Natural Science
Vol.08 No.12(2016), Article ID:72823,17 pages
10.4236/ns.2016.812055
Optimization of Nitrogen, Phosphorus and Salt for Lipid Accumulation of Microalgae: Towards the Viability of Microalgae Biodiesel
Carolina T. Miranda1*, Daniel V. N. de Lima1, Georgia C. Atella2, Paula F. de Aguiar3, Sandra M. F. O. Azevedo1
1Biophysics Institute Carlos Chagas Filho, Federal University of Rio de Janeiro, Rio de Janeiro, RJ, Brasil
2Medical Biochemistry Institute Leopoldo De Meis, Federal University of Rio de Janeiro, Rio de Janeiro, RJ, Brasil
3Chemistry Institute, Federal University of Rio de Janeiro, Rio de Janeiro, RJ, Brasil

Copyright © 2016 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: November 15, 2016; Accepted: December 16, 2016; Published: December 19, 2016
ABSTRACT
In recent years, microalgae biodiesel has attracted expressive attention and investment, once it was considered a potential resource for energy. Although the wide use of microalgae biodiesel is restricted by its high production cost. For cost-efficient and sustainable production of biodiesel from microalgae, a proper understanding of the variables and their impacts on physiology of the strains is required. In this study, a simple factorial design 23 was used to find optimal conditions for the cultivation of Ankistrodesmus sp. and Chlamydomonas sp. in batch culture. The three components considered were nitrate, phosphate and sodium chloride, used to assess the metabolic versatility of the strains in brackish conditions. The results showed that culture medium with 0.04 g∙L−1 nitrate, 0.01 g∙L−1 phosphate and 5.0 g∙L−1 sodium chloride resulted to be the most effective condition to growth and fatty acids accumulation. Using this optimal condition, Ankistrodesmus sp. and Chlamydomonas sp. increased in 2.1 and 2.4 folds their fatty acids yield, respectively. Importantly, this protocol reduced 75% of the nitrate and phosphate concentrations of the original medium (ASM-1). Additionally, fatty acids analysis found that these strains were mainly constituted of C16-C18, in accordance with the requirements for biodiesel production. The simple factorial design applied here proved to be an important tool towards a better understanding of synergistic effects of tested factors on microalgae metabolism, and the resulting information could be used effectively to improve microalgae cultivation.
Keywords:
Optimization, Microalgae, Lipids, Ankistrodesmus, Chlamydomonas, Experimental Design

1. Introduction
Optimization of microalgae growth in cultures has been a challenge ever since culturing conditions were established by pioneering studies more than a century ago [1] [2] . Interest in this issue increased when microalgae became an alternative choice for the exploitation of several high value products including lipids, pigments, bioactive compounds and chemicals for industrial applications. According to current knowledge, microalgae biodiesel is at the forefront of the next generation of biofuel systems [3] [4] [5] . The numerous attributes of microalgae include: 1) high phototrophic productivity and thus the ability of large scale microalgae to use natural light [6] [7] ; 2) fast growth rates [8] [9] ; 3) the potential for non-cropland cultivation [10] [11] ; 4) the potential to trap greenhouse gases (CO2) and recycle waste water and nutrients [12] [13] [14] ; and 5) huge potential to convert sunlight into reduced carbon molecules, such as carbohydrates and lipids [15] . So far the production of biodiesel from microalgae has obtained significant advances in laboratory scale. However, the costs of microalgae production, low lipid yield and water demand still restrict their large scale exploitation. In this sense, the optimization of culture conditions figure as a key step to minimizing costs and to achieving desirable conditions to exploit the production of microalgae as a biodiesel feedstock [16] [17] [18] .
