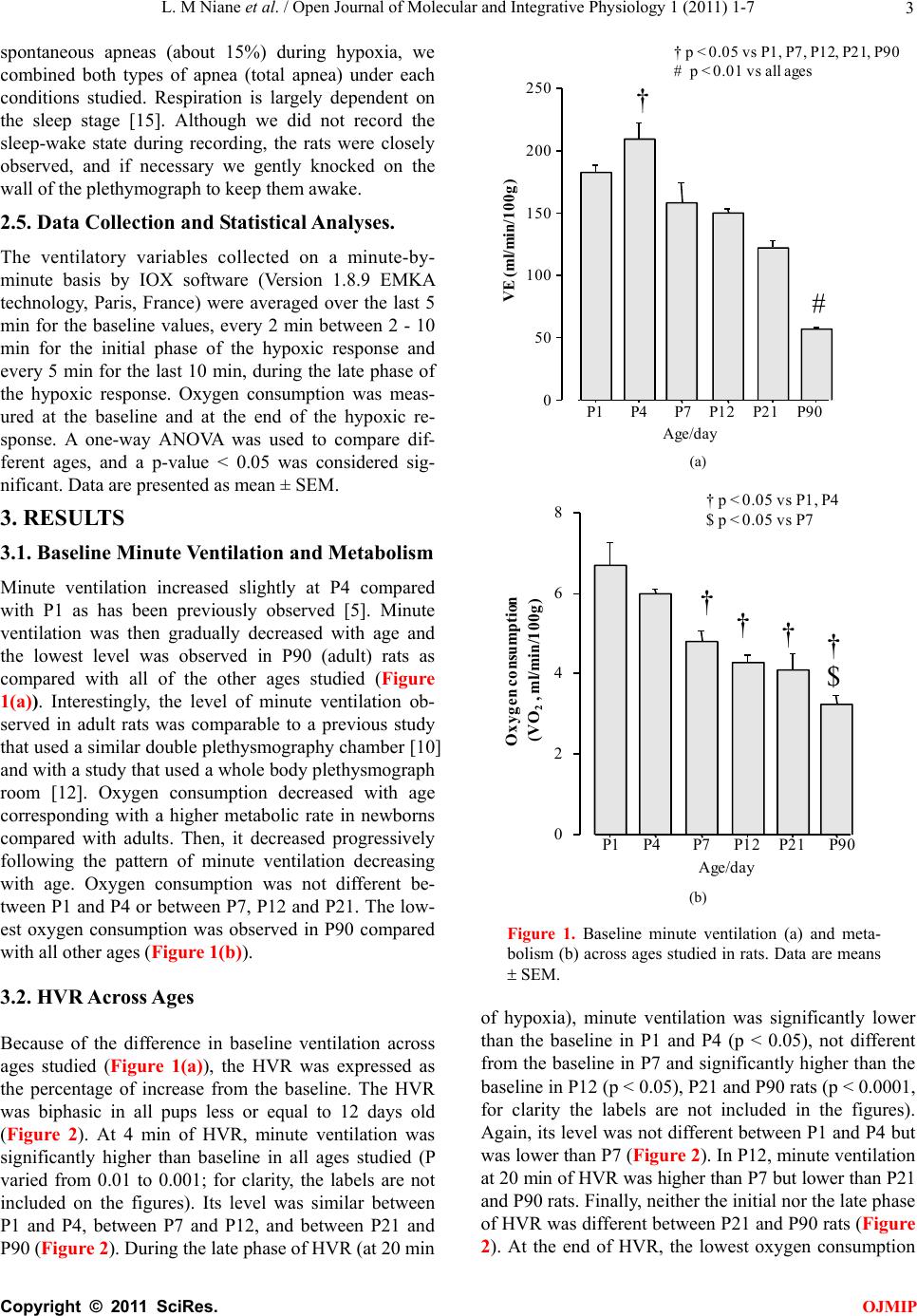
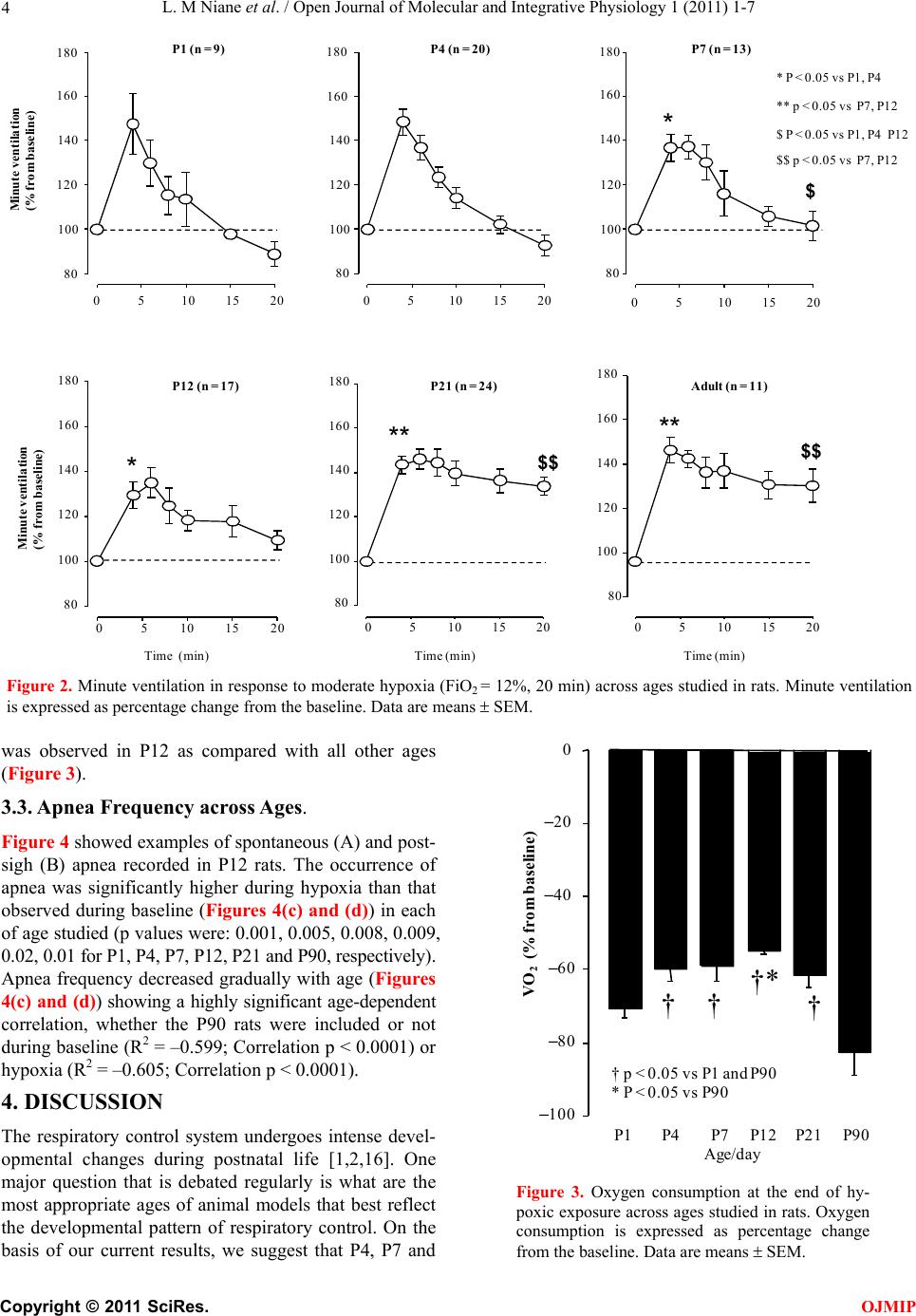
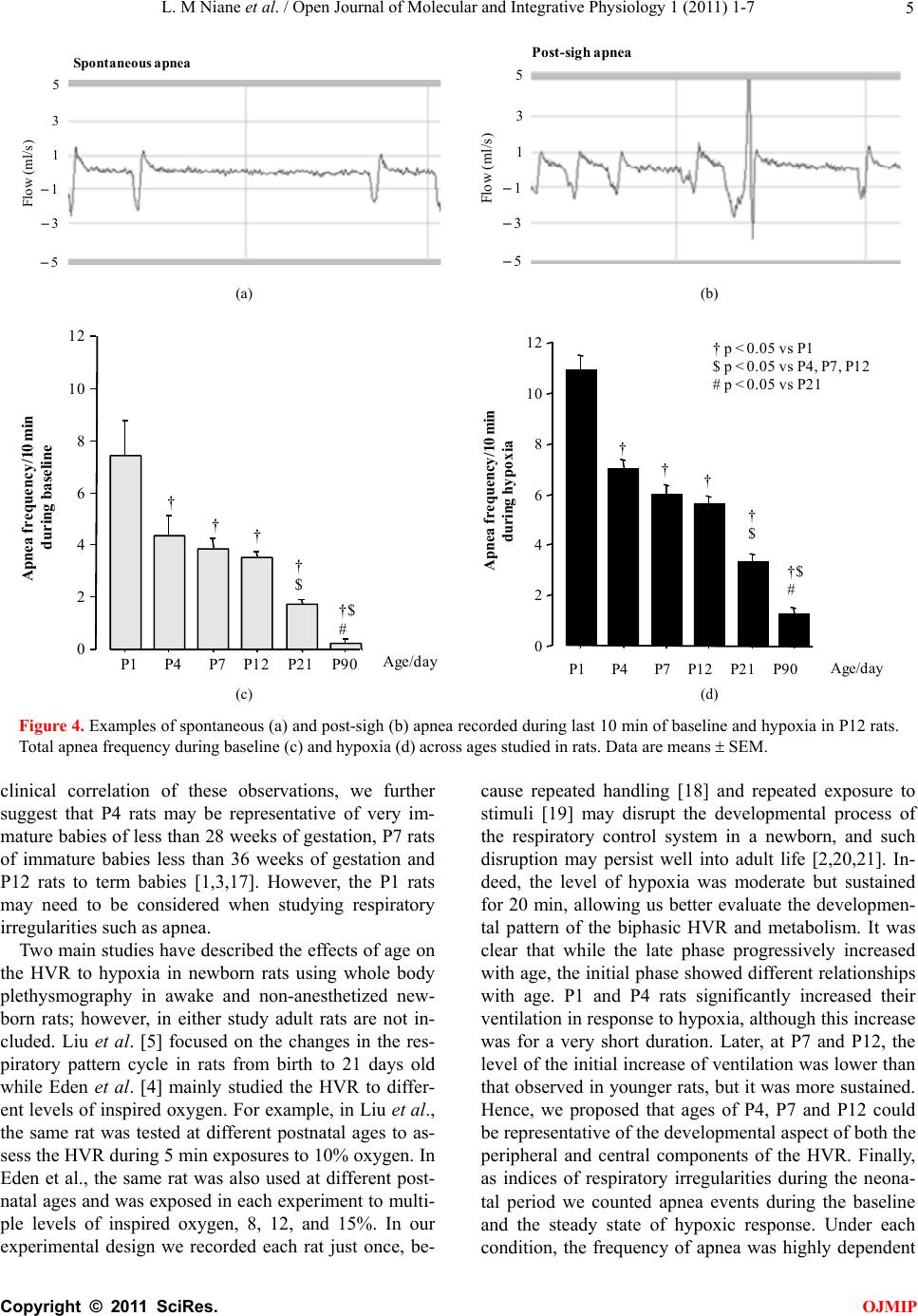
L. M Niane et al. / Open Journal of Molecular and Integrative Physiology 1 (2011) 1-7
6
on age providing a useful index of immaturity of the
respiratory control system. It is suggested that apnea
frequency can be used as an efficient parameter to study
respiratory irregularities in rats and an additional vari-
able to study the development of respiratory control.
One pitfall related to the use of whole body plethys-
mography is the accuracy of tidal volume measurements.
Despite this limitation, this method remains the best
available for measuring ventilatory variables (including
tidal volume) in awake, unrestrained small animals
