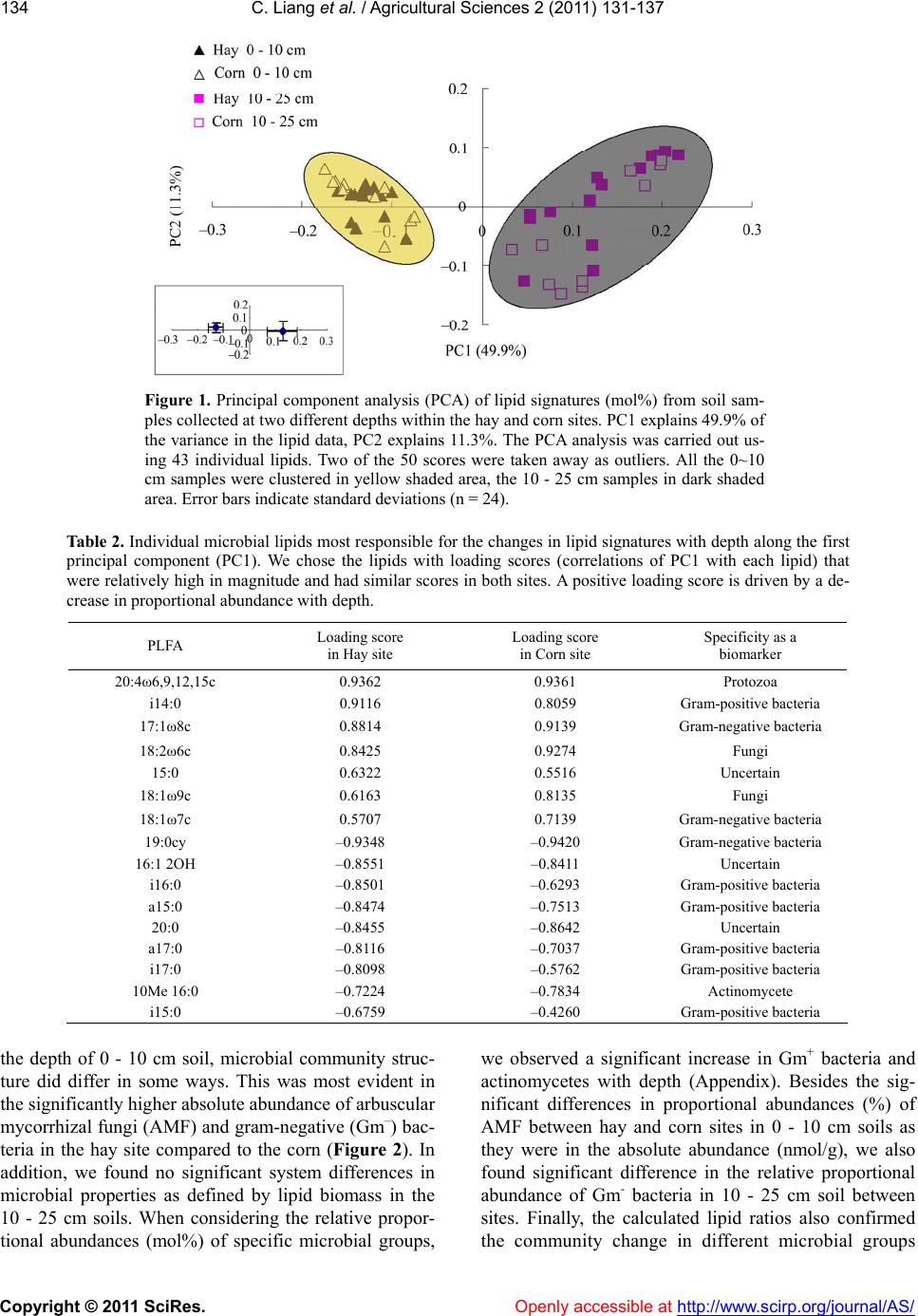
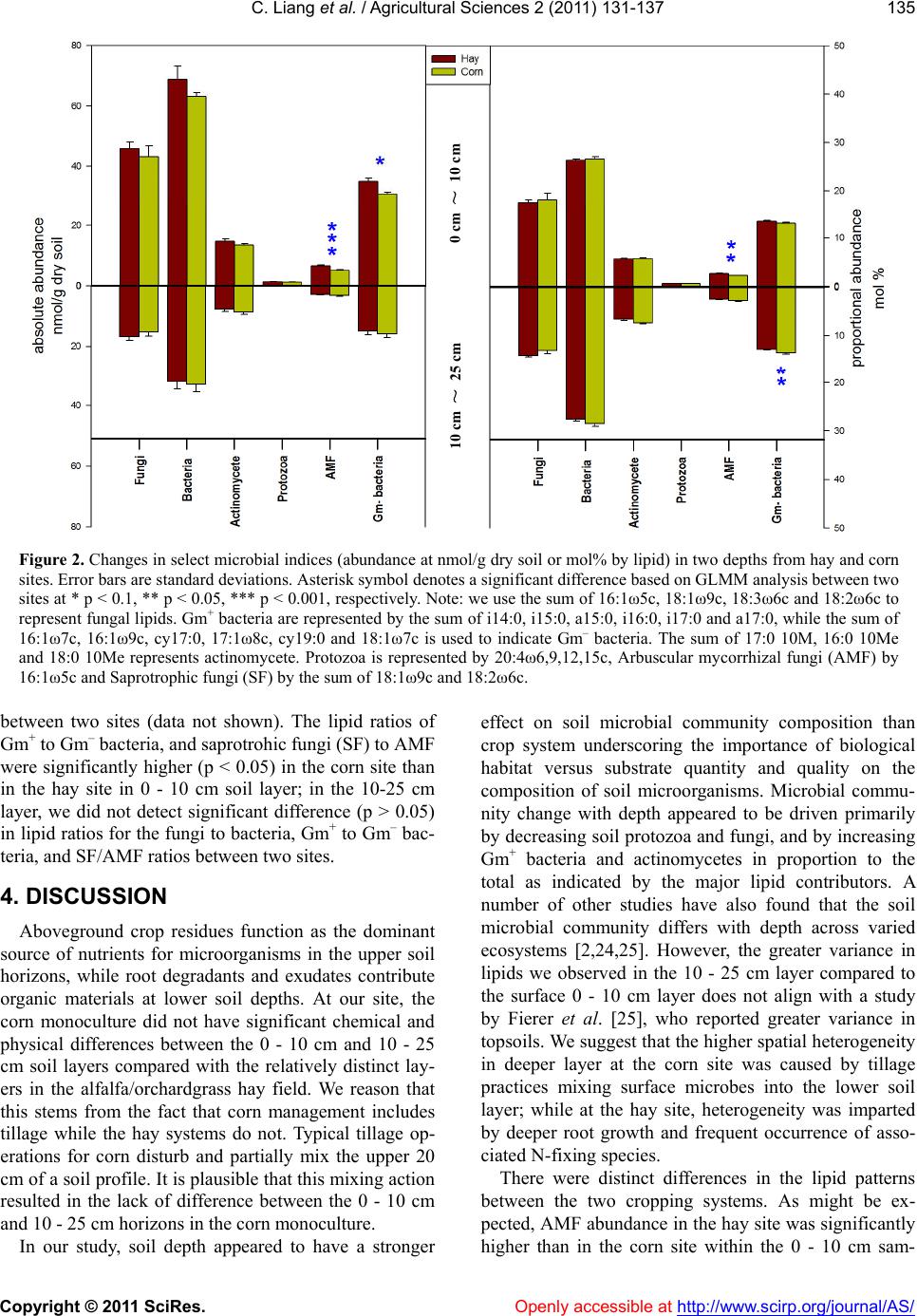
C. Liang et al. / Agricultural Sciences 2 (2011) 131-137
Copyright © 2011 SciRes. Openly accessible at http://www.scirp.org/journal/AS/
136
pling depth. This was likely to the result of tillage in the
corn site, which destroys fungal hyphae [26] and dis-
rupts the signaling action initiated by rhizobial nodula-
tion that can also stimulate mycorrhizal colonization
[27]. Historic tillage may have affected Gm– bacteria as
well since higher absolute abundance of Gm– bacteria
and fungi were found in non-tilled than tilled 0 - 5 cm
soil [28]. Higher Gm– bacterial abundance at the hay site
in the 0 - 10 cm depth may be also explained by the en-
riched symbiotic Gm– N-fixing rhizobia with legume
alfalfa plants, as the rhizosphere usually harbors more
Gm– bacteria and fewer Gm+ bacteria [29].
In conclusion, we found that total microbial biomass
was constant between hay and corn sites, but markedly
decreased at the 10 - 25 cm depth in a southern Wiscon-
sin agroecosystem. Despite similar biomass between
sites, or distinct biomass between depths, there were
distinct microbial communities with a greater change in
composition and biomass between depths than between
sites. These results suggest habitat (depth in soils) may
be more important in controlling microbial community
composition than plant species and that pre-existing dif-
ferences in soil microbial communities under different
land use legacies may serve as an important covariate in
cropping systems analyses.
5. ACKNOWLEDGEMENTS
We would like to thank Laura Lipps, John Hall, Lawrence Oates,
and Stephan Miramontes for the assistance with field sampling, Harry
Read for assistance with analyzing lipid biomarkers, and Ting-Li Lin
(UW-Madison Department of Statistics, CALS statistical consultant)
and Masayuki Ushio for statistical expertise. This work was funded by
the DOE Great Lakes Bioenergy Research Center (DOE BER Office of
Science DE-FC02-07ER64494).
