 Chinese Medicine, 2011, 2, 62-70 doi:10.4236/cm.2011.22012 Published Online June 2011 (htt p://www.SciRP.org/journa l/cm) Copyright © 2011 SciRes. CM Pharmacological Modulation by Shakuyakukanzoto (Shao-Yao-Gan-Cao-Tang) and the Ingredients in Rat Intestinal Smooth Muscle Hiroyasu Satoh, Kiminori Tsuro Departm e nt of P harmacol ogy, Traditional Herbal Medicine, School of Medi c ine, Nara Medical University, Kashihara, Japan E-mail: hysat@naramed-u.ac.jp Received January 21, 2011; revised March 10, 2011; accepted May 24, 2011 Abstract Shakuyakukanzoto (Shao-Yao-Gan-Cao-Tang), a formulation of Japanese herbal (Kampo) medicines, is composed of Paeoniae Radix and Glycyrrhizae Radix. Effects of Shakuyakukanzoto and the ingredients on rat intestinal tract were ex amined. Shakuyakuk anzoto (0.01 - 0.3 mg/ml) relaxed a carbachol (CCh, 0.3 μM) -induced contraction in a concentration-dependent manner. Both components (Paeoniae Radix and Glycyr- rhizae Radix) also relaxed the CCh-induced contraction. At 0.1 to 1 mM, their constituents (paeoniflorin and glycyrrhetic acid) and the metabolic products (18-α- and 18-β-glycyrrhetinic acids) exerted almost the same actions. The relaxations induced by Shakuyakukanzoto were not modified by 1 μM nicardipine, 10 μM su- ramin (ATP receptor inhibitor) and several K+ channel inhibitors, but was attenuated by 20 μM IBMX (a phosphodiesterase inhibitor). Also, IBMX inhibited the relaxations induced by paeoniflorin and glycyrrhetic acid, but not by other ingredients. Nicardipine decreased the relaxation of just 18-α-glycyrrhetinic acid. Even in non-treatment with CCh, Shakuyakukanzoto relaxed the intestinal tract. CCh (0.3 μM) elicited spontane- ous contractions in 23% specimens, depressed by application of Shakuyakukanzoto. These results indicate that Shakuyakukanzoto causes a remarkable relaxation by the anti-cholinergic and the PDE inhibitory actions, but by minor contribution of Ca2+ channel inhibition. Thus, Shakuyakukanzoto exerts an anti-spasmodic ac- tion due to the interaction with pharmacological effects of its ingredients. Keywords: Shakuyakukanzoto, Paeoniae Radix, Paeoniflorin, Glycyrrhetic Acid, PDE Inhibition, Anti-Cholinergic Action, Ca2+ Channel Inhibition, Intestinal Tract 1. Introduction Since traditional Japanese herbal (Kampo) medicines are composed of a mixture with lots of herbs, they produce multiple pharmacological and physiological functions. Shakuyakukanzoto (Shao-Yao-Gan-Cao-Tang), a kind of Kampo formulations, is composed of just two compo- nents; Paeoniae Radix and Glycyrrhizae Radix. The main ingredient of Paeoniae Radix is paeoniflorin, and that of Glycyrrhizae Radix is glycyrrhetic acid. Shakuyakukanzoto has been mostly used for the re- laxant effect of skeletal muscle [1]. Nicotinic ACh re- ceptors on neuromuscular junction play an important role for the contraction. Paeoniflorin produced the relaxation by means of a depolarized blockade like succinylcholine [2]. Paeoniflorin regulates Ca2+ movement near around neuromuscular junction, and glycyrrhetic acid inhibits Ca2+-activated K+ (IKCa) channel to repolarize or hyper- polarize the membrane [3]. The combination with Paeo- niae radix and Glycyrrhizae radix enforces the relaxant action of skeletal muscle. Also, Shakuyakukanzoto may be useful to relieve a pain, and exhibit an anti-spasmodic action in gastrointes- tinal smooth muscle [4]. The relaxations of smooth mus- cles induced by Kampo medicines depend on mainly a phosphodiesterase (PDE) inhibition [5]. Most recent re- ports have also demonstrated to play a key role for regu- lation of the gap junction on gastrointestinal smooth muscle [6-8]. Until now, there is less information of more detailed pharmacological mechanisms for the gastrointestinal ac- tions of Shakuyakukanzoto, the components (Paeoniae  H. SATOH ET AL. Copyright © 2011 SciRes. CM Radix and Glycyrrhizae Radix), especially the ingredients (paeoniflorin and glycyrrhetic acid) and the metabolic products (18-β- and 18-α-glycyrrhetinic acids). In the present experiments, therefore, the pharmacological ac- tions of Shakuyakukanzoto and the ingredients on the re- laxation were investigated using rat intestinal smooth muscle. 2. Material and Methods All experiments were carried out, according to the guide- lines laid down by the Nara Medical University Animal Welfare Committee, and also under the terms of the Declaration of Helsinki. 