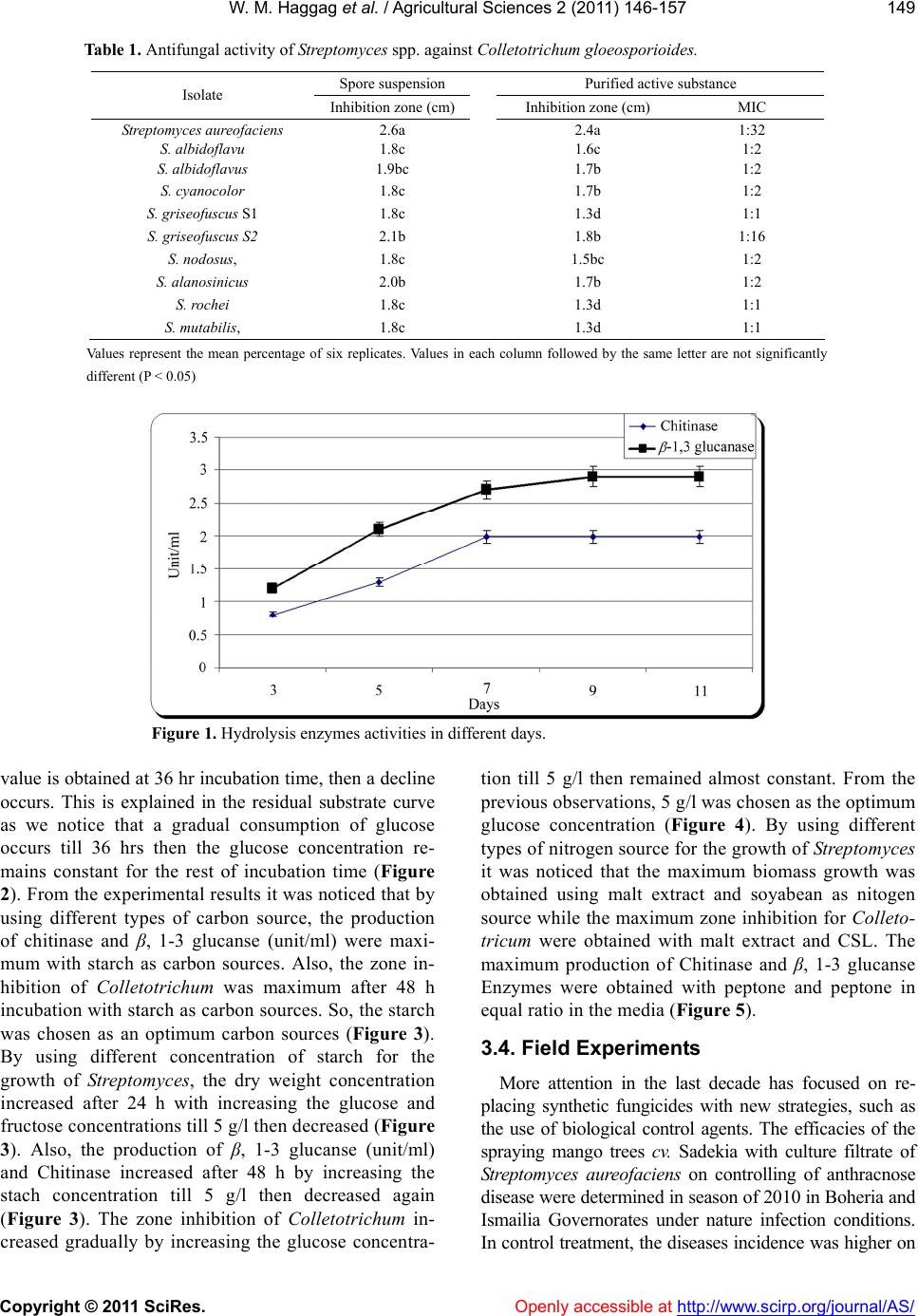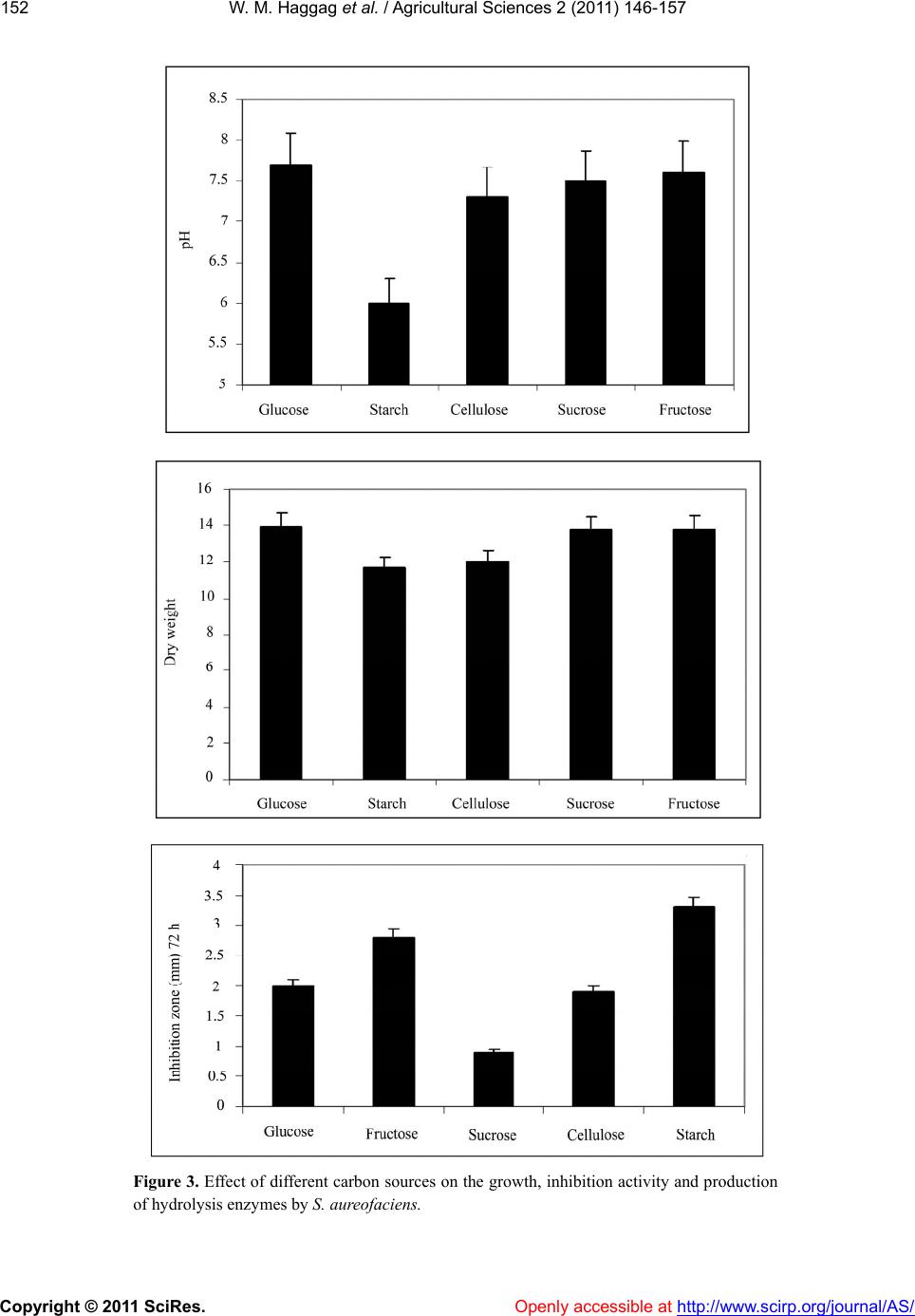