D. M. Ribeiro et al. / Agricultural Sciences 2 (2011) 78-85
Copyright © 2011 SciRes. Openly accessible at http://www.scirp.org/journal/AS/
84
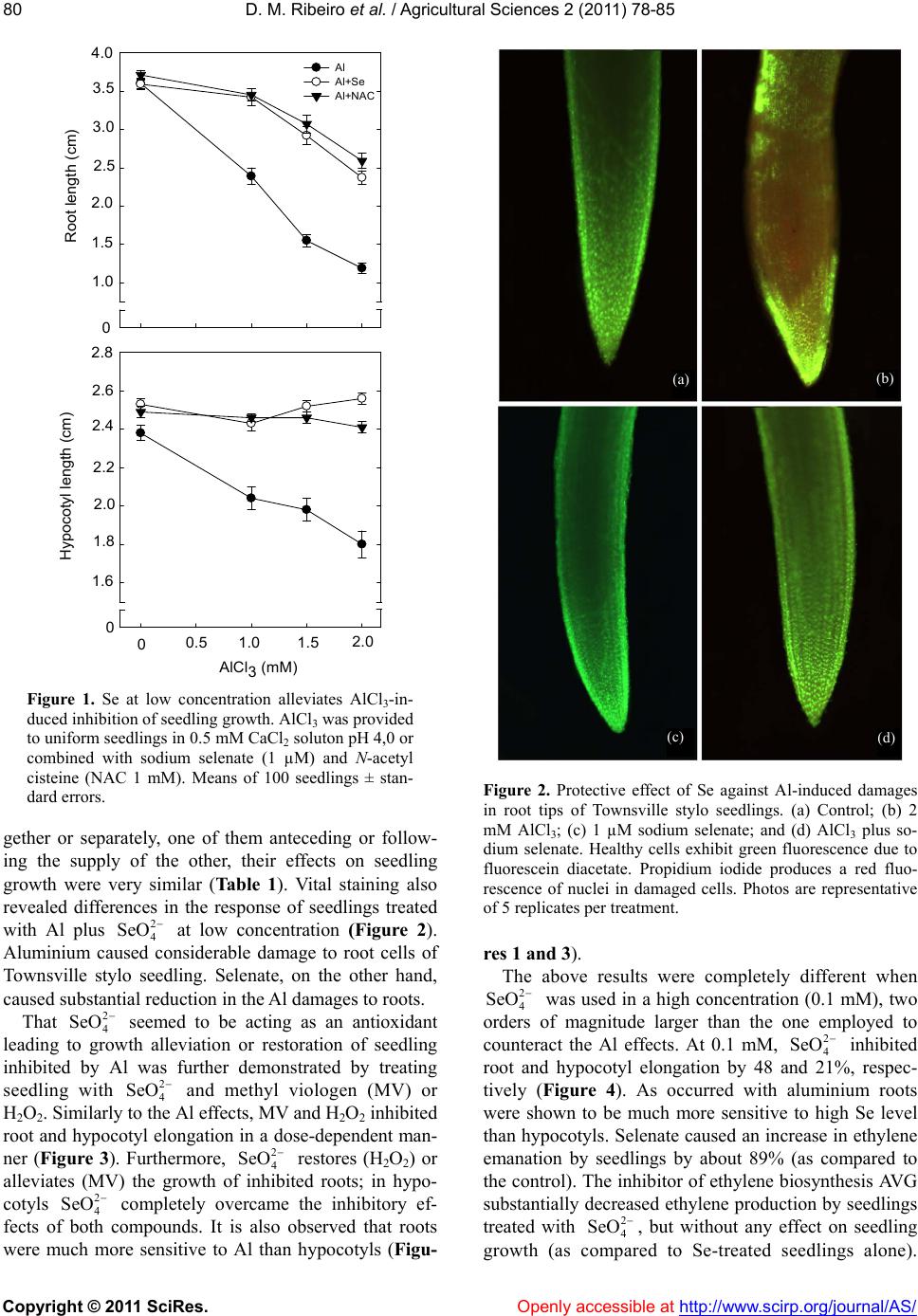
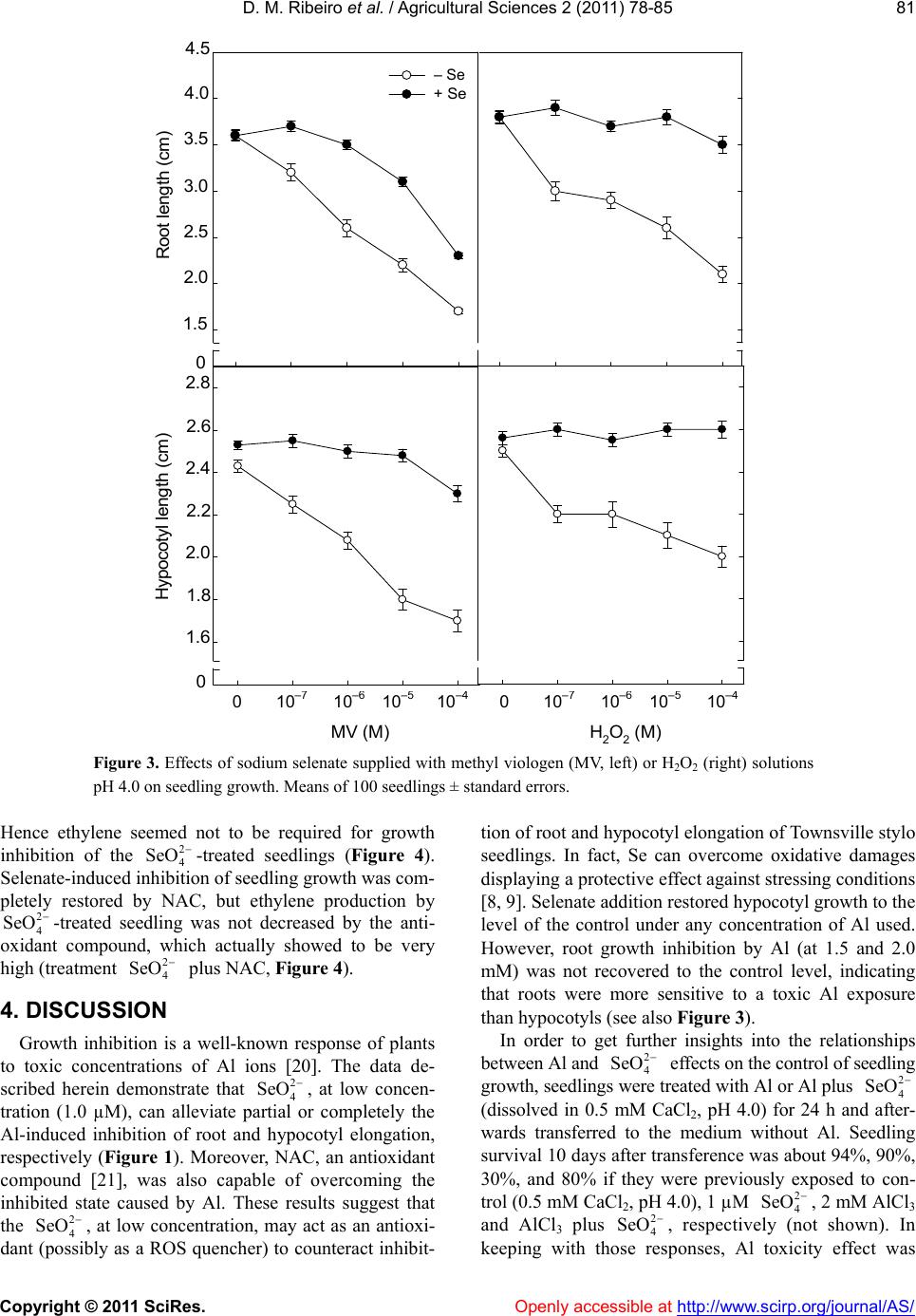
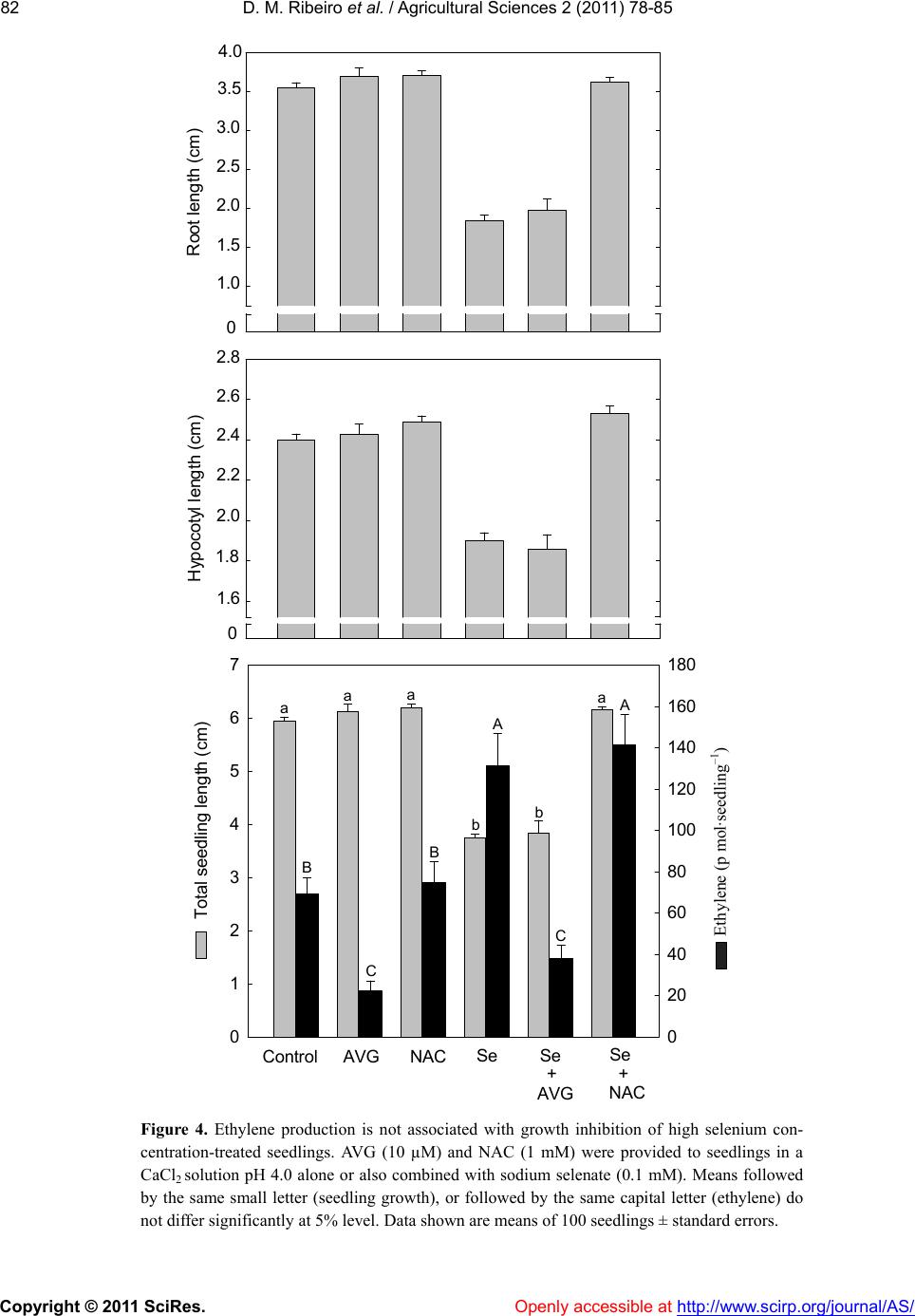
In summary, the action of on seedling growth
of Townsville stylo seedling was shown to depend on its
concentration. At low concentration, it promotes the re-
lease of the Al growth inhibition, seeming to work as a
scavenger of free radicals. At high concentrations, inhi-
bition of seedling growth is not associated with a
-dependent ethylene biosynthesis, but seems as-
sociated to the ROS generation.
