Improved Comb Filter Based Approach for Effective Prediction of Protein Coding Regions in DNA Sequences
98
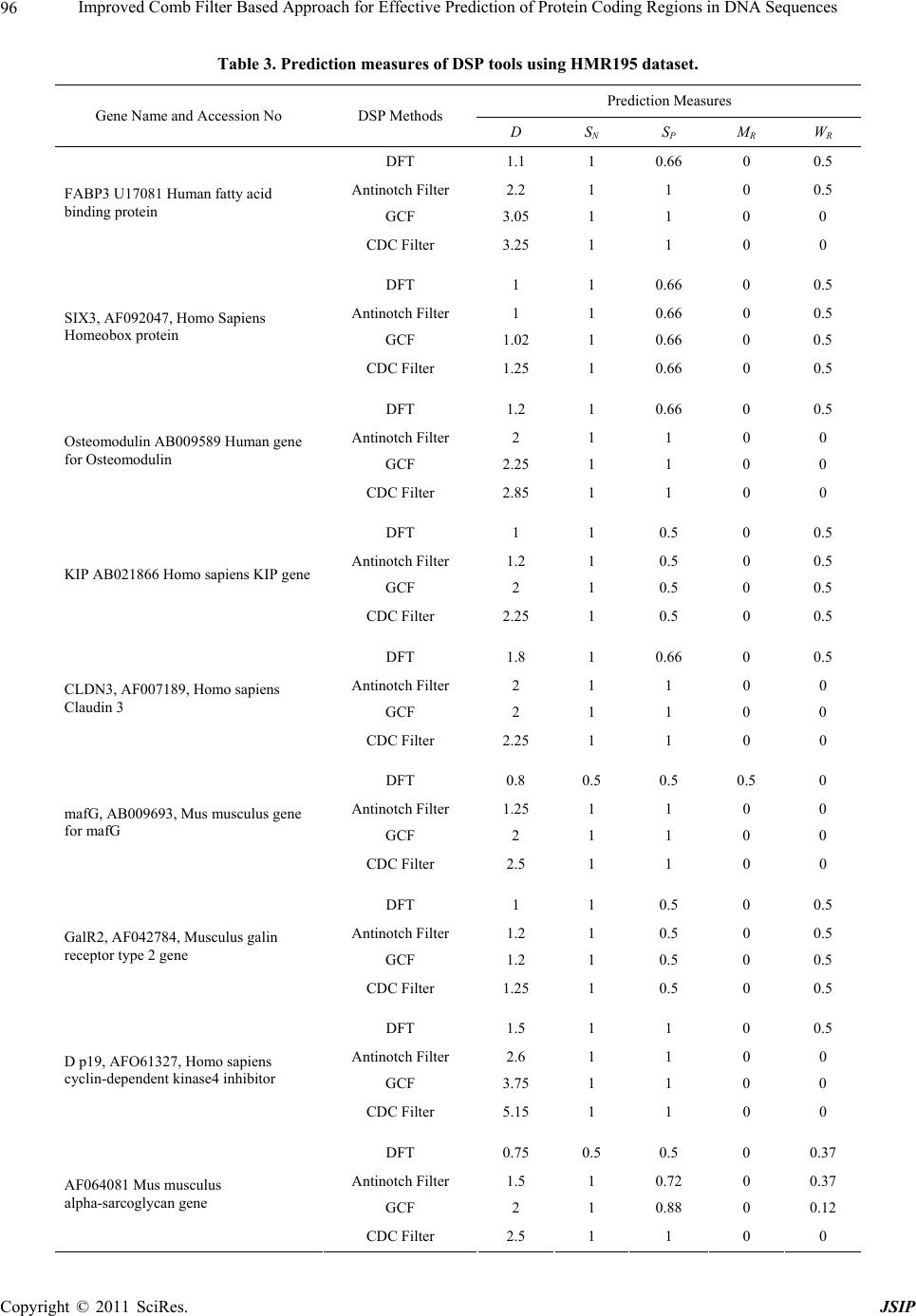
Table 4. Computational complexities of DSP methods for
prediction of protein coding region using period-3 property.
Prediction Technique Multiplications Additions
Using N-point DFT N2(C) N(N – 1)(C)
Using N-point FFT (N/2)log2N(C) Nlog2N(C)
Allpass Antinotch Filtering (2N + 1)(R) 2N(R)
Multistage Filtering 5(R) 2N(R)
Generalized Comb Filtering 3(R) 2 (R)
CDC Filtering NIL 2 (R)
‘C’ and ‘R’ refer to complex and real arithmetic operations respectively.
can therefore be used as an efficient tool for the identifi-
cation of protein coding regions of DNA sequences. As
such comb filter is simple in structure, involves less
computational complexity and better prediction effi-
ciency compared to the FFT-based methods and other
digital filtering methods. Hence it can be used as a com-
putationally efficient and better alternative to other DSP
approach to the prediction of protein coding regions.
REFERENCES
[1] Z. Wang, Y. Z. Chen and Y. X. Li, “A Brief Review of
Computational Gene Prediction Methods,” Genomics Pro-
teomics Bioinformatics, Vol. 2, No. 4, 2004, pp. 216-221.