2.1. Experimental Procedures Wistar rats (8 to 15 weeks-old), weighing approximately 300 g, were anesthetized with ether, and euthanized by exsanguination. The intestinal tract was quickly removed, and the isolated intestinal tract was cut into rings of 1.5 cm in length. The strips were suspended in a jacketed organ chamber fill ed wit h 20 m l m odifi ed Ty rode s olution. The strips were suspended between both sides with stainless st e el st irrups. The lower s tirrup was anchore d a nd the upper stirrup was attached to a force-displacement transducer (Nihon Kohden TB-652T, Tokyo, Japan) to record the isometric force. All strips were stretched to generate a resting tension of 1.0 g, which was optimal fo r contractions with muscarin ic ACh receptor agon ist. After 40 min of resting, carbachol (CCh, 0.3 μM) was added to the tissue bath. After the contractile response became steady, the drugs were cumulatively administrated into the bath solution. The effects of each con centration of th e drugs were measured 5 - 7 min after the responses beca me steady. To examine the involvement with Ca2+ channel, PDE or other mechanisms, the pretreatment with 1 μM nicardipine, 20 μM IBMX or oth er inhibitors was carried out. Each of the experiments was examined at least qua- druplicates. The responses were analyzed as a percentage change from the value before an application of drugs. 2.2. Experiments of Spontaneous Contractions Pretreatment with CCh (0.3 μM) was usually carried out. Under the conditions, the spontaneous contractions were exhibited occasionally in some specimens. The effects of Shakuyakukanzoto on the spontaneous contractions were investigated. 2.3. Experiments in the Absence of CCh Using rat ileum in non-pretreatment with CCh, the ef- fects of Shakuyakukanzoto and the ingredients on ga- strointestinal smooth muscle were examined using the same experimental techniques. 2.4. Solution and Drugs The modified Tyrode solution was comprised of (in mM); 136.8 mM NaCl, 2.7 mM KCl, 1.8 mM CaCl2, 1.1 mM MgCl2, 0.4 mM NaH2PO4, 11.9 mM NaHCO3, and 5.6 mM glucose. The chamber solution was kept at 36.5˚C and oxygenated with 95% O2 and 5% CO2. The drugs used were Shakuyakukanzoto, Paeoniae Radix and Glycyrrhizae Radix (Tsumura Co., Tokyo, Japan), as a spray-dried powder extracted with boiling water of a ground raw materials. Each drug was dis- solved with DMSO. Other drugs used were CCh, 18-α-glycyrrhetinic acid and 18-β-glycyrrhetinic acid (Sigma Chemical, MO. U.S.A.), and paeoniflorin and glycyrrhtic acid (Wako Chemical, Kyoto, Japan). Nicar- dipine (Ca2+ channel inhibitor), Bay K 8644 (Ca2+ chan- nel stimulator), suramin (ATP receptor inhibitor), apa- min (Ca2+-activated K+ channel inhibitor), glibencl amide (ATP-sensitive K+ channel inhibitor), tetraethyammo- nium (TEA, voltage-dependent K+ channel inhibitor) and 3-isobutyl-1-methylxanthine (IBMX, phosphodiesterase inhibitor) (Sigma) were also used. 2.5. Statistical Analyses To compare the pair values, we are used statistical me- thods of the Student’s t-test and ANOVA followed by post-hoc tests (Dunn-Bonferonii test) using Excel (Mi- crosoft Inc., Washington, U.S.A.) and S-PLUS (Mathe- matical System Inc., Washington, U.S.A.). All values are represented as means ± SEM. A p value of less than 0.05 was considered significant. 3. Results 3.1. Effects of Shakuya kukan zoto on CCh-Induced Contraction Pretreatment with 0.3 μM CCh produced a strong con- traction of isolated ileum; by 1.4 ± 0.2 g (n = 157). Then, Shakuyakukanzoto (0.01 to 0.3 mg/ml) was admini- strated cumulatively into the bath, and at over 0.1 mg/ml significantly relaxed the CCh-indu ced contraction; at 0.3 mg/ml by 27.7 ± 3.3% (n = 12, P < 0.001). The res- ponses were concentration-dependent. These results are summarized in Table 1. The relaxation induced by Shakuyakukanzoto (0.3 mg/ml) increased (by 37.7 ± 2.0%, n = 8, P < 0.001) at 1 μM nicardipine, but decreased to 15.1 ± 3.2% (n = 5, P < 0.05) at 20 μM IBMX. Interestingly nicardipine rather 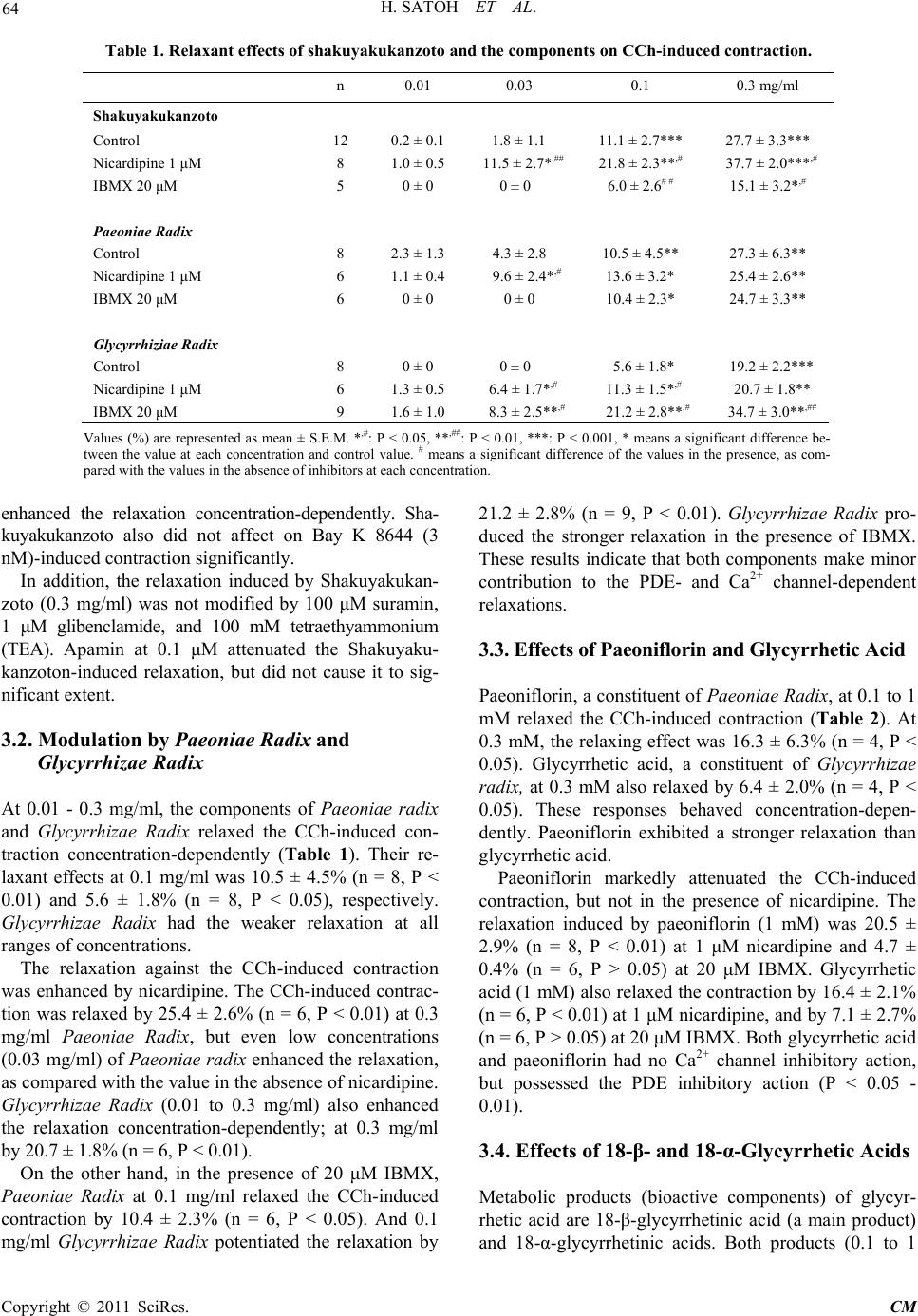 H. SATOH ET AL. Copyright © 2011 SciRes. CM Table 1. Relaxant effects of shakuyakukanzoto and the components on CCh-induced con t raction. n 0.01 0.03 0.1 0.3 mg/ml Shaku yakukanzoto Control 12 0.2 ± 0.1 1.8 ± 1.1 11.1 ± 2.7*** 27.7 ± 3.3*** Nicardipine 1 μM 8 1.0 ± 0.5 11.5 ± 2.7*,## 21.8 ± 2.3**,# 37.7 ± 2.0***,# IBMX 20 μM 5 0 ± 0 0 ± 0 6.0 ± 2.6# # 15.1 ± 3 .2*,# Paeoniae Radix Control 8 2.3 ± 1.3 4.3 ± 2.8 10.5 ± 4.5** 27.3 ± 6.3** Nicardipine 1 μM 6 1.1 ± 0.4 9 .6 ± 2.4*,# 13.6 ± 3. 