BA were significantly associ-
ated with percentages of survival of explants and callus
formation. On the contrary, either the lowest concentra-
tion of 2,4-D and the highest concentration of BA or a
higher concentration of 2,4-D and lower concentration of
BA showed synergistic effects on callus induction from
leaf segments. Of the combinations of 2,4-D and BA
tested, 2 mg·L–1 2,4-D and 2 mg·L–1 BA were found as
optimal concentrations for the best callus induction
(40.0%). The color of callus induced from each combina-
tion was slightly changed from whitish to yellow and
most yellow callus showed granular type. However, cal-
lus mass obtained from highest concentrations of 3
mg·L–1 2,4-D was reddish yellow in color.
Addition of CW to the same media containing 2,4-D
and BA, frequencies of callusing were relatively low as
compared to those on the media devoid of CW. Almost
all of the leaf segments died on CW containing media.
However, percentage of callusing was stable on the me-
dia containing concentration of 3 mg·L–1 2,4-D and 1
mg·L–1 BA as devoid of CW.
Recently, combinations of 2,4-D and TDZ have been
reported for the callus induction of ornamental plants
including some orchid genera, Cymbidium [7], Vanda
coerulea [9], Cypripedium formosanum [10]. However,
BA alone or in combination with 2,4-D totally inhibited
callus induction in Paphiopedilum hybrid [11]. Similarly,
[12] mentioned that the combination of 2,4-D and BA
could not effectively induce callus from leaf segment in
Phalaenopsis. In the present study, combination of 2,4-D
and BA has successfully induced callus from leaf seg-
ments within a short period of time. However, the pres-
ence of CW inhibited callus induction from leaf segment
in this species. A similar result was observed in Pha-
laenopsis [12]. It is clear that CW has an inhibitory effect
on callus induction from leaf segments of orchids. Ac-
Copyright © 2011 SciRes. AJPS