Microalgae growth is controlled by light, nutrients, temperature, salinity and pH. To adapt their metabolism to different environments, changes in shape, size, mobility and chlorophyll content occur to ensure the acquisition of light and nutrients [19] [20] . Lipids are fundamental components in virtually every aspect of microalgae life. Processes such as energy transfer, signal transduction, biosynthesis of macromolecules and photosynthesis are membrane bound, and highly dependent on lipid composition. Thus, lipids that maintain the physiological functions of membranes allow microalgae to re- adjust to environmental change and to tolerate severe stress. Microalgae species can accumulate lipids in the form of triacylglycerol (TAG) up to 20% - 30% of their dry cell weight, and some few species even up to 50% [21] . TAGs are constituted of three fatty acids esterified to a glycerol backbone and are one of the most concentrated forms of energy available in eukaryotic cells [22] . There is a vast range of commercial applications for these lipids, which includes biodiesel production, leading to an increased interest to improve microalgae culture techniques that ensure high levels of productivity [23] [24] [25] . Until now, the most commonly applied stress to improve TAG accumulation in microalgae has been the removal of nitrogen from culture medium [26] [27] . However, the deficiencies have been also observed to limit the growth of microalgae bring an unsatisfactory result. Other stress conditions such as phosphorus depletion, and high pH, salinity, light or temperature have been described, but quantitative data are still lacking. High concentrations of sodium chloride (up to 1.0 M) have been reported to increase intracellular accumulation of fatty acids in microalgae [28] [29] . In addition, the inhibitory effect of sodium chloride in cultures of microalgae has been exploited for contamination control that could lead to biomass losses in open ponds systems; however advances in these area are still in progress [30] [31] . Nevertheless, since culturing success generally depends on the environmental conditions of the proposed cultivation area, as well as the available infrastructure, it can be difficult to compare data reported by different authors.
Our study aimed to establish the nitrate, phosphate and sodium chloride concentrations to improve the fatty acids accumulation of two indigenous strains of microalgae, as well as to provide an effective way of saving resources without thereby compromising the performance of cultures. Additionally, microalgae biomass obtained at optimized condition was subjected to further fatty acids analysis. The option of using brackish water instead of fresh-water for algae culturing, especially in Brazilian arid and semi-arid regions, can enhance the economic potential and environmental sustainability of microalgae biofuel systems. We chose a simple factorial design combined with response surface methodology (RSM) to assess a large range of experimental concentrations in order to optimize culture conditions and avoid the defects brought by single- factor optimization. These methods have been successfully utilized in others fields, such as the chemical industry and engineering, but to our knowledge have been scarcely reported to optimization of microalgae cultures [32] [33] .
2. Material and Methods
2.1. Strains and Cultivation
Ankistrodesmus sp. (ANRF-1) and Chlamydomonas sp. (CHLRN-1) isolated from Brazilian ponds were obtained from the Laboratory of Cianobacterial Ecophysiology and Toxicology culture collection in the Biophysics Institute Carlos Chagas Filho-Federal University of Rio de Janeiro. Strains were chosen with respect to their ability to grow under high light intensities and their potential to produce lipids with biotechnological relevance. Batch cultures were growth in ASM-1 medium [34] , containing 0.003 g CaCl2・2H2O, 0.004 g SO4・7H2O, 0.002 g K2HPO4・3H2O, 0.02 g NaNO3, 0.005 g MgSO4・7H2O, 0.003 g Na2HPO4・7H2O, 0.0007 g Na2EDTA, 0.0002 g FeCl3・6H2O, 0.0001 g MnCl2・4H2O, 0.0002 g H3BO3, 0.0000001 g CuCl, 0.000002 g COCl2∙6H2O, 0.00003 g ZnCl2 and salinity 0.5 ppt per liter of ultrapure water. Cultures were sparged with constant air bubbling (0.5 L∙min−1) and incubated at 22˚C ± 2˚C with a light intensity of 1400.0 µmol photons m−2∙s−1 provided by cool white fluorescent light and a photoperiod of 12 hours. The initial concentration of both strains was 5 × 10−5 cells mL−1, corresponding to 43.3 mg biomass for Ankistrodesmus sp. and 19.0 mg for Chlamydomonas sp.