2 * 25.4 ± 2.6* * IBMX 20 μM 6 0 ± 0 0 ± 0 10.4 ± 2.3* 24.7 ± 3.3** Glycyrrhiziae Radix Control 8 0 ± 0 0 ± 0 5.6 ± 1.8* 19.2 ± 2.2*** Nicardipine 1 μM 6 1.3 ± 0.5 6.4 ± 1.7*,# 11.3 ± 1 .5*,# 20.7 ± 1.8** IBMX 20 μM 9 1.6 ± 1.0 8. 3 ± 2.5**,# 21.2 ± 2.8**,# 34.7 ± 3.0**,## Values (%) are represent ed as mean ± S.E.M. *,#: P < 0.05, **,##: P < 0.01, ***: P < 0.001, * means a significant difference be- tween the value at each concentration and control value. # means a significant difference of the values in the presence, as com- pared with the values in the absence of inhibitors at each concentration. enhanced the relaxation concentration-dependently. Sha- kuyakukanzoto also did not affect on Bay K 8644 (3 nM)-induced contraction significantly. In addition, the relaxation induced by Shakuyakukan- zoto (0.3 mg/ml) was not modified by 100 μM suramin, 1 μM glibenclamide, and 100 mM tetraethyammonium (TEA). Apamin at 0.1 μM attenuated the Shakuyaku- kanzoton-induced relaxation, but did not cause it to sig- nificant extent. 3.2. Modulation by Paeoniae Radix and Glycyrrhizae Radix At 0.01 - 0.3 mg/ml, the components of Paeoniae radix and Glycyrrhizae Radix relaxed the CCh-induced con- traction concentration-dependently (Table 1). Their re- laxant effects at 0.1 mg/ml was 10.5 ± 4.5% (n = 8, P < 0.01) and 5.6 ± 1.8% (n = 8, P < 0.05), respectively. Glycyrrhizae Radix had the weaker relaxation at all ranges of c oncentrat i ons. The relaxation against the CCh-induced contraction was enhanced by nicardipine. The CCh-induced contrac- tion was relaxed by 25.4 ± 2.6% (n = 6, P < 0.01) at 0.3 mg/ml Paeoniae Radix, but even low concentrations (0.03 mg/ml) of Paeoniae radix enhanced the relaxation, as compared with the value in the absence of nicardipine. Glycyrrhizae Radix (0.01 to 0.3 mg/ml) also enhanced the relaxation concentration-dependently; at 0.3 mg/ml by 20.7 ± 1.8% (n = 6, P < 0.01). On the other hand, in the presence of 20 μM IBMX, Paeoniae Radix at 0.1 mg/ml relaxed the CCh-induced contraction by 10.4 ± 2.3% (n = 6, P < 0.05). And 0.1 mg/ml Glycyrrhizae Radix potentiated the relaxation by 21.2 ± 2.8% (n = 9, P < 0.01). Glycyrrhizae Radix pro- duced the stronger relaxation in the presence of IBMX. These results indicate that both components make minor contribution to the PDE- and Ca2+ channel-dependent relaxations. 3.3. Effects of Paeoniflorin and Glycyrrhetic Acid Paeoniflorin, a constituen t of Paeoniae Radix, a t 0.1 to 1 mM relaxed the CCh-induced contraction (Table 2). At 0.3 mM, the relaxing effect was 16.3 ± 6.3% (n = 4, P < 0.05). Glycyrrhetic acid, a constituent of Glycyrrhizae radix, at 0.3 mM also relaxed by 6.4 ± 2.0% (n = 4, P < 0.05). These responses behaved concentration-depen- dently. Paeoniflorin exhibited a stronger relaxation than glycyrrhetic acid. Paeoniflorin markedly attenuated the CCh-induced contraction, but not in the presence of nicardipine. The relaxation induced by paeoniflorin (1 mM) was 20.5 ± 2.9% (n = 8, P < 0.01) at 1 μM nicardipine and 4.7 ± 0.4% (n = 6, P > 0.05) at 20 μM IBMX. Glycyrrhetic acid (1 mM) also relaxed the contraction by 16.4 ± 2.1% (n = 6, P < 0.01) at 1 μM nicardipine, and by 7.1 ± 2.7% (n = 6, P > 0.05) at 20 μM IBMX. Both glycyrrhetic acid and paeoniflorin had no Ca2+ channel inhibitory action, but possessed the PDE inhibitory action (P < 0.05 - 0.01). 3.4. Effects of 18-β- and 18-α-Glycyrrhetic Acids Metabolic products (bioactive components) of glycyr- rhetic acid are 18-β-glycyrrhetinic acid (a main product) and 18-α-glycyrrhetinic acids. Both products (0.1 to 1 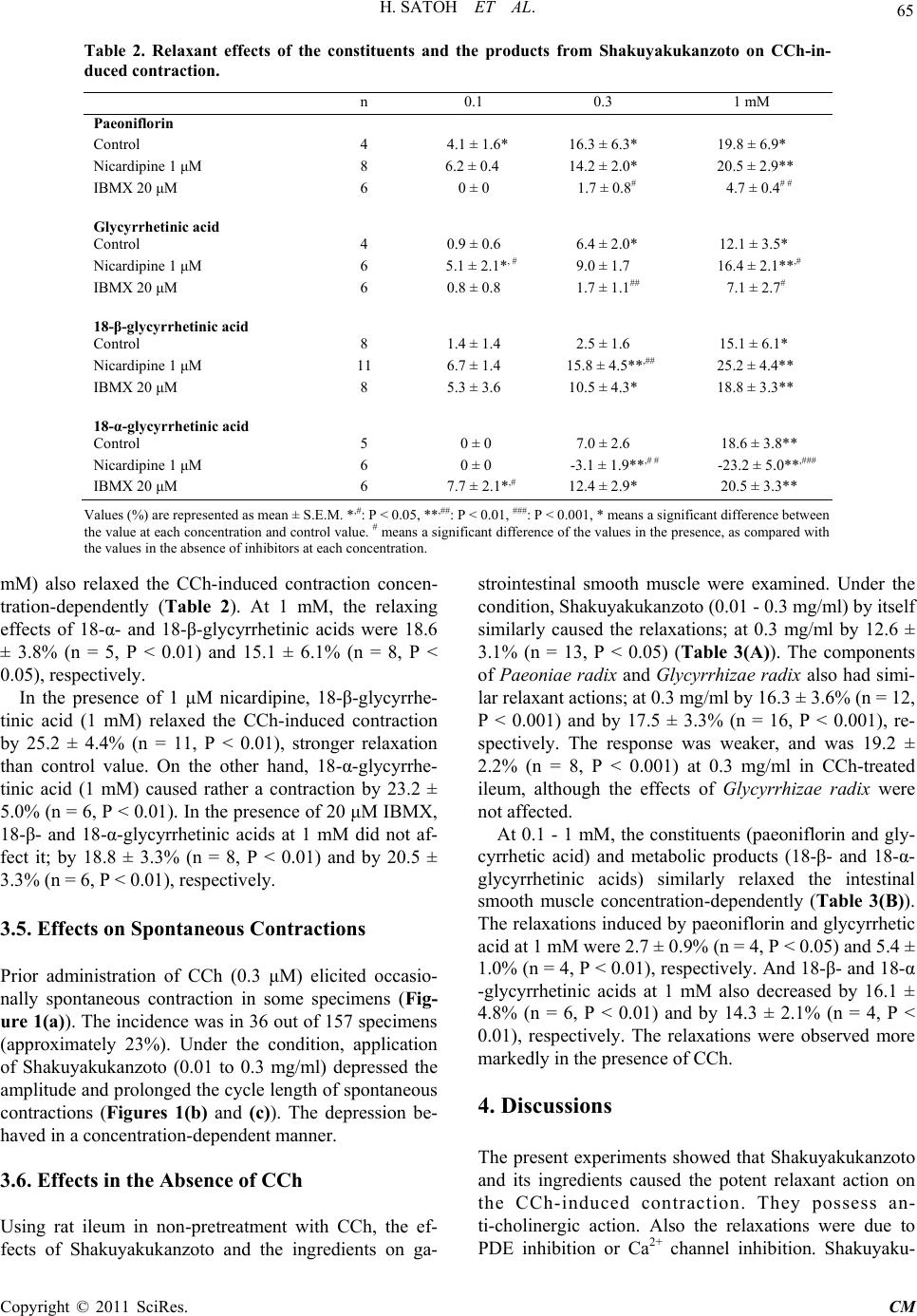 H. SATOH ET AL. Copyright © 2011 SciRes. CM Table 2. Relaxant effects of the constituents and the products from Shakuyakukanzoto on CCh-in- duced contr action. Glycyrrhetinic acid IBMX 20 μM 6 0.8 ± 0.8 1.7 ± 1.1 7.1 ± 2.7 Nicardipine 1 μM 11 6.7 ± 1. 