2
4
SeO
2
4
SeO
5. ACKNOWLEDGEMENTS
Thanks are due to FAPEMIG (Foundation for Research Support of
Minas Gerais State) for the post-doctoral fellowship awarded to D.M.R.
and for the financial support during the conduct of this research.
REFERENCES
[1] Delhaize, E. and Ryan, P.R. (1995) Aluminum toxicity
and tolerance in plants. Plant Ph ys iol ogy, 107, 315- 321.
[2] Kochian, L.V., Hoekenga, O.A. and Pineros, M.A (2004)
How do crop plants tolerate acid soils? Mechanisms of
aluminium tolerance and phosphorous efficiency. Annual
Review of Plant Biol og y, 55, 459-493.
doi:10.1146/annurev.arplant.55.031903.141655
[3] Neill, S.J., Desikan, R. and Hancock, J.T. (2002) Hydro-
gen peroxide signalling. Current Opinion in Plant Biol-
ogy, 5, 388-395. doi:10.1016/S1369-5266(02)00282-0
[4] Oracz, K., Bouteau, H,E.M., Farrant, J. M., Cooper, K.,
Belghazi, M., Job, C. et al. (2007) ROS production and
protein oxidation as a novel mechanism for seed dor-
mancy alleviation. The Plant Journal, 50, 452-465.
d oi :1 0.1111 /j .13 65-313X.2007.03063.x
[5] Bailey-Serres, J. and Mittler, R.M. (2006) The roles of
reactive oxygen species in plant cells. Plant Physiology,
141, 331. doi:10.1104/pp.104.900191
[6] Carol, R. and Dolan, L. (2006) The role of reactive oxy-
gen species in cell growth: Lessons from root hairs.