2.2. Determination of Growth, Biomass Production and Lipid Extraction
The microalgae growth was measured using a Casy Counter equipment standard for dye-free determination of cell viability by Electrical Current Exclusion (ECE). 100.0 µL of lugol-fixed samples diluted in 5.0 mL Casy Ton were gently stirred to determine cell concentrations by measuring capillary 60 µm. For biomass analysis samples of each culture was harvested on the tenth day of cultivation (at the stationary stage of cells) and then concentrated by centrifugation at 2800 × g for 15 min at 4˚C. Pellets were washed with ultrapure water and dried in a freeze dryer and stored at −20˚C until analysis. The dry weight was determined gravimetrically. For lipid extraction, biomass was pre-treated using methanol and then kept for 10 min in a sonifier cell disruptor on ice. Lipid extraction was performed using the Bligh & Dyer method [35] adapted for microalgae cells. A solution of chloroform, methanol and water (1:2:0.5 v/v) was added to the mixture and shaken for 3 hours at 160 rpm to extract the lipids. The solution was centrifuged to separate biomass of the organic phase and evaporated under flowing nitrogen gas to determine the yield of lipid extract by gravimetry.
2.3. Experimental Design
A simple 23 factorial design was used in order to optimize nitrate, phosphate and salt concentrations to improve the lipid yield of Ankistrodesmus sp. and Chlamydomonas sp. (Table 1). In addition, three experiments were performed at intermediate concentrations (level 0) for an estimate of the experimental error. The compounds were tested at three different concentrations: absence (−1), intermediate (0) and elevated (+1). These were as follows: for NaNO3 (X1) 0.00 g∙L−1 (−1), 0.04 g∙L−1 (0) and 0.08 g∙L−1 (+1), for KH2PO4/Na2HPO4∙7H2O (X2) 0.00 g∙L−1 (−1), 0.01 g∙L−1 (0) and 0.02g∙L−1 (+1), and for NaCl (X3) 0.00 g∙L−1 (−1), 5.0 g∙L−1 (0) and 10.0 g∙L−1 (+1). Except for NaCl, the levels (0) and (+) correspond to 25% and 50% of their concentrations in ASM-1 medium. Limits chosen for each variable were based on previous experiments and literature review, taking into account the operating viability. As controls, we included three replicates of the intermediate concentrations of level (0) for each compound tested. Experiments were analyzed using a mathematical model as follows:
y = b0 + (b1 × 1) + (b2 × 2) + (b3 × 3) + (b12 × 1 × 2) + (b13 × 1 × 3) + (b23 × 2 × 3) + (b123 × 1 × 2 × 3) + (b11 × 1 × 1) + (b22 × 2 × 2) + (b33 × 3 × 3)
where y is the predictive response (biomass production and lipid yield); b0 is the model intercept and b1, b2, b3, b12, b13, b23, b123 and b11, b22, b33 are the regression coefficients obtained by linear regression [36] . Were evaluated the responses of biomass productivity (mg∙L−1∙d−1); lipid productivity (mg∙L−1∙d−1) and lipid yield (percentage of biomass) (Table 1).

Table 1. Variables and factor levels of the simple factorial design.
2.4. Surface Response
The optimum response range was determined by Surface Response Methodology (RSM) [37] . Using all three variables, this methodology was applied by fixing one of the variables (sodium chloride) at different levels (concentrations), and preparing the surface with the two remaining variables assigning increments of 0.2 in the range of the nitrate and KH2PO4/Na2HPO4∙7H2O concentrations between −1 and +1 levels. The response surfaces were represented by a three-dimensional graph and significance analysis of these variables was evaluated trough Derringer Function, a multi criteria decision methodology. Data analysis was performed with Sigma Plot®version12.5.