[11,22,23]. As discussed previously [8,9,14,12], all ex-
periments were conducted under similar conditions, and
the tidal volume was corrected by considering baro-
metric pressure, humidity in the plethysmograph, body
temperature and body weight. Conversely, ventilation is
affected by sleep state [15], and rats during the first 15
days of life spend about 70% of their time asleep [24,25].
However, rats were carefully observed during recording,
and if they showed signs of falling asleep, we gently
knocked on the wall of the recording chamber to keep
the rat awake [12,14].
In conclusion, respiratory irregularities such as apnea
and periodic breathing are frequently observed in pre-
term infants. These irregularities are related to an imma-
turity of the breathing control system [17], and rats are
regularly used as a model. We propose that rats at 4, 7
and 12 days old could be used to study the developmen-
tal pattern of mechanisms, factors or drugs that affect the
HVR. However, rats at P1 old would be included to bet-
ter describe the age-dependence of apnea frequency.
5. ACKNOWLEDGEMENTS
This study was supported, in part, by the CIHR operating grant MOP-
81101 to A. Bairam. We thank Mrs. Melanie Pelletier and Sylvie Viger
for animal care.
REFERENCES
[1] Bissonnette, J.M. (2000) Mechanisms regulating hypoxic
respiratory depression during fetal and postnatal life.
American Journal of Physiology Regulatory, Integrative
and Comparative Physiology, 278, R 1391-1400.
[2] Carroll, J.L. (2003) Developmental plasticity in respira-
tory control. Journal of Applied Physiology, 94, 375-389.