4 15.8 ± 4.5** 25.2 ± 4.4** 18-α-glycyrrhetinic acid Values (%) are represented as mean ± S.E.M. *,#: P < 0.05, **,##: P < 0.01, ###: P < 0.001, * means a significant difference between the valu e at each con centr ation and co ntro l valu e. # means a sig nifi cant d ifferen ce of t he val ues i n the p resen ce, as co mpared with the values in the absence of inhibitors at each concentration. mM) also relaxed the CCh-induced contraction concen- tration-dependently (Table 2). At 1 mM, the relaxing effects of 18-α- and 18-β-glycyrrhetinic acids were 18.6 ± 3.8% (n = 5, P < 0.01) and 15.1 ± 6.1% (n = 8, P < 0.05), respectively. In the presence of 1 μM nicardipine, 18-β-glycyrrhe- tinic acid (1 mM) relaxed the CCh-induced contraction by 25.2 ± 4.4% (n = 11, P < 0.01), stronger relaxation than control value. On the other hand, 18-α-glycyrrhe- tinic acid (1 mM) caused rather a contraction by 23.2 ± 5.0% (n = 6, P < 0.01). In the presence of 20 μM IBMX, 18-β- and 18-α-glycyrrhetinic acids at 1 mM did not af- fect it; by 18.8 ± 3.3% (n = 8, P < 0.01) and by 20.5 ± 3.3% (n = 6, P < 0.01), respectively. 3.5. Effects on Spontaneous Contractions Prior administration of CCh (0.3 μM) elicited occasio- nally spontaneous contraction in some specimens (Fig- ure 1(a)). The incidence was in 36 out of 157 specimens (approximately 23%). Under the condition, application of Shakuyakukanzoto (0.01 to 0.3 mg/ml) depressed the amplitude and prolonged the cycle length of spontaneous contractions (Figures 1(b) and (c)). The depression be- haved in a concentration-dependent manner. 3.6. Effects in th e Absence of CCh Using rat ileum in non-pretreatment with CCh, the ef- fects of Shakuyakukanzoto and the ingredients on ga- strointestinal smooth muscle were examined. Under the condition, Shakuyakukanzoto (0.01 - 0.3 mg/ml) by itself similarly caused the relaxations; at 0.3 mg/ml by 12.6 ± 3.1% (n = 13, P < 0.05) (Table 3(A)). The components of Paeoniae radix and Glycyrrhizae radix also had simi- lar relaxant actions; at 0.3 mg/ml by 16.3 ± 3.6% (n = 12, P < 0.001) and by 17.5 ± 3.3% (n = 16, P < 0.001), re- spectively. The response was weaker, and was 19.2 ± 2.2% (n = 8, P < 0.001) at 0.3 mg/ml in CCh-treated ileum, although the effects of Glycyrrhizae radix were not affected. At 0.1 - 1 mM, the constituents (paeoniflorin and gly- cyrrhetic acid) and metabolic products (18-β- and 18-α- glycyrrhetinic acids) similarly relaxed the intestinal smooth muscle concentration-dependently (Table 3(B)). The relaxations induced by paeoniflorin and glycyrrhetic acid at 1 mM were 2.7 ± 0.9% (n = 4, P < 0.05) and 5.4 ± 1.0% (n = 4, P < 0.01), r espectively. And 18 -β- and 18-α -glycyrrhetinic acids at 1 mM also decreased by 16.1 ± 4.8% (n = 6, P < 0.01) and by 14.3 ± 2.1% (n = 4, P < 0.01), respectively. The relaxations were observed more markedly in the pre s e nce of CCh. 4. Discussions The present experiments showed that Shakuyakukanzoto and its ingredients caused the potent relaxant action on the CCh-induced contraction. They possess an- ti-cholinergic action. Also the relaxations were due to PDE inhibition or Ca2+ channel inhibition. Shakuyaku- 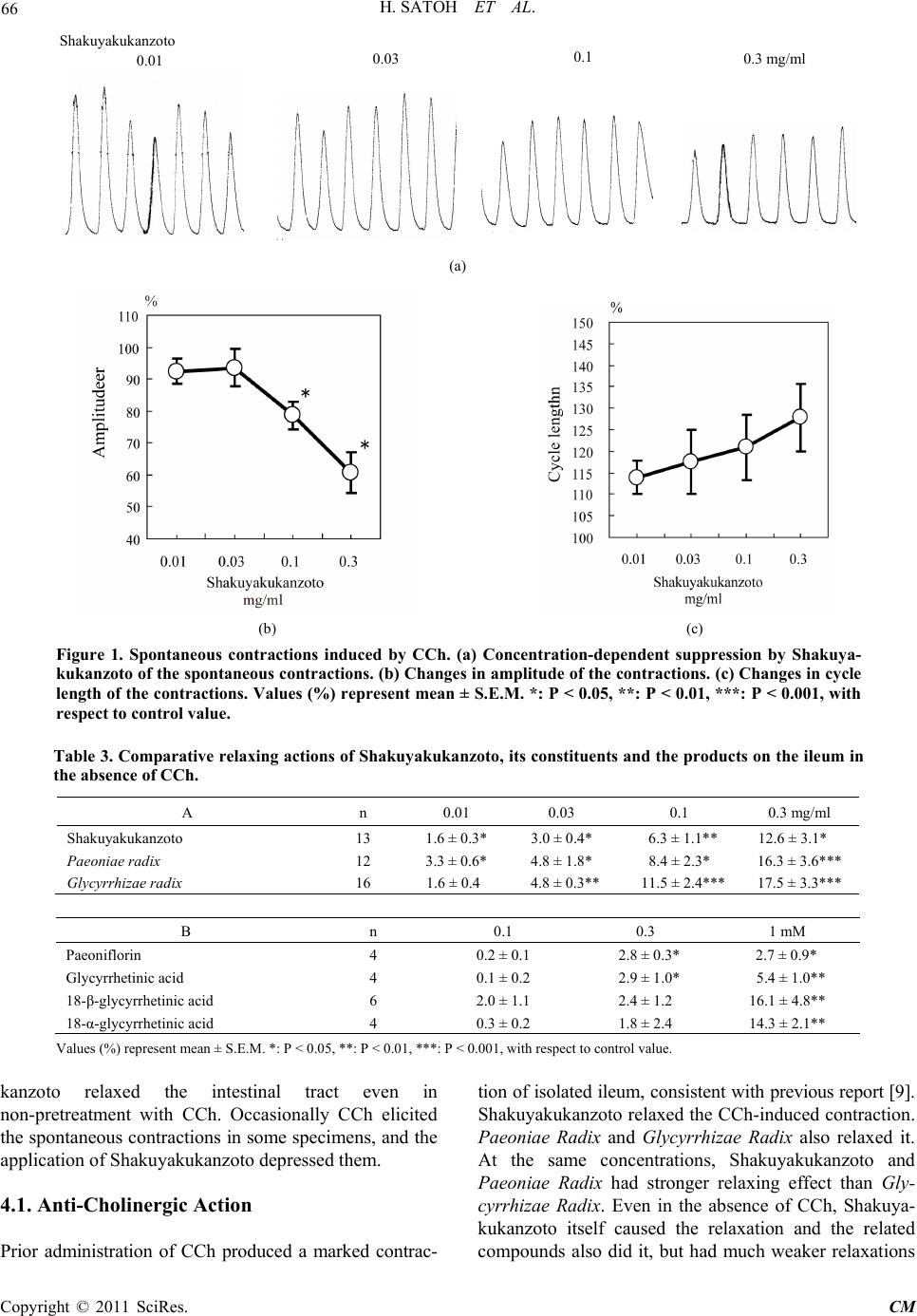 H. SATOH ET AL. Copyright © 2011 SciRes. CM (a) (b) (c) Figure 1. Spontaneous contractions induced by CCh. (a) Concentration-dependent suppression by Shakuya- kukanzoto of the spontaneous contractions. (b) Changes in amplitude of the contractions. (c) Changes in cycle length of the contractions. Values (%) represent mean ± S.E.M. *: P < 0.05, **: P < 0.01, ***: P < 0.001, with respect to control value. Table 3. Comparative relaxing actions of Shakuyakukanzoto, its constituents and the products on the ileum in the absence of CCh. A n 0.01 0.03 0.1 0.3 mg/ml Shakuyakukanzoto 13 1. 6 ± 0.3* 3.0 ± 0. 4 * 6.3 ± 1.1** 12.6 ± 3.1* Paeoniae radix 12 3.3 ± 0.6* 4.8 ± 1.8 * 8.4 ± 2.3* 16.3 ± 3.6*** Glycyrrhizae radix 16 1.6 ± 0.4 4.8 ± 0.3** 11.5 ± 2.4*** 17.5 ± 3.3*** B n 0.1 0.3 1 mM Paeoniflorin 4 0.2 ± 0.1 2.8 ± 0.3* 2.7 ± 0.9* Glycyrrhetinic acid 4 0.1 ± 0.2 2.9 ± 1.0* 5.4 ± 1.0** 18-α-glycyrrhetinic acid 4 0.3 ± 0.2 1.8 ± 2.4 14.3 ± 2.1** Values (%) represent mean ± S.E.M. *: P < 0.05, **: P < 0.01, ***: P < 0.001, with respect to control value. kanzoto relaxed the intestinal tract even in non-pretreatment with CCh. Occasionally CCh elicited the spontaneous contractions in some specimens, and the application of Shakuyakukanzoto depresse d t he m. 4.1. Anti-Cholinergic Action Prior administration of CCh produced a marked contrac- tion of isolated ileu m, consistent with previous report [9]. Shakuyakukanzoto relaxed the CCh-induced contra ction. Paeoniae Radix and Glycyrrhizae Rad ix also