2.5. Fatty Acid Methyl Ester Analysis (FAMEs)
Lipid samples extracted from Ankistrodesmus sp. and Chlamydomonas sp. were esterified according to Harwood [38] , and samples up to 50.0 mg were dissolved in 1.0 mL of toluene to which 2.0 mL sulfuric acid 1% (in methanol) was added. The solution was kept overnight at 50˚C, after which 1.0 mL of sodium chloride 5% was added. The fatty acids were extracted twice with 2.0 mL of hexane and dried under flowing nitrogen gas. All samples were analyzed by gas chromatography using a Shimadzu (GP2010 Plus), equipped with an electron ionization detector (EI-70 eV), aquadrupolemass analyzer and an Agilent HP Ultra 2 (5%-phenyl)-methylpolysiloxane column. Helium was used as carrier gas with a flow rate of 32.9 cms−1. The injector was set at 250˚C and temperature programming was performed as follows: the column temperature was elevated to 40˚C - 160˚C at a heating rate of 30˚C min−1, 160˚C - 233˚C at a heating rate of 1˚C min−1 and from 233˚C until 300˚C at a heating rate of 30˚C min−1 for 10 min. The interface and the ion source were kept at 240˚. The components were identified by comparing their retention times and fragmentation patterns with standard Supelco 37 Component FAME Mix-Sigma. Nonadecanoic acid (C19:0) was used as an internal standard.
3. Results and Discussion
3.1. Optimization of Nitrate, Phosphate and Sodium Chloride Concentrations
So far various studies have been carried out to demonstrate that the nitrogen and pho- sphorous concentration affect the growth and lipid accumulation contrary. To maximize cell growth, sufficient concentrations of these nutrients would be required while their starvation can increase lipid accumulation for several microalgae species [39] [40] . It is imperative that an ideal condition for microalgae culturing should favor these two aspects.
In this study, a majority of the cells inoculated was viable and in a condition to divide immediately after inoculation of twelve experiments. The exponential growth was observed to begin after first day of inoculation being zero the length of lag phase. However, nitrogen and phosphorous absence limited the exponential growth of the cultures, as showed in design points 1 and 3, respectively reflecting in a shorter duration of exponential growth phase (data not shown).The results indicate that modifications of nitrate, phosphate and sodium chloride concentrations strongly affected the biomass productivity of Ankistrodesmus sp. and Chlamydomonas sp., between 14.0 - 30.4 mg∙L−1∙d−1 and 21.6 - 85.0 mg∙L−1∙d−1, respectively (Table 2). These results are in consonance with the general findings of other authors [41] [42] . The results also showed that reduced nitrate and phosphate concentrations improved lipid accumulation in the tested strains. Lipid productivity of Ankistrodesmus sp. and Chlamydomonas sp. varied between the twelve experimental conditions, ranging from 3.0 to 8.0 mg∙L−1∙d−1 and from 10.3 to 16.5 mg∙L−1∙d−1, respectively. Significant enhancements of lipid yield, up to 20% of biomass dry weight for both strains were obtained in most of the conditions tested except in experiments 2 and 4 for Ankistrodesmus sp. and 2, 4, 6, and 8 for Chlamydomonas sp., all of which had the highest nitrate concentrations. Reduced nitrate and phosphate concentrations when combined with added sodium chloride also affected positively the lipid accumulation of Chlamydomonas sp. (experiments 6 to 8 and level 0), even with a slight decrease of biomass productivity, in accordance with results reported by other researchers [31] [43] (Table 2).
The Derringer function (D) in Figure 1 illustrates the statistical analysis of the results. In this figure, the Y axis corresponds to the design points, the X axis corresponds to the desirability degree ranging from 0 (undesirable) to 1 (most desirable) and the bar
Table 2. Matrix of the experimental design with experimental values for biomass productivity, lipid productivity and lipid yield (% of biomass dry weight) on the 10th day of cultivation.
