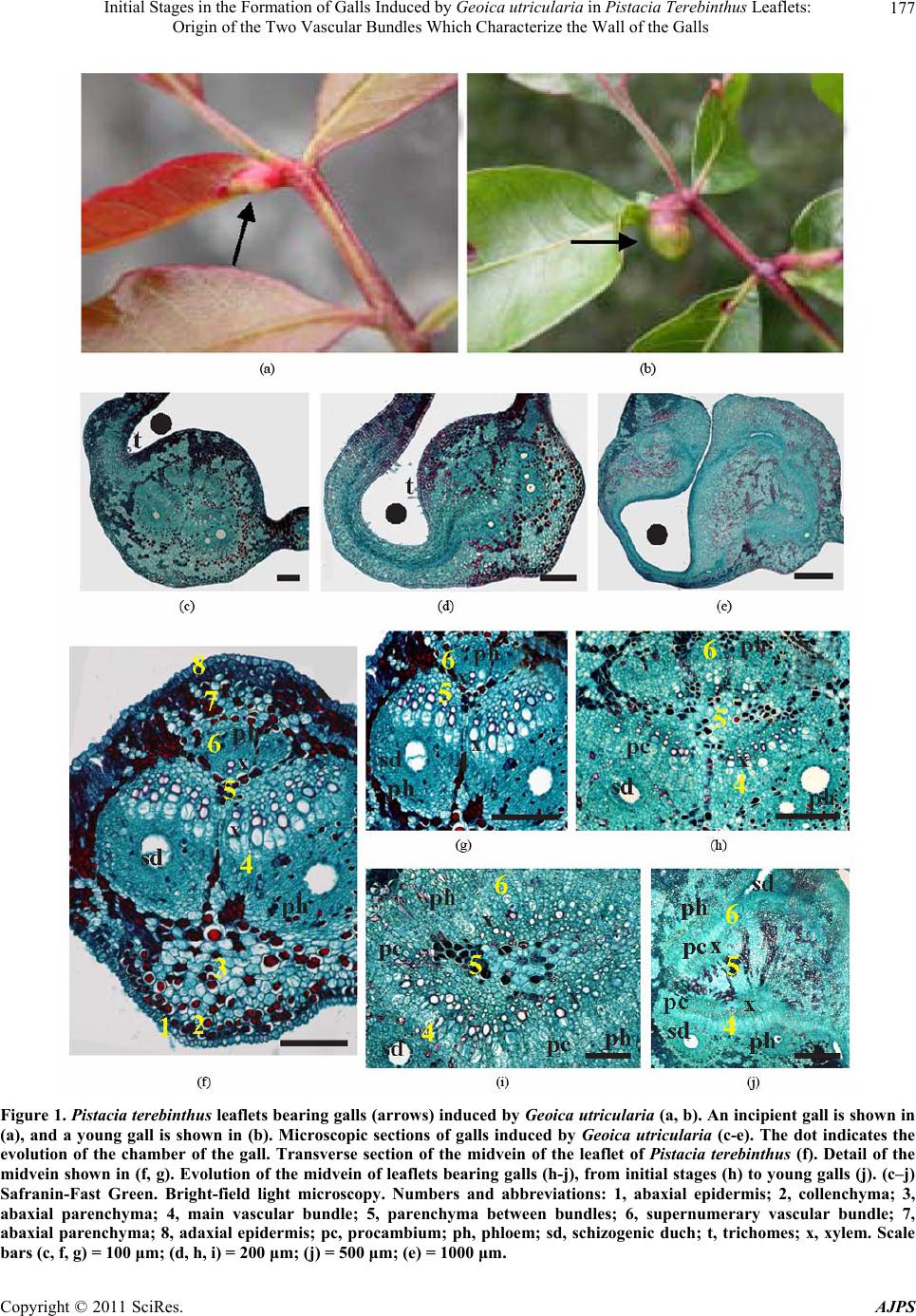
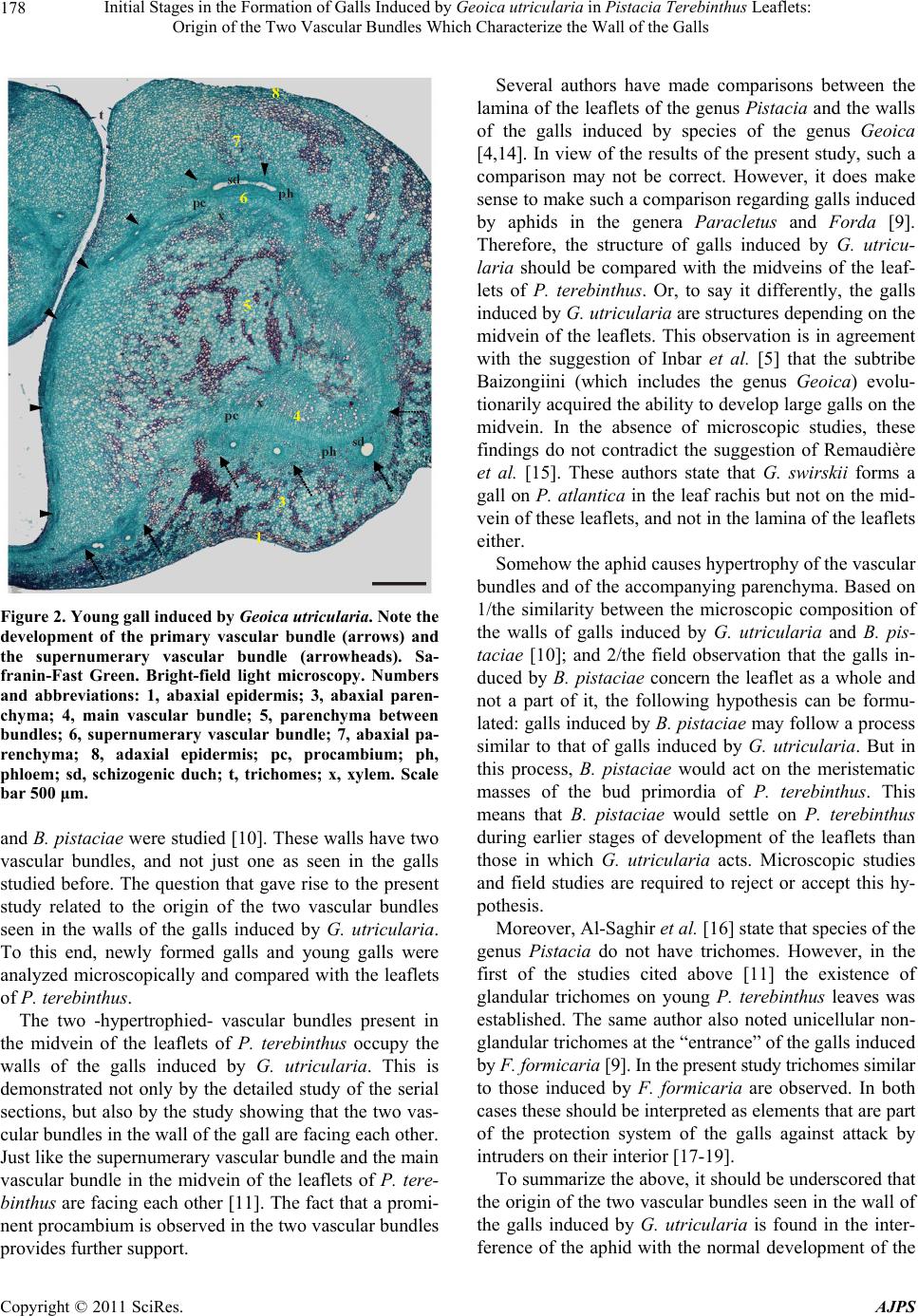
Initial Stages in the Formation of Galls Induced by Geoica utricularia in Pistacia Terebinthus Leaflets: 179
Origin of the Two Vascular Bundles Which Characterize the Wall of the Galls
midvein of the leaflets, especially of the two vascular
bundles and the surrounding parenchyma cells.
5. Acknowledgements
The author wishes to thank to Nicolas Pérez Hidalgo,
Antonio Enzina García, Adoración Candelas González
and Juan Nieto Nafría for revising the manuscript and
Ronald Hartong of TECcientífica for his linguistic assis-
tance. I also thank the Junta de Castilla y León for fund-
ing project LE006A09.
REFERENCES
[1] N. J. Spiller, F. M. Kimmins and M. Llewellyn, “Fine
Structure of Aphid Stylet Pathways and Its Use in Host
Plant Resistance Studies,” Entomologia Experimentalis et
Applicata, Vol. 38, No. 3, 1985, pp. 293-295.
doi:10.1111/j.1570-7458.1985.tb03534.x
[2] W. F. Tjallingh and T. Hogen-Esch, “Fine Structure of
Aphid Stylet Routes in Plant Tissues in Correlation with
EPG Signals,” Physiological Entomology, Vol. 18, No. 3,
1993, pp. 317-328.
doi:10.1111/j.1365-3032.1993.tb00604.x
[3] M. Inbar, “The Evolution of Gall Traits in the Fordinae,”
In: K. Ozaki, J. Yukawa, T. Ohgushi and P. E. Price, Eds.,
Ecology and Evolution of Galling Arthropods and Their
Associates, Springer-Verlag, Tokyo, 2006, pp. 265-273.