relaxed it. At the same concentrations, Shakuyakukanzoto and Paeoniae Radix had stronger relaxing effect than Gly- cyrrhizae Radix. Even in the absence of CCh, Shakuya- kukanzoto itself caused the relaxation and the related compounds also did it, but had much weaker relaxations  H. SATOH ET AL. Copyright © 2011 SciRes. CM as compared with those in presence of CCh. Glycyrrhi- zae Radix makes minor contribution to the CCh-induced relaxation, because it also relaxed the ileum without treatment with CCh to almost the same extent. Paeoniae Radix has been reported to inhibit the con- tractions induced by nicotin e and electric stimulations [3]. The contractions are produced by ACh release from nerve ending mediated through neuroganglionic ACh receptors. Paeoniflorin prevents the damages mediated through muscarinic (M1) receptor in rat hipp ocampal ( CA1) neurons and ameliorates the dysfunctions [10,11]. In the present experiments, both paeoniflorin and glycyrrhetic acid relaxed the CCh-induced contraction. Paeoniflorin had stronger relaxation. 18-α- and 18-β-glycyrrhetinic acids also had the concentration-dependent relaxation. These findings indicate that Shakuyakukanzoto and its containing compounds exhibit the anti-cholinergic action, presumably mediated through M1 (or M3) receptor. Espe- cially Paeoniae Radix and paeoniflorin in Shakuyakukan- zoto possess the s tronge r ant i-cholinergic action. 4.2. Ca2+ Channel Inhibition The inhibition of Ca2+ channel has previously been shown. Paeoniflorin inhibits Ca2+ current in NG108-15 neuronal ce lls, and Na+ current in hippocampus neurons [12], presumably leading to the relaxation of intestinal tract. The inhibition of Na+ current results in a decline of cellular Ca2+ concentration ([Ca2+]i) via Na/Ca ex- change. Paeoniflorin also relaxes the isolated rat aorta due to the [Ca2+]i decline and the increases in NO and cGMP [13]. In the present experiments, however, Shakuyakukan- zoto and the containing compounds had no Ca2+ channel inhibitory action. This is supported by the minor action of Shakuyakukanzoto on Bay K 8644 (3 nM)-induced con- traction. Interestingly, they d ecreased the relaxant actions by addition of nicardipine. Only 18-α-glycyrrhetinic acid had the inhibitory action of Ca2+ channel, and produced rather the contraction. The mechanisms are now unclear yet, but cellular signaling pathways such as Rho kinase, MLCK and P K-C might be involved [14-17]. Therefore, Shakuyakukanzoto and the related com- pounds have less or no effect on Ca2+ channel, but might exert the relaxation by the inhibitions of Na+ and KCa channels and the activation of cGMP signaling pathway. 4.3. PDE Inhibition Shakuyakukanzoto and the ingredients caused the strong PDE inhibitory action. Especially, Glycyrrhizae Radix, paeoniflorin and glycyrrhetic acid were marked. The PDE inhibition (as a result cAMP accumulation) can produce the potent relaxation of smooth muscle. Most Kampo formulations have been reported to exert the PDE inhibitory action [5]. The PDE inhibition was ob- served in paeoniflorin but not in Paeoniae Radix. So, the resultant effects are responsible for an interaction among the containi ng compounds. 4.4. Depression of the Spontaneous Contractions and Spasms The smooth muscle cells may exhibit the spontaneous contractions. The foundation of pacemaker mechanisms is very similar to that of sino-atrial (SA) nodal cells of heart [18]. It is a pendulum movement with a repetitive depolarization and repolarization. In rat pregnant uterus smooth muscle cells with spontaneous contractions, a hyperpolarization-activated inward (or pacemaker) cur- rent (If), is similarly identified [19]. In general, the spon- taneous activity of smooth muscle cells may be largely dependent on transient Ca2+ sparks. Interstitial cells of Cajal, gastrointestinal pacemakers, exhibit Ca2+ release from IP3-dependent Ca2+ stores by activating a Ca2+- dependent cationic current that drives pacemaker depola- rization [20]. The [Ca2+]i elevation activates the KCa channels to produce the repolarization of spontaneous action potentials [21], as well as the Ca2+-activated Cl- channels to produce the depolarization during pacemaker potential [22]. Most recently in guinea pig SA nodal cells, however, we have been found minor contribution of transient Ca2+ sparks [23]. It is not yet clear now, but might be closely