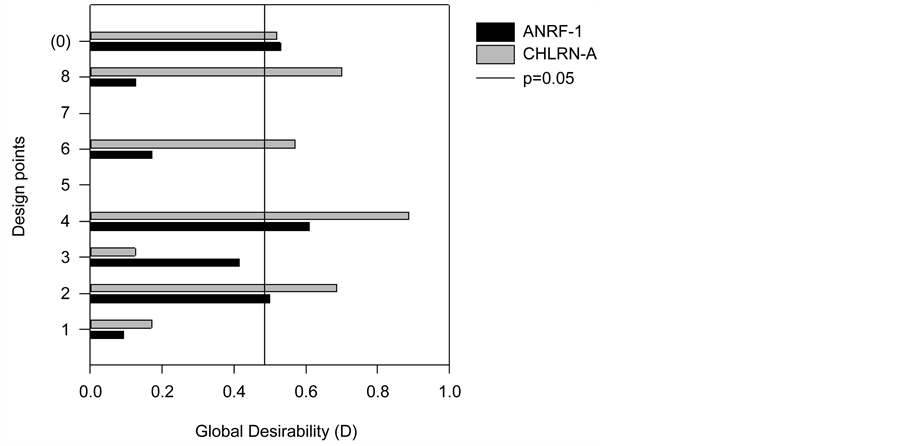
Figure 1. Derringer function of biomass and lipid productivity from Ankistrodesmus sp. (ANRF-1) and Chlamydomonas sp. (CHLRN-1) for each design point experimented.
length corresponds proportionally to the significance of each factor and its association at the levels tested (−1), (0) and (+1) on the variables biomass and lipid productivity, simultaneously. Values that exceed the vertical line are statistically significant at the 95% confidence level. The results indicated that the three factors as their interactions were significant for both biomass and lipid productivity of Ankistrodesmus sp. and Chlamydomonas sp., at the 95% confidence level. According to this statistical analysis, cell productivity and lipid yield are attained by concentrations of 0.08 g∙L−1 NaNO3, 0.02 g∙L−1 KH2PO4/Na2HPO4 (design point 4) at the 95% confidence level. The interaction between NaCl with NaNO3 and KH2PO4/Na2HPO4 is also significant (at the 95% confidence level) when all factors were at mid-level (design point 0).
3.2. Surface Response Analysis
The surface response analysis (Figure 2(a)) showed that maximal biomass productivity for Ankistrodesmus sp. (34.0 mg∙L−1∙d−1) was achieved with concentrations of nitrate at mid-level (0.04 g∙L−1), phosphate at the +1 level (0.02 g∙L−1) and sodium chloride at the −1 level (0.00 g∙L−1). For Chlamydomonas sp., maximum productivity (90.0 mg∙L−1∙d−1) was produced when nitrate and phosphate were set at the +1 level (0.08 and 0.02 g∙L−1, respectively), independent of the sodium chloride concentration (Figure 3(a), Figure 3(c) and Figure 3(e)). In addition, was verified that changes in the order of 0.5 of the phosphate concentration decreasing from the +1 level (0.02 g∙L−1) to the −1 level (0.00 g∙L−1) had an important impact on biomass productivity of both strains, leading to a significant reduction on biomass production. The highest lipid productivity for Ankistrodesmus sp. (9.0 mg∙L−1∙d−1) was achieved with concentrations of nitrate at mid-level (0.04 g∙L−1), phosphate at the +1 level (0.02 g∙L−1), sodium chloride at mid-level (5.0

Figure 2. 3D response surface and contour plots of nitrate, phosphate and sodium chloride concentrations levels on biomass production (a) (c) and (e) and lipid yield (b) (d) and (f) of Ankistrodesmus sp. when NaCl was fixed at level (−1), (0) and (+1), respectively.

Figure 3. 3D response surface and contour plots of nitrate, phosphate and sodium chloride concentrations levels on biomass production (a) (c) and (e) and lipid yield (b) (d) and (f) of Chlamydomonas sp. when NaCl was fixed at level (−1), (0) and (+1), respectively.
g∙L−1) and at the +1 level (10.0 g∙L−1), as shown in (Figure 2(d) and (Figure 2(f)). Lipid yield in Chlamydomonas sp. (Figure 3(b)) was higher when nitrate, phosphate and sodium chloride concentrations were kept at the −1 level. A combination of nitrate at the −1 level, phosphate at the +1 level and sodium chloride at the −1 level resulted in a highly satisfactory lipid yield. However, to achieve this outcome, the concentrations must be rigorously controlled since small deviations may result in a significant decrease of biomass and lipid production. The most robust surface with high yields of biomass and lipids was obtained at intermediate levels (0) for nitrate (0.04 g∙L−1), phosphate (0.01 g∙L−1) and sodium chloride (5.0 g∙L−1) and can therefore be considered as better conditions for both strains.