Journal of Experimental Botany, 57, 1829-1834.
doi:10.1093/jxb/erj201
[7] Seppänen, M., Turakainen, M. and Hartikainen, H. (2003)
Selenium effects on oxidative stress in potato. Plant Sci-
ence, 165, 311-319.
doi:10.1016/S0168-9452(03)00085-2
[8] Filek, M., Keskinen, R., Hartikainen, H., Szarejko, I.,
Janiak, A., Miszalski, Z. et al. (2008) The protective role
of selenium in rape seedlings subjected to cadmium
stress. Journal of Plant Physiology, 165, 833-844.
doi:10.1016/j.jplph.2007.06.006
[9] Hartikainen, H., Xue, T. and Piironen, V. (2000) Sele-
nium as an antioxidant and pro-oxidant in ryegrass. Plant
and Soil, 225, 193-200.
doi:10.1023/A:1026512921026
[10] Mora, M., Pinilla, L., Rosas, A. and Cartes, P. (2008)
Selenium uptake and its influence on the antioxidative
system of white clover as affected by lime and phospho-
rus fertilization. Plant and Soil, 303, 139-149.
doi:10.1007/s11104-007-9494-z
[11] Brown, T.A. and Shrift, A. (1982) Selenium: Toxicity and
tolerance in higher plants. Biological Review, 57, 59-84.
d oi :1 0.1111 /j .14 69-185X.1982.tb00364.x
[12] Eustice, D.C., Kull, F.J. and Shrift, A. (1981) Selenium
toxicity: Aminoacylation and peptide bond formation
with selenomethionine. Plant Physiology, 67, 1054- 1058.
doi:10.1104/pp.67.5.1054
[13] Konze, J.R., Schilling, N. and Kende, H. (1978) En-
hancement of ethylene formation by selenoamino acids.
Plant Physiology, 62, 397-401.
doi:10.1104/pp.62.3.397
[14] Pinheiro, F.J.A., Barros, R.S., Coelho, T.G. and Souza, B.
M.L. (2008) Breaking dormancy of Stylosanthes humilis
seeds with selenium compounds. Seed Science Research,
18, 47-53. doi:10.1017/S0960258508890253
[15] Feng, R., Wei, C., Tu, S. and Sun, X. (2009) Interactive
effects of selenium and arsenic on their uptake by Pteris
vittata L. under hydroponic conditions. Environmental
and Experimental Botany, 65, 363-368.
doi:10.1016/j.envexpbot.2008.11.013
[16] Williams, R.J., Reid, R., Schultze-Kraft, R., Souza Costa,
N.M. and Thomas, B.D. (1984) Natural distribution of
Stylosanthes. In: Stace, H.M. and Edye, L.A. Eds., The
Biology and Agronomy of Stylosanthes, Academic Press,
Sydney, 73-110.
[17] Gardner, C.J. (1984) The Dynamics of Stylosanthes Pas-
tures. In: Stace, H.M. and Edye, L.A. Eds., The Biology
and Agronomy of Stylosanthes, Academic Press, Sydney,
333-357.
[18] Rangel, A.F., Mobin, M., Rao, I.M. and Horst, W.J. (2005)
Proton toxicity interferes with the screening of common
bean (Phaseolus vulgaris L.) genotypes for aluminium
resistance in nutrient solution. Journal of Plant Nutrition
and Soil Science, 168, 607-616.
doi:10.1002/jpln.200520509
[19] Jones, K.H. and Senft, J.A. (1985) An improved method
to determine cell viability by simultaneous staining with
fluorescein diacetate-propidium iodide. Journal of His-
tochemistry and Cytochemistry, 33, 77-79.
doi:10.1177/33.1.2578146
[20] Jones, D.L., Blancaflor, E.B., Kochian, L.V. and Gilroy,
S. (2008) Spatial coordination of aluminium uptake,
production of reactive oxygen species, callose production
and wall rigidification in maize root plant. Plant Cell and
Environment, 29, 1309-1318.
d oi :1 0.1111 /j .13 65-3040.2006.01509.x
[21] Lobréaux, S., Thoiron, S. and Briat, J.-F. (1995) Induc-
tion of ferritin synthesis in maize leaves by an
iron-mediated oxidative stress. The Plant Journal, 8,
443-449. doi:10.1046/j.1365-313X.1995.08030443.x
[22] Babbs, C.F., Phan, J.A. and Coolbaugh, R.C. (1989) Le-
thal hydrogen radical production in paraquat-treated
plants, Plant Physiogy, 90, 1267-1270.
doi:10.1104/pp.90.4.1267
[23] Foyer, C.H., Lelandais, M. and Kunert, K.J. (1994)
Photooxidative stress in plants. Physiologia Plantarum,
92, 696-717. doi: 10.1111 /j .1399-3054.1994.tb03042.x
[24] Xue, T., Hartikainen, H. and Piironen, V. (2001) Anti-
oxidative and growth-promoting effect of selenium on
senescing lettuce. Plant and Soil, 237, 55-61.
doi:10.1023/A:1013369804867
[25] Barros, R.S. and Freitas, A.W.P. (2001) Selenomethion-
ine as a dormancy-breaking agent in seeds of Stylosan-
thes humilis. Acta Physiologicae Plantarum, 23, 279-284.
doi:10.1007/s11738-001-0034-9