related with the connection of Ca2+ channels on the plasma membrane and the sarcoplasmic reticulum (SR) in interstitial smooth muscle. In this study, after administration of CCh, spontaneous contractions occurred with approximately 23% incidence. The KCa channel in aortic smooth muscle cells is inhi- bited by β-adrenoceptor and muscarinic receptor stimula- tions and also by PK-C stimulation [24]. Glycyrrhetic acid also inhibits KCa channel [3]. In this study, applica- tion of Shakuyakukanzoto ceased the spontaneous con- tractions. Thus, the depression would be partly due to inhibition of KCa channel and regulation of Ca2+ move- ment near around neuromuscular junction, because of minor contribution of Ca2+ channel. In skeletal muscles, nicotinic ACh receptor on neuro- muscular junction plays an important role for the con- traction. Paeoniflorin produces the relaxation by means of a depolarized blockade like succinylcholine [2]. Paeo- niflorin regulates Ca2+ movement near around neuro- muscular junction, and glycyrrhetic acid inhibits KCa channel [3]. In rat hippocampal slice, furthermore, the dependence of anti-cholinergic action has been found [10]. The combination with Paeoniae Radix and Glycyr- rhizae Radix would enforce the relaxant action of skelet- al muscle. Most recent reports have demonstrated that  H. SATOH ET AL. Copyright © 2011 SciRes. CM Table 4. Pharmacological actions of Shakuyakukanzoto and the ingredients. Anti-cholinergic action Ca2+ channel inhibition PDE inhibition Shakuyakukanzoto + – + Paeoniae radix + – – Glycyrrhiziae radix – – – Paeoniflorin + – + Glycyrrhetinic acid + – + 18-β-glycyrrhetinic acid + – – 18-α-glycyrrhetinic acid + + – Shakuyakukanzoto may be sufficiently effective for spasmodic diseases of gastrointestinal tract, as well as for cramp and twitch of skeletal muscle. The anti- spasmodic and muscle relaxing effects are exerted not only by the anti-cholinergic action (mediated through nicotinic and muscarinic ACh receptors) but also by PDE inhibitory actions. Glycyrrhetic acid and the me- tabolic products can slow myocardial conduction via modulation of gap junction, but no t via that o f the ion ic channels [25]. Also 18-β- and 18-α-glycyrrhetinic acids exert a blocking action of gap junction [6,8]. The blockade of gap junction prevents spasmodic diseases in gastrointestinal smooth muscle, and reduces epilep- togenicity and arrhythmogenesis [7]. Thus, the phar- macokinetic properties may be well suitable for the transient clamp in leg skeletal muscle and the gastroin- testinal spasms. 5. Conclusions Shakuyakukanzoto possesses the higher bioactivities for rat ileum, and is so effective for many diseases in clinical uses. Shakuyakukanzoto produced a remarkable relaxa- tion by 1) the anti-cholinergic and 2) the PDE inhibitory actions, but by 3) minor contribution of Ca2+ channel inhibition. Also, Shakuyakukanzoto exerted an anti- spasmodic action due to the interaction with pharmaco- logical effects of its ingredients. The absorption is rapid from intestine, and the plasma concentration-time curves are fitted with a mean terminal half-life (T1/2) of 116.2 min [26]. Glycyrrhetic and 18-β- glycyrrhetinic acids possess a free radical scaven ging property [27,28], and anti-allergic activi ti es su ch as p ass iv e cutaneous anaphylaxis and skin contact inflammation [29,30]. However, the pathological findings of the meta- bolic products from glycyrrhetic acid have also been well known [31]. Finally, the pharmacological characteristics of Sha- kuyakukanzoto, the ingredients and the related com- pounds are summarized on Table 4. Further extensive studies are needed to elucidate in more detail mechan- is ms. 6. Acknowledgements The authors wish to express thanks for the supply of Shakuyakukanzoto extract (Tsumura Co.). 