The accuracy of these protocol was verified through experiments repetition using the optimized conditions described above (level 0), as well as ASM-1 medium (control) in triplicate. The results of both treatments (optimized condition and control) were very similar, confirming the validity of the model and protocol used in this study. After a 10 day growth period in optimized medium, we observed a strong increase of lipid content (126%) in Chlamydomonas sp. compared with lipid production in ASM-1 medium (Figure 4(b)). Chlamydomonas was not considered an oleaginous alga until recent studies indicated that they can accumulate considerable amounts of fatty acids under appropriate conditions [44] [45] [46] . In our study, Chlamydomonas sp. (CHLRN-1) achieved a lipid productivity of 15.0 mg∙L−1∙d−1 (24% of biomass) of which 68% were fatty acids, higher than reported in other relevant studies with indigenous strains of Chlamydomonas [36] [46] [47] . The lipid content of Ankistrodesmus sp. (ANRF-1) increased by 36% in the optimized medium (Figure 4(a)), with a lipid productivity of 8.4 mg∙L−1∙d−1 rich in fatty acids (13% of lipid extract) favorable for biodiesel production. Compared with Chlamydomonas sp., the Ankistrodesmus strain had a slower growth rate resulting in a biomass yield that could be considered a limiting factor for industrial purposes. However, in addition to the key importance of volumetric biomass and lipid yield, other factors such as ease of cultivation, harvesting, and prevailing environmental
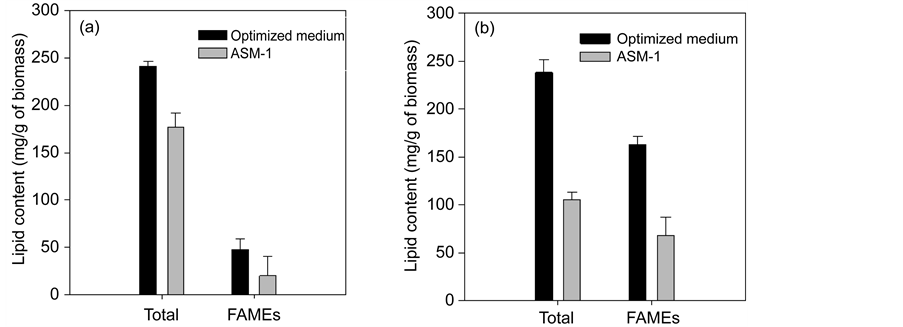
Figure 4. Differences between optimized medium and ASM-1 in the lipid content and FAMEs of Ankistrodesmus sp. (a) and Chlamydomonas sp. (b). Data points are means of three independent experiments and error bars show SD.
conditions also affect the final choice of microalgae strains as feedstock for biodiesel production [47] . While cell growth was reduced with the reduction of nitrogen and phosphorus concentrations, the successful increase of lipid accumulation under the same condition may counterbalance the losses in biomass productivity. It is relevant from a biotechnological point of view, which must also consider the economic cost of chemicals for industrial cultivation of microalgae (Figure 3).
3.3. Fatty Acid Methyl Esters (FAMEs)
The degree of saturation/unsaturation of fatty acids determines the quality of biodiesel. Therefore, we examined the fatty acid profiles produced from cultures of Ankistrodesmus sp. and Chlamydomonas sp. under operating conditions determined in this study. Fatty acids were identified and quantified by GC/MS following extractive methylation to fatty acid methyl esters (FAMEs). The total content of FAMEs for Ankistrodesmus sp. was 47.4 mg∙g−1 of the produced biomass in the optimized medium, and did not differ significantly from that obtained in ASM-1 medium(19.9 mg