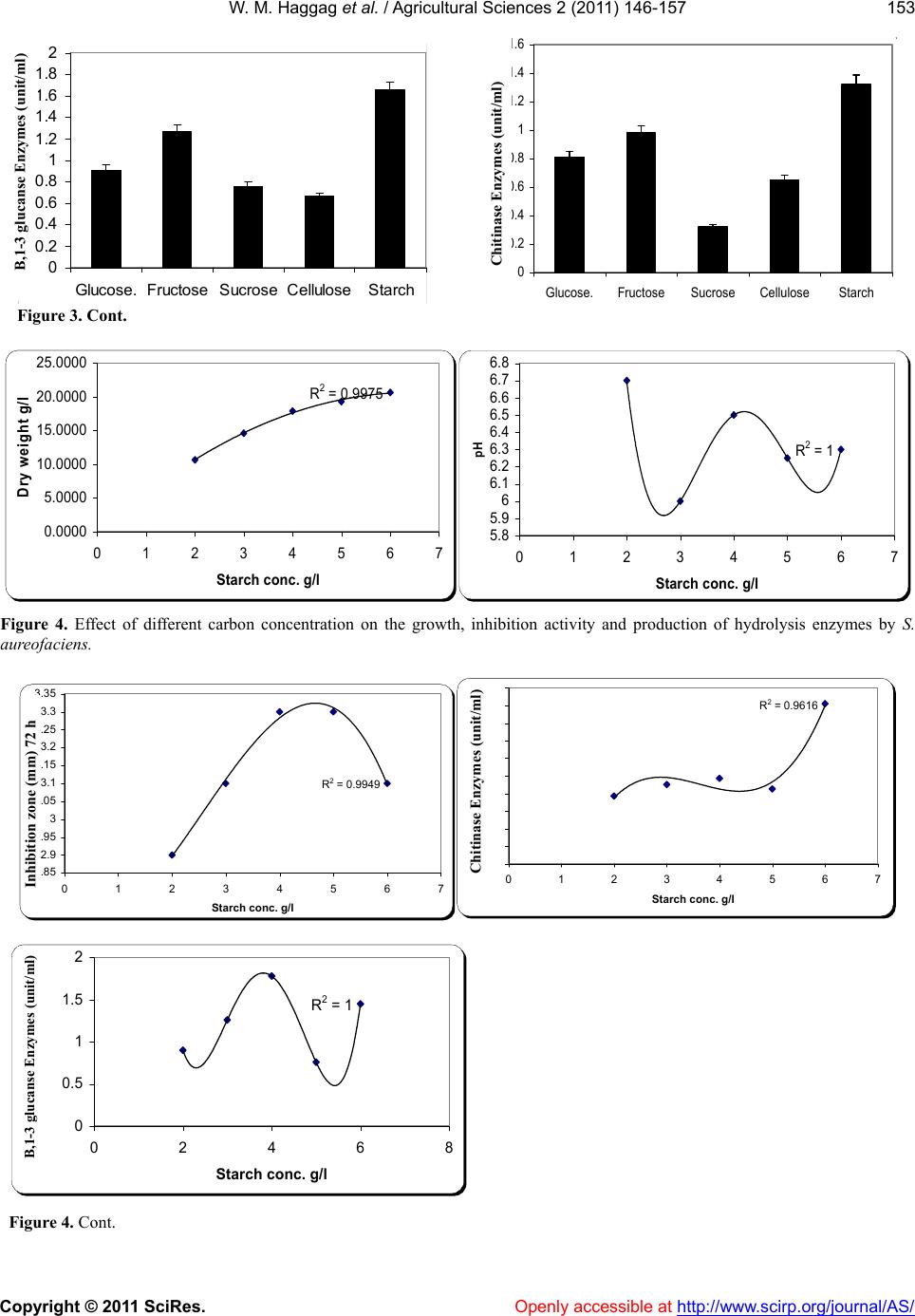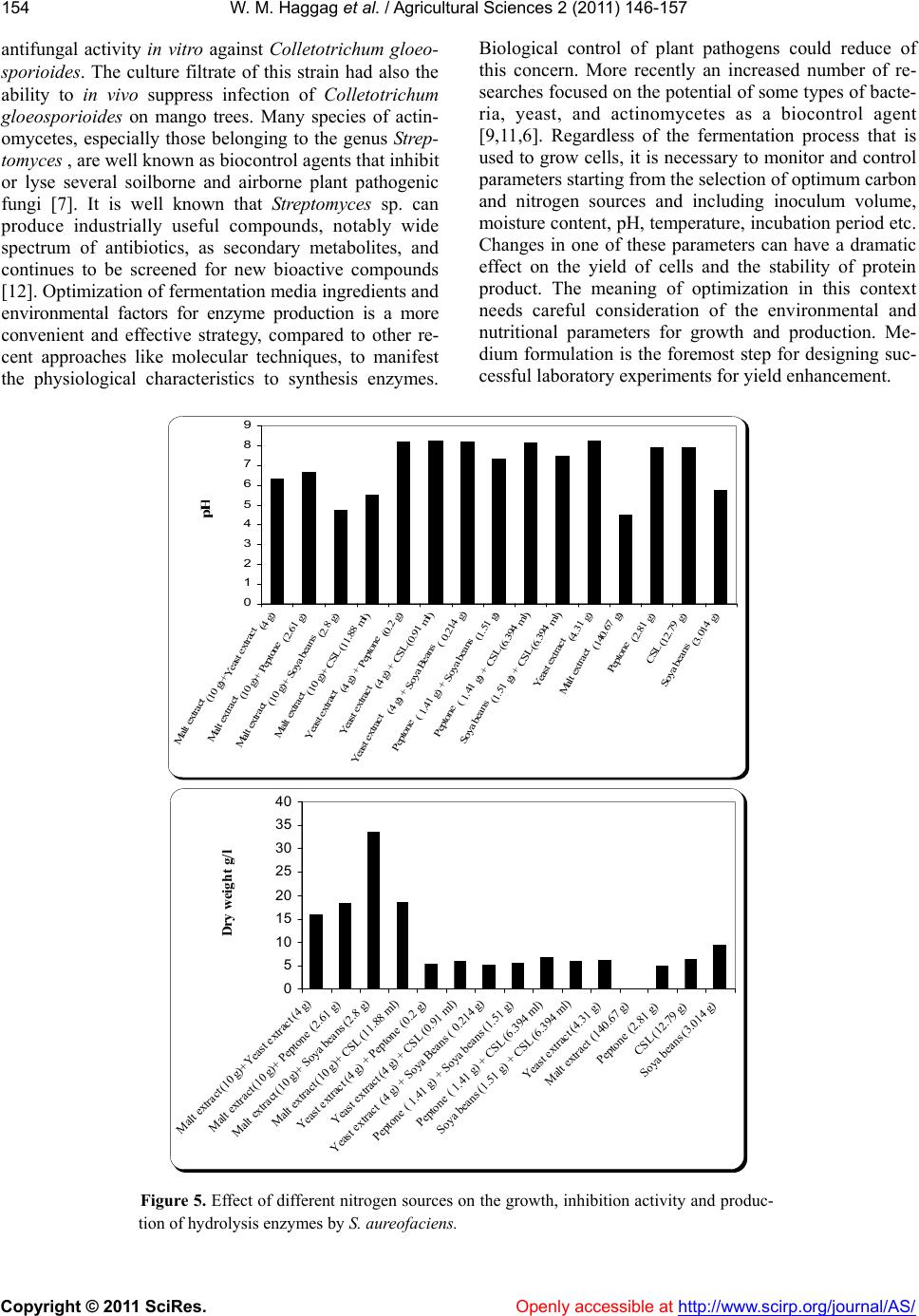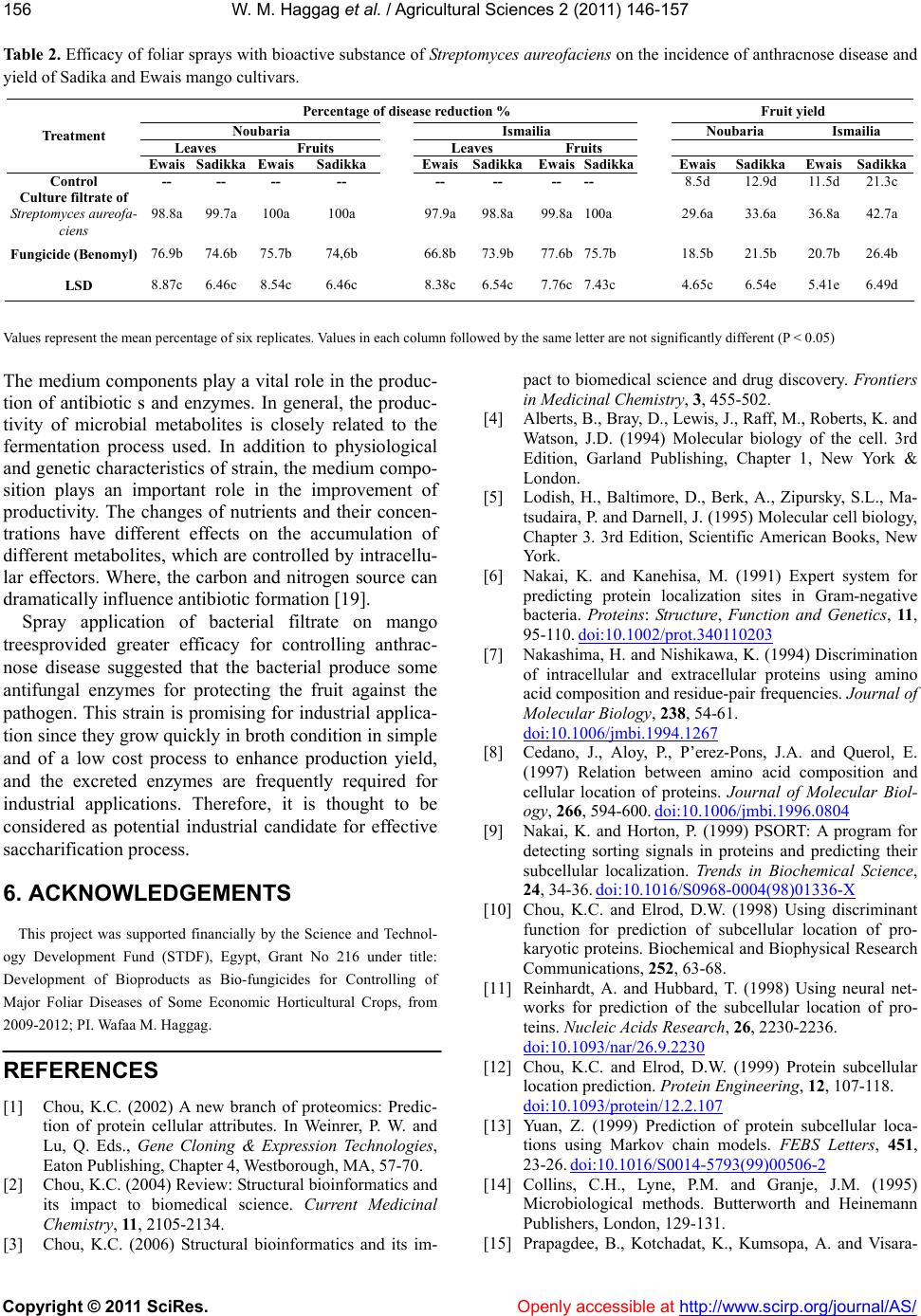
W. M. Haggag et al. / Agricultural Sciences 2 (2011) 146-157
Copyright © 2011 SciRes. http://www.scirp.org/journal/AS/
156
Table 2. Efficacy of foliar sprays with bioactive substance of Streptomyces aureofaciens on the incidence of anthracnose disease and
yield of Sadika and Ewais mango cultivars.