7. References [1] M. Kimura, I. Kimura, K. Takahashi, M. Muroi, M. Yo- shizaki, M. Kanaoka and I. Kitagawa, “Blocking Effects of Blended Paeoniflorin or Its Related Compounds with Glycyrrhizin on Neuromuscular Junctions in Frog and Mouse,” Japanese Journal of Pharmacology, Vol. 36, No. 3, 1984, pp. 275-282. doi:10.1254/jjp.36.275 [2] M. Kimura, I. Kimura and H. Nojima, “Depolarizing Neuromuscular Blocking Action Induced by Electro- pharmacological Coupling in the Combined Effect of Paeoniflorin and Glycyrrhizin,” Japanese Journal of Pharmacology, Vol. 37, No. 4, April 1985, pp. 395-399. doi:10.1254/jjp.37.395 [3] M. Kimura, I. Ki mura and M. Kimura, “Dec reasing Effects by Glycyrrhizin and Paeoniflorin on Intracellular Ca2+- Ae- quorin Luminescence Transients with or without Caffeine in Directly Stimulated-Diaphragm Muscle of Mouse,” Japa- nese Journal of Pharmacology, Vol. 39, No. 3, November 1985, pp. 387-390. doi:10.1254/jjp.39.387 [4] J. X. He, E. Goto, T. Akao and T. Tani, “Interaction be- tween Shaoyao-Gancao-Tang and a Laxative with Respect to Alteration of Paeoniflorin Metabolism by Intestinal Bac- teria in Rats,” Phytomedicine, Vol. 14, No. 7-8, August 2007, pp. 454-459. doi:10.1016/j.phymed.2006.09.014 [5] Y. Saegusa, A. Sugiyama, A. Takahara, Y. Nagasawa and K. Hashimoto, “Relationship between Phoshpodiesterase Inhibition Induced by Several Kampo Medicines and Smooth Muscle Relaxation of Gastrointestinal Tract Tis- sues of Rats,” Journal of Pharmacology, Vol. 93, No. 1, 2003, pp. 62-68. [6] Y. Takeda, S. M. Ward, K. M. Sanders and S. D. Koh, “Effects of the Gap Junction Blocker Glycyrrhetinic Acid on Gastrointestinal Smooth Muscle Cells,” American Journal of Physiology, Gastrointestinal Liver Physiology, Vol. 289, No.4, April 2005, pp. G832-G841. doi:10.1152/ajpgi.00389.2004 [7] Z. Gajda, E. Hermesz, E. Gyengesi, Z. Szupera and M. Szente, “The Functional Significance of Gap Junction Channels in the Epileptogenicity and Seizure Susceptibil- ity of Juvenile Rats,” Epilepsia, Vol. 47, No. 6, 2006, pp. 1009-1022. doi:10.1111/j.1528-1167.2006.00573.x  H. SATOH ET AL. Copyright © 2011 SciRes. CM [8] E. E. Daniel, A. F. EI-Yazbi, M. Mannarino, G. Galante, G. Boddy, J. Livergant and T. E. Oskouei, “Do Gap Junc- tions Play a Role in Nerve Transmissions as well a s Pac- ing in Mouse Intestine,” American Journal of Physiology, Gastrointestinal Liver Physiology, Vol. 292, No. 3, 2007, pp. G734-G745. doi:10.1152/ajpgi.00428.2006 [9] A. Sugaya, H. Okita, Y. Takazu and E. Sugaya, “Neru- pharmacological Study of Saiko-keisi-to,” Shoyaku, Vol. 29, 1975, pp. 160-165. [10] K. Tabata, K. Matsumoto and H. Watanabe, “Paeoniflorin, a Major Constituent of Peony Root, Reverses Muscarinic M1-Receptor Antagonist-Induced Suppression of Long- Term Potentiation in the Rat Hipp ocampal Slice ,” Japanese Journal of Pharmacology, Vol. 83, 2000, pp. 25-30. doi:10.1254/jjp.83.25 [11] J. Liu, D. Z. Jin, L. Xiao and X. Z. Zhu, “Paeoniflorin Attenuates Chronic Cerebral Hypoperfusion-Induced Learning Dysfunction and Brain Damage in Rat,” Brain Research, Vol. 1089, No. 1, May 2006, pp. 162-170. doi:10.1016/j.brainres.2006.02.115 [12] G. Q. Zhang, X. M. Hao, S. Z. Chen, P. A. Zhou, H. P. Cheng and C. H. Wu, “Blockade of Paeoniflorin on So- dium Current in Mouse Hippocampal CA1 Neurons,” Ac- ta Pharmacologica Sinica, Vol. 24, No. 12, December 2003, pp. 1248-1252. [13] T. Y. Tsai, S. N. Wu, Y. C. Liu, A. Z. Wu and Y. C. Tsai, “Inhibitory Action of L -type C a2+ Current by Paeoniflorin, a Major Constituent of Peony Root, in NG108-15 Neu- ronal Cells,” European. Journal of Pharmacology, Vol. 523, No. 1-3, October 2005, pp. 16-24. doi:10.1016/j.ejphar.2005.08.042 [14] K. S. Murth y, Y. S. Yee, J. R. Grider and G. M. Ma khlouf, “Phorbol-Stimulated Ca2+ Mobilization and Contraction in Dispersed Intestinal Smooth Muscle Cells,” Journal of Pharmacology and Experimental Therapeutics, Vol. 294, No. 3, 2000, pp. 991-996. [15] K. S. Murthy, J. R. Grider, J. F. Kuemmerle and G. M. Makhlouf, “Sustained Muscle Contraction Induced by Agonists, Growth Factors, and Ca2+ Mediated by Distinct PKC Isozymes,” American Journal of Physiology, Ga- strointestinal Liver Physiology, Vol. 279, No. 1, July 2000, pp. G201-G210. [16] F. Nakao, S. Kobayashi, K. Mogami, Y. Mizukami, S. Shi- rao, S. Miwa, N. Todoroki-Ikeda, M. Ito and M. Mats uzaki, “Involvement of Src Family Protein Tyrosine Kinase in Ca2+ Sensitization of Coronary Artery Contraction Mediated by a Sphingosylphosphorylcholin-Rho-Kinase Pathway,” Circulation Research, Vol. 91, 2002, pp. 952-960. doi:10.1161/01.RES.0000042702.04920.BF [17] S-K. Ryu, D. S. Ahn, Y-E. Cho, S-K. Choi, Y-H. Kim, K, G. Morgan and Y-H. Lee, “Augmented-Induced Ca2+- Sensitization of Mesenteric Artery Contraction in Spon- taneous Hypertensive Rat,” Naunyn-Schmiedeberg Arc- hive of Pharmacology, Vol. 373, No. 1, 2006, pp. 30-36. doi:10.1007/s00210-006-0036-7 [18] H. Satoh, “Sino-Atrial Nodal Cells of