doi:10.1007/4-431-32185-3_23
[4] D. Wool, R. Aloni, O. Ben-Zvi and M. Wollberg, “A
Galling Aphid Furnishes Its Home with a Built-in Pipe-
line to the Host Food Supply,” Entomologia Experimen-
talis et Applicata, Vol. 91, No. 1, 1999, pp. 183-186.
doi:10.1046/j.1570-7458.1999.00482.x
[5] M. Inbar, M. Wink and D. Wool, “The Evolution of Host
Plant Manipulation by Insects: Molecular and Ecological
Evidence from Gall-Forming Aphids on Pistacia,” Mo-
lecular Phylogenetics and Evolution, Vol. 32, No. 2, 2004,
pp. 504-511. doi:10.1016/j.ympev.2004.01.006
[6] J. Meyer, “Plant Galls and Gall Inducers,” Gebrüder
Borntraeger, Berlin, 1987.
[7] M. S. Mani, “Ecology of Plant Galls,” Dr. Junk Publisher,
The Hague, 1964.
[8] M. Arduin and J. E. Kraus, “Anatomia e Ontogenia de
Galhas Foliares de Piptadenia Gonoacantha (Fabales,
Mimosaceae) ,” Boletim de Botânica da Universidade de
São Paulo, Vol. 14, 1995, pp. 109-130.
[9] R. Álvarez, A. Encina and N. P. Hidalgo, “Histological
Aspects of Three Pistacia Terebinthus Galls Induced by
Three Different Aphids: Paracletus Cimiciformis, Forda
Marginata and Forda Formicaria,” Plant Science, Vol.
176, No. 2, 2009, pp. 303-314.
doi:10.1016/j.plantsci.2008.11.006
[10] R. Álvarez, “Microscopic Study of the Walls of Galls
Induced by Geoica utricularia and Baizongia Pistaciae in
Pistacia Terebinthus: A Contribution to the Phylogeny of
Fordini,” Arthropod-Plant Interactions, 2011, submitted
for publication.
[11] R. Álvarez, A. Encina and N. P. Hidalgo, “Pistacia Tere-
binthus L. Leaflets: An Anatomical Study,” Plant Sys-
tematic Evolution, Vol. 272, No. 1-4, 2008, pp. 107-118.
doi:10.1007/s00606-007-0640-0
[12] B. Ortiz-Rivas, D. Martínez-Torres and N. Pérez-Hidalgo,
“Molecular Phylogeny of Iberian Fordini (Aphididae:
Eriosomatinae): Implications for the Taxonomy of Genera
Forda and Paracletus,” Systematic Entomology, Vol. 34,
No. 2, 2009, pp. 293-306.
doi:10.1111/j.1365-3113.2008.00464.x
[13] M. Martínez-Millán and S. R. S. Cevallos-Ferriz, “Ar-
quitectura Foliar de Anacardiaceae. Leaf Architecture of
Anacardiaceae,” Revista Mexicana de Biodiversidad, Vol.
76, 2005, pp. 137-190.
[14] D. Wool and N. Bar-El, “Population Ecology of the Gall-
ing Aphid Forda Formicaria von Heyden in Israel:
Abundance, Demography, and Gall Structure,” Israel
Journal of Zoology, Vol. 41, 1995, pp. 175-192.
[15] G. Remaudière, M. Inbar, J. Menier and A. Sumida, “Un
Nouveau Geoica Gallicole sur Pistacia atlantica en Jor-
danie Hemiptera, Aphididae, Eriosomatinae, Fordini,”
Revue Française d’Entomologie, Vol. 26, No. 1, 2004, pp.
37-42.
[16] M. G. Al-Saghir, D. M. Porter and E. T. Nilsen, “Leaf
Anatomy of Pistacia Species (Anacardiaceae),” Journal
of Biological Sciences, Vol. 6, No. 2, 2006, pp. 242-244.
doi:10.3923/jbs.2006.242.244
[17] J. Meyer and J. Maresquelle, “Anatomie des Galles,”
Ggebrüder Borntrager, Berlin, 1983.
[18] A. T. Simmons, G. M. Gurr, D. McGrath, H. I. Nicol and
P. M. Martin, “Trichomes of Lycopersicon spp. and Their
Effect on Myzus Persicae (Sulzer) (Hemiptera: Aphidi-
dae),” Australian Journal of Entomology, Vol. 42, 2003,
pp. 373-378. doi:10.1046/j.1440-6055.2003.00376.x
[19] M. Arduin, G. W. Fernandes and J. E. Kraus, “Morpho-
genesis of Galls Induced by Baccharopelma Dracunculi-
foliae (Hemiptera: Psyllidae) on Baccharis Dracunculifo-
lia (Asteraceae) Leaves,” Brazilian Journal of Biology,
Vol. 65, No. 4, 2005, pp. 559-571.
doi:10.1590/S1519-69842005000400002
Copyright © 2011 SciRes. AJPS