Percentage of disease reduction % Fruit yield
Noubaria Ismailia Noubaria Ismailia
Leaves Fruits Leaves Fruits
Treatment
Ewais Sadikka Ewais Sadikka EwaisSadikkaEwaisSadikka Ewais Sadikka EwaisSadikka
Control -- -- -- -- -- -- -- --
8.5d 12.9d 11.5d21.3c
Culture filtrate of
Streptomyces aureofa-
ciens 98.8a 99.7a100a 100a 97.9a98.8a 99.8a100a 29.6a 33.6a 36.8a42.7a
Fungicide (Benomyl) 76.9b 74.6b 75.7b 74,6b 66.8b73.9b 77.6b75.7b 18.5b 21.5b 20.7b26.4b
LSD 8.87c 6.46c 8.54c 6.46c 8.38c6.54c 7.76c7.43c 4.65c 6.54e 5.41e6.49d
Values represent the mean percentage of six replicates. Values in each column followed by the same letter are not significantly different (P < 0.05)
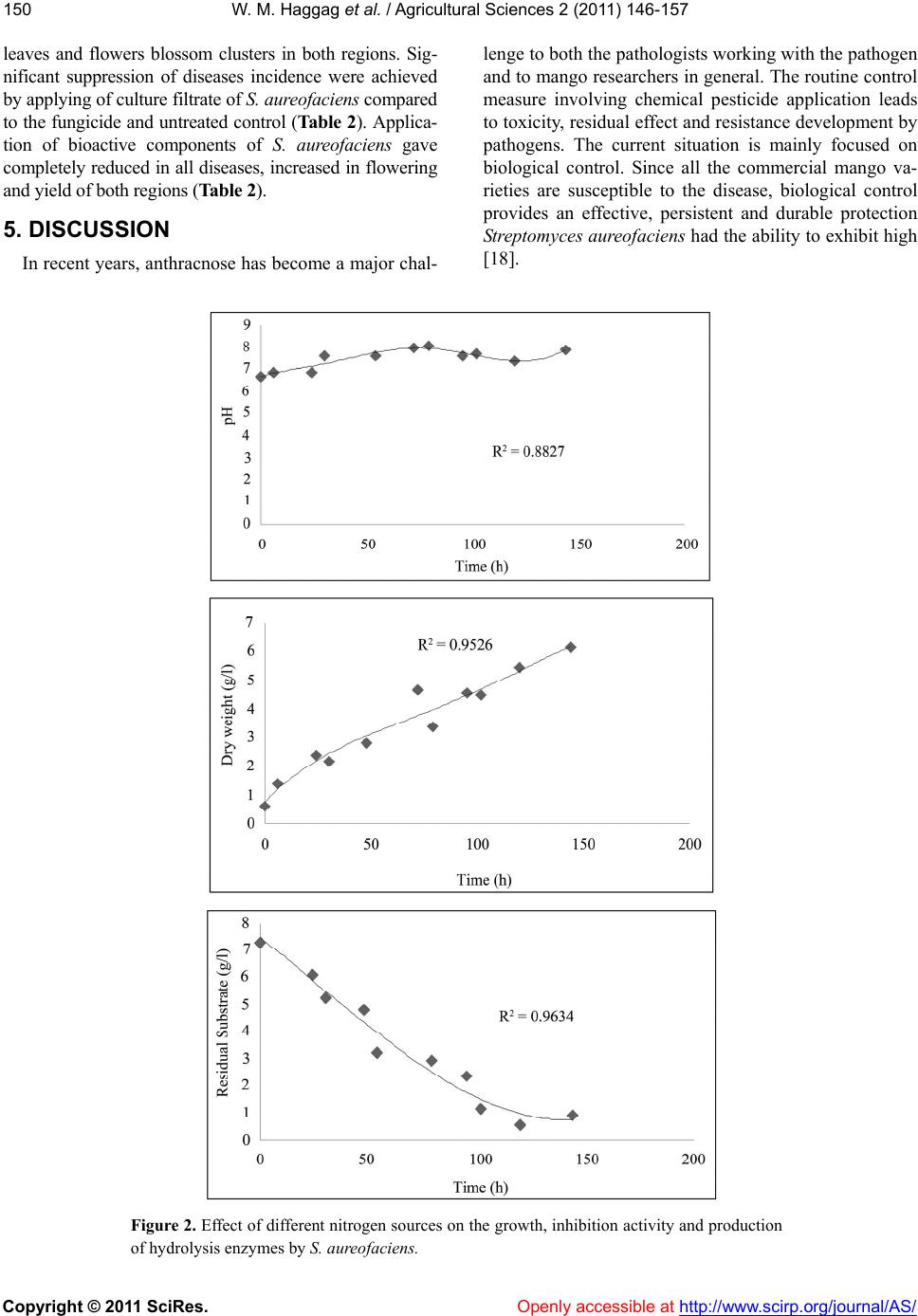
The medium components play a vital role in the produc-
tion of antibiotic s and enzymes. In general, the produc-
tivity of microbial metabolites is closely related to the
fermentation process used. In addition to physiological
and genetic characteristics of strain, the medium compo-
sition plays an important role in the improvement of
productivity. The changes of nutrients and their concen-
trations have different effects on the accumulation of
different metabolites, which are controlled by intracellu-
lar effectors. Where, the carbon and nitrogen source can
dramatically influence antibiotic formation [19].