Mammalian Hearts: Ionic Currents and Gene Expression of Pacemaker Ionic Channels,” J ournal of Smooth Muscle Research, Vol. 39, No. 5, 2003, pp.175-193. doi:10.1540/jsmr.39.175 [19] H. Satoh, “Identification of Hyperpolarization-Activated Inward Current in Uterine Smooth Muscle Cell during Pregnancy,” General Pharmacology, Vol. 26, No. 6, Oc- tober 1995, pp. 1335-1338. doi:10.1016/0306-3623(95)00006-M [20] L. Thomsen, T. L. Robinson, J. C. F. Lee, L. A. Farraway, M. J. G. Hughes, S. W. Andrews and J. D. Huizinga, “In- terstitial Cells of Cajal Generate a Rhythmic Pacemaker Current,” Nature Medicine, Vol. 4, No. 7, July 1998, pp. 848-851. doi:10.1038/nm0798-848 [21] M. T. Nelson, H. Cheng, M. Rubart, L. F. Santana, A. D. Bonev, H. J. Knot and W. J. Lederer, “Relaxation of Ar- terial Smooth Muscle by Calcium Sparks,” Science, Vol. 270, No. 5236, October 1995, pp. 633-637. doi:10.1126/science.270.5236.633 [22] R. ZhuGe, S. M. Sims, R. A. Tuft, K. E. Fogarty and J. V. Jr. Walsh, “Ca2+ Sparks Activate K+ and Cl- Channels Resulting in Spontaneous Transient Currents in Guinea- Pig Tracheal Myocytes,” Journal of Physiology (Lon- don) , Vol. 513, No. 3, December 1998, pp. 711-718. doi:10.1111/j.1469-7793.1998.711ba.x [23] Y. Himeno, F. Toyoda, H. Satoh, A. Amano, C. Y. Cha, H. Matsumura and A. Noma, ”Minor Contribution to Pacemaking in Guinea Pig SA Node,” American Journal of Physiology, Heart Circulation Physiology, Vol. 300, 2011, pp. H251-H261. doi:10.1152/ajpheart.00764.2010 [24] H. Satoh, “Modulation of Ca2+-Activated K+ Current by Isoprenaline, Carbachol, and Phorbol Ester in Cultured (and Fresh) Rat Aortic Vascular Smooth Muscle Cells,” General Pharmacology, Vol. 27, No. 2, March 1996, pp. 319-324. doi:10.1016/0306-3623(95)02005-5 [25] P. Kojodjojo, P. Kanagaratnam, O. R. Segal, W. Hussain and N. S. Peters, “The Effects of Carbenoxolone on Hu- man Myocardial Conduction: A Tool to Investigate the Role of Gap Junction Uncoupling in Human Arrhythmo- genesis,” Journal of the American College of Cardiology, Vol. 48, No. 6, September 2006, pp. 1242-1249. doi:10.1016/j.jacc.2006.04.093 [26] L. C. Chen, M. H. Chou, M. F. Lin and L. L. Yang, “Pharmacokinetics of Paeoniflorin after Oral Administr a- tion of Dshao-yao Gan-chao Tang in Mice,” Japanese Journal of Pharmacology, Vol. 88, No. 3, 2002, pp. 250- 255. doi:10.1254/jjp.88.250 [27] H. Hosseinzadeh, A. M. Nassiri and S. Parvardeh, “The Effects of Carbenoxolone, a Semisynthetic Derivative of Glycyrrhizinic Acid, on Peripheral and Central Ischemia- Reperfusion Injuries in the Skeletal Muscle and Hippo- campus of Rats,” Phytomedicine, Vol. 12, No. 9, Sep- tember 2005, pp. 632-637. doi:10.1016/j.phymed.2004.07.007 [28] I. G. Sava, V. Battaglia, C. A. Rossi, M. Salvi and A. Toni- nello, “Free Radical Scavenging Action of the Natual Po- lyamine Spermine in Rat Liver Mitochondria,” Free Radi- cal Biology and Medicine, Vol. 41, No. 8, October 2006, pp. 1272-1281. doi:10.1016/j.freeradbiomed.2006.07.008  H. SATOH ET AL. Copyright © 2011 SciRes. CM [29] H. Y. Park, S. H. Pa rk, H. K. Yoon, M. J. Han and D. H. Kim, “Anti-Allergic Activity of 18-β-Glycyrrhetinic Ac- id-3-O-β-D-Glucuronide,” Archives of Pharmacal Re- search, Vol. 27, 2004, pp. 57-60. doi:10.1007/BF02980047 [30] O. H. Kang, J. A. Kim, Y. A. Choi, H. J. Park, D. K. Kim, Y. H. An, S. C. Choi, K. J. Yun, Y. H. Nah, X. F. Cai, Y. H. Kim, K. H. Bae and Y. M. Lee, “Inhibition OH Inter- leukin-8 Production in the Human Col onic Epit helia l Cell Line HT-29 by 18-β-Glycyrrhetinic Acid,” International Journal of Molecular Medicine, Vol. 15, 2005, pp. 981- 985. [31] H. Kato, M. Kanaoka, S. Yano and M. Kobayashi, “3- Monoglucuronyl-Glycyrrehetic Acid is a Major Metabo- lite That Causes Licorice-Induced Pseudoaldosteronism,” Journal of Clinical Endocrinology and Metabolism, Vol. 80, No. 6, June 1995, pp. 1929-1933. Abbreviations ACh: acetylcholine ATP: adenosine triphosphate ANOVA: analysis of varience CA: hippocampal neuron CCh: carbachol DMSO: dimethyl sulfoxide T1/2: half-life IBMX: 3-isobutyl-1-methylxanthine If: hyperpolarization-activated inward (or pacemaker) current IL: interleukin IP3: inositol triphosphate KCa: Ca2+-activated K+ channel M1 receptor: muscarinic receptor MLCK: myosin light chain kinase PDE: phosphodiesterase PK-C: protein kinase C SA node: sino-atrial node SR: sarcoplasmic reticulum TEA: tetraethyammonium
|