REFERENCES
[1] Lundquist, E.J., Scow, K.M., Jackson, L.E., Uesugi, S.L.
and Johnson, C.R. (1999) Rapid response of soil micro-
bial communities from conventional, low input, and or-
ganic farming systems to a wet/dry cycle. Soil Biology
and Biochemistry, 31, 1661-1675.
doi:10.1016/S0038-0717(99)00080-2
[2] Yao, H., Bowman, D. and Shi, W. (2006) Soil microbial
community structure and diversity in a turfgrass
chronosequence: Land-use change versus turfgrass man-
agement. Applied Soil Ecology, 34, 209-218.
doi:10.1016/j.apsoil.2006.01.009
[3] Zelles, L., Rackwitz, R., Bai, Q., Beck, T. and Beese, F.
(1995) Discrimination of microbial diversity by fatty acid
profiles of phospholipids and lipopolysaccharides in dif-
ferently cultivated soils. Plant and Soil, 170, 115-122.
doi:10.1007/BF02183059
[4] Anderson, T.-H. (2003) Microbial eco-physiological
indicators to asses soil quality. Agriculture, Ecosystems
& Environment, 98, 285-293.
doi:10.1016/S0167-8809(03)00088-4
[5] Visser, S. and Parkinson, D. (1992) Soil biological crite-
ria as indicators of soil quality: Soil microorganisms.
American Journal of Alternative Agriculture , 7, 33-37.
doi:10.1017/S0889189300004434
[6] Steenwerth, K.L., Jackson, L.E., Calderon, F.J., Strom-
berg, M.R., Scow, K.M. (2002) Soil microbial commu-
nity composition and land use history in cultivated and
grassland ecosystems of coastal California. Soil Biology
and Biochemistry, 34, 1599-1611.
doi:10.1016/S0038-0717(02)00144-X
[7] Balser, T.C. and Firestone, M.K. (2005) Linking micro-
bial community composition and soil processes in a
California annual grassland and mixed-conifer forest.
Biogeochemistry, 73, 395-415.
doi:10.1007/s10533-004-0372-y
[8] Nusslein, K. and Tiedje, J.M. (1999) Soil bacterial com-
munity shift correlated with change from forest to pas-
ture vegetation in a tropical soil. Applied and Environ-
mental Microbiology, 65, 3622-3626.
[9] Kennedy, A.C., Stubbs, T.L. and Schillinger, W.F. (2004)
Soil and crop management effects on soil microbiology.
CRC Press, Boca Raton, FL.
[10] Myers, R.T., Zak, D.R., White, D.C. and Peacock, A.
(2001) Landscape-Level Patterns of Microbial Commu-
nity Composition and Substrate Use in Upland Forest
Ecosystems. Soil Science Society of America Journal, 65,
359-367. doi:10.2136/sssaj2001.652359x
[11] Grayston, S.J., Griffith, G.S., Mawdsley, J.L., Campbell,
C.D. amd Bardgett, R.D. (2001) Accounting for variabil-
ity in soil microbial communities of temperate upland
grassland ecosystems. Soil Biology and Biochemistry, 33,
533-551. doi:10.1016/S0038-0717(00)00194-2
[12] Ushio, M., Wagai, R., Balser, T.C. and Kitayama, K.
(2008) Variations in the soil microbial community com-
position of a tropical montane forest ecosystem: Does
tree species matter? Soil Biology and Biochemistry, 40,
2699-2702. doi:10.1016/j.soilbio.2008.06.023
[13] Van der Heijden, M.G.A., Bardgett, R.D. and van
Straalen, N.M. (2008) The unseen majority: soil mi-
crobes as drivers of plant diversity and productivity in
terrestrial ecosystems. Ecology Letters, 11, 296-310.
doi:10.1111/j.1461-0248.2007.01139.x
[14] Zak, D.R., Holmes, W.E., White, D.C., Peacock, A.D.
and Tilman, D. (2003) Plant diversity, soil microbial
communities, and ecosystem function: are there any links?
Ecology, 84, 2042-2050. doi:10.1890/02-0433
[15] Bartelt-Ryser, J., Joshi, J., Schmid, B., Brandl, H. and
Balser, T. (2005) Soil feedbacks of plant diversity on soil
microbial communities and subsequent plant growth.
Perspectives in Plant Ecology, Evolution and Systematics,
7, 27-49. doi:10.1016/j.ppees.2004.11.002
[16] Pankhurst, C.E., Blair, B.L., Magarey, R.C., Stirling, G.R.,
Garside, A.L. (2005) Effects of biocides and rotation
breaks on soil organisms associated with the poor early
growth of sugarcane in continuous monoculture. Plant
and Soil, 268, 255-269.
doi:10.1007/s11104-004-0287-3
[17] Robertson, G.P., Dale, V.H., Doering, O.C., Hamburg,
S.P., Melillo, J.M., Wander, M.M., Parton, W.J., Adler,
P.R., Barney, J.N., Cruse, R.M., Duke, C.S., Fearnside,