Openly accessible at
Spray application of bacterial filtrate on mango
treesprovided greater efficacy for controlling anthrac-
nose disease suggested that the bacterial produce some
antifungal enzymes for protecting the fruit against the
pathogen. This strain is promising for industrial applica-
tion since they grow quickly in broth condition in simple
and of a low cost process to enhance production yield,
and the excreted enzymes are frequently required for
industrial applications. Therefore, it is thought to be
considered as potential industrial candidate for effective
saccharification process.
6. ACKNOWLEDGEMENTS
This project was supported financially by the Science and Technol-
ogy Development Fund (STDF), Egypt, Grant No 216 under title:
Development of Bioproducts as Bio-fungicides for Controlling of
Major Foliar Diseases of Some Economic Horticultural Crops, from
2009-2012; PI. Wafaa M. Haggag.
REFERENCES
[1] Chou, K.C. (2002) A new branch of proteomics: Predic-
tion of protein cellular attributes. In Weinrer, P. W. and
Lu, Q. Eds., Gene Cloning & Expression Technologies,
Eaton Publishing, Chapter 4, Westborough, MA, 57-70.
[2] Chou, K.C. (2004) Review: Structural bioinformatics and
its impact to biomedical science. Current Medicinal
Chemistry, 11 , 2105-2134.
[3] Chou, K.C. (2006) Structural bioinformatics and its im-
pact to biomedical science and drug discovery. Frontiers
in Medicinal Chemistry, 3, 455-502.
[4] Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. and
Watson, J.D. (1994) Molecular biology of the cell. 3rd
Edition, Garland Publishing, Chapter 1, New York &
London.
[5] Lodish, H., Baltimore, D., Berk, A., Zipursky, S.L., Ma-
tsudaira, P. and Darnell, J. (1995) Molecular cell biology,
Chapter 3. 3rd Edition, Scientific American Books, New
York.
[6] Nakai, K. and Kanehisa, M. (1991) Expert system for
predicting protein localization sites in Gram-negative
bacteria. Proteins: Structure, Function and Genetics, 11,
95-110. doi:10.1002/prot.340110203
[7] Nakashima, H. and Nishikawa, K. (1994) Discrimination
of intracellular and extracellular proteins using amino
acid composition and residue-pair frequencies. Journal of
Molecular Biology, 238, 54-61.
doi:10.1006/jmbi.1994.1267
[8] Cedano, J., Aloy, P., P’erez-Pons, J.A. and Querol, E.
(1997) Relation between amino acid composition and
cellular location of proteins. Journal of Molecular Biol-
ogy, 266, 594-600. doi:10.1006/jmbi.1996.0804
[9] Nakai, K. and Horton, P. (1999) PSORT: A program for
detecting sorting signals in proteins and predicting their
subcellular localization. Trends in Biochemical Science,
24, 34-36. doi:10.1016/S0968-0004(98)01336-X
[10] Chou, K.C. and Elrod, D.W. (1998) Using discriminant
function for prediction of subcellular location of pro-
karyotic proteins. Biochemical and Biophysical Research
Communications, 252, 63-68.
[11] Reinhardt, A. and Hubbard, T. (1998) Using neural net-
works for prediction of the subcellular location of pro-
teins. Nucleic Acids Research, 26, 2230-2236.
doi:10.1093/nar/26.9.2230
[12] Chou, K.C. and Elrod, D.W. (1999) Protein subcellular
location prediction. Protein Engineering, 12, 107-118.
doi:10.1093/protein/12.2.107
[13] Yuan, Z. (1999) Prediction of protein subcellular loca-
tions using Markov chain models. FEBS Letters, 451,
23-26. doi:10.1016/S0014-5793(99)00506-2
[14] Collins, C.H., Lyne, P.M. and Granje, J.M. (1995)
Microbiological methods. Butterworth and Heinemann
Publishers, London, 129-131.
[15] Prapagdee, B., Kotchadat, K., Kumsopa, A. and Visara-