[2] D. Anastassiou, “Genomic Signal Processing,” Signal
Processing Magazine, Vol. 18, No. 4, 2001, pp. 8-20.
doi:10.1109/79.939833
[3] J. W. Fickett, “The Gene Identification Problem: Over-
view for Developers,” Computers & Chemistry, Vol. 20,
No. 1, 1996, pp. 103-118.
doi:10.1016/S0097-8485(96)80012-X
[4] R. Voss, “Evolution of Long-Range Fractal Correlations
and 1/f Noise in DNA Base Sequences,” Physical Review
Letters, Vol. 68, No. 25, 1992, pp. 3805-3808.
doi:10.1103/PhysRevLett.68.3805
[5] P. D. Cristea, “Genetic signal Representation and Analy-
sis,” Proceedings of SPIE Conference, International
Biomedical Optics Symposium (BIOS'02), Vol. 4623, 2002,
pp. 77-84.
[6] A. K. Brodzik and O. Peters, “Symbol-Balanced Quater-
nionic Periodicity Transform for Latent Pattern Detection
in DNA Sequences,” IEEE International Conference on
Acoustics, Speech, and Signal Processing (ICASSP '05),
Vol. 5, 2005, pp. 373-376.
[7] T. M. Nair, S. S. Tambe and B. D. Kulkarni, “Application
of Artificial Neural Networks for Prokaryotic Transcrip-
tion Terminator Prediction,” FEBS Letters, Vol. 346, No.
2-3, 1994, pp. 273-277.
doi:10.1016/0014-5793(94)00489-7
[8] A. S. Nair and S. P. Sreenathan, “A Coding Measure
Scheme Employing Electron-Ion Interaction Pseudopo-
tential (EIIP),” Bioinformation, Vol. 1, No. 6, 2006, pp.
197-202.
[9] G. L. Rosen, “Signal Processing for Biologically-Inspired
Gradient Source Localization and DNA Sequence Analy-
sis,” Ph.D. Thesis, Georgia Institute of Technology, At-
lanta, 2006.
[10] A. S. Nair and S. P. Sreenathan, “An Improved Digital
Filtering Technique Using Frequency Indicators for Lo-
cating Exons,” Journal of the Computer Society of India,
Vol. 36, No. 1, 2006.
[11] R. Zhang and C. T. Zhang, “Z Curves, an Intuitive Tool
for Visualizing and Analyzing the DNA Sequences,”
Journal of Biomolecular Structure & Dynamics, Vol. 11,
No. 4, 1994, pp. 767-782.
[12] A. Rushdi and J. Tuqan, “Gene Identification Using the
Z-Curve Representation,” IEEE International Conference
on Acoustics, Speech and Signal Processing, Toulouse,
14-19 May 2006, pp. 1024-1027.
[13] M. Akhtar, J. Epps and E. Ambikairajah, “On DNA Nu-
merical Representations for Period-3 Based Exon Predic-
tion,” IEEE International Workshop on Genomic Signal
Processing and Statistics, Tuusula, 2007.
[14] B. D. Silverman and R. Linsker, “A Measure of DNA
Periodicity,” Journal of Theoretical Biology, Vol. 118,
No. 3, 1986, pp. 295-300.
doi:10.1016/S0022-5193(86)80060-1
[15] S. Tiwari, S. Ramachandran, A. Bhattacharya, S. Bhatta-
charya and R. Ramaswamy, “Prediction of Probable
Genes by Fourier Analysis of Genomic Sequences,” Bio-
informatics, Vol. 13, No. 3, 1997, pp. 263-270.
doi:10.1093/bioinformatics/13.3.263
[16] D. Anastassiou, “Digital Signal Processing of Bio-
molecular Sequences,” Technical Report, Columbia Uni-
versity, 2000-20-041, April 2000.
[17] D. Anastassiou, “Frequency-Domain Analysis of Bio-
molecular Sequences,” Bioinformatics, Vol. 16, No. 12,
2000, pp. 1073-1082.
doi:10.1093/bioinformatics/16.12.1073
[18] P. P. Vaidyanathan and B. J. Yoon, “Digital Filters for
Gene Prediction Applications,” IEEE Asilomar on Signals,
Systems, and Computers, Monterey, 3-6 November 2002,
pp. 306-310.
[19] P. P. Vaidyanathan and B. J. Yoon, “The Role of Signal
Processing Concepts in Genomics and Proteomics,”
Journal of the Franklin Institute, Vol. 341, No. 1-2, 2004,
pp. 111-135. doi:10.1016/j.jfranklin.2003.12.001
[20] D. Koltar and Y. Lavner, “Gene Prediction by Spectral
Rotation (SR) Measure: A New Method for Identifying
Protein-Coding Regions,” Genome Research, Vol. 13, No.
8, 2003, pp. 1930-1937.
[21] A. Fuentes, J. Ginori and R. Abalo, “A New Predictor of
Coding Regions in Genomic Sequences Using a Combi-
nation of Different Approaches,” International Journal of
Biomedical and Life Sciences, Vol. 3, No. 2, 2007, pp.
1-5.
Copyright © 2011 SciRes. JSIP