[3] Cohen, G. and Katz-Salamon, M. (2005) Development of
chemoreceptor responses in infants. Respiratory Physi-
ology & Neurobiology, 149, 233-242.
doi:10.1016/j.resp.2005.02.013
[4] Eden, G.J. and Hanson, M.A. (1987) Maturation of the
respiratory response to acute hypoxia in the newborn rat.
Journal of Physiology, 392, 1-9.
[5] Liu, Q., Lowry, T.F. and Wong-Riley, M.T. (2006) Post-
natal changes in ventilation during normoxia and acute
hypoxia in the rat: implication for a sensitive period.
Journal of Physiology, 577, 957-970.
doi:10.1113/jphysiol.2006.121970
[6] Romijn, H.J., Hofman, M.A. and Gramsbergen, A. (1991)
At what age is the developing cerebral cortex of the rat
comparable to that of the full-term newborn human baby?
Early Human Development, 26, 61-67.
doi:10.1016/0378-3782(91)90044-4
[7] Behan, M. and Wenninger, J.M. (2008) Sex steroidal
hormones and respiratory control. Respiratory Physiol-
ogy & Neurobiology, 164, 213-221.
doi:10.1016/j.resp.2008.06.006
[8] Julien, C., Bairam, A. and Joseph, V. (2008) Chronic
intermittent hypoxia reduces ventilatory long-term fa-
cilitation and enhances apnea frequency in newborn rats.
American Journal of Physiology Regulatory, Integrative
and Comparative Physiology, 294, R1356-1366.
doi:10.1152/ajpregu.00884.2007
[9] Niane, L.M., Donnelly, D.F., Joseph, V. and Bairam, A.
(2010) Ventilatory and carotid body chemoreceptor re-
sponses to purinergic P2X receptor antagonists in new-
born rats. Journal of Applied Physiology, 110, 83-94.
doi:10.1152/japplphysiol.00871.2010
[10] Julien, C.A., Niane, L., Kinkead, R., Bairam, A. and
Joseph, V. (2010) Carotid sinus nerve stimulation, but not
intermittent hypoxia, induces respiratory LTF in adult
rats exposed to neonatal intermittent hypoxia. American
Journal of Physiology Regulatory, Integrative and Com-
parative Physiology, 299, R192-205.
doi:10.1152/ajpregu.00707.2009
[11] Bartlett, D.Jr. and Tenney, S.M. (1970) Control of
breathing in experimental anemia. Respiratory Physiol-
ogy, 10, 384-395. doi:10.1016/0034-5687(70)90056-3
[12] Montandon, G., Bairam, A. and Kinkead, R. (2006)
Long-term consequences of neonatal caffeine on ventila-
tion, occurrence of apneas, and hypercapnic chemoreflex
in male and female rats. Pediatric Research, 59, 519-524.
doi:10.1203/01.pdr.0000203105.63246.8a
[13] Mendelson, W.B., Martin, J.V., Perlis, M., Giesen, H.,
Wagner, R. and Rapoport, S.I. (1988) Periodic cessation
of respiratory effort during sleep in adult rats. Physiology
& Behavior, 43, 229-234.
doi:10.1016/0031-9384(88)90243-0
[14] Julien, C.A., Joseph, V. and Bairam, A. (2010) Caffeine
reduces apnea frequency and enhances ventilatory
long-term facilitation in rat pups raised in chronic inter-
mittent hypoxia. Pediatric Research, 68, 105-111.
doi:10.1203/PDR.0b013e3181e5bc78
[15] Parmeggiani, P.L. (1985) Regulation of circulation and
breathing during sleep: experimental aspects. Annals of
Clinical Research, 17, 185-189.
[16] Putnam, R.W., Conrad, S.C., Gdovin, M.J., Erlichman,
J.S. and Leiter, J.C. (2005) Neonatal maturation of the
hypercapnic ventilatory response and central neural CO2
chemosensitivity. Respiratory Physiology & Neurobiol-
ogy, 149, 165-179. doi:10.1016/j.resp.2005.03.004
[17] Darnall, R.A., Ariagno, R.L. and Kinney, H.C. (2006)
The late preterm infant and the control of breathing,
sleep, and brainstem development: a review. Clinics in
Perinatology, 33, 883-914. doi:10.1016/j.clp.2006.10.004
[18] Gulemetova, R. and Kinkead, R. (2011) Neonatal stress
increases respiratory instability in rat pups. Respiratory
Physiology & Neurobiology, in press.
doi:10.1016/j.resp.2011.01.014
[19] Mateika, J.H. and Narwani, G. (2009) Intermittent hy-
C
opyright © 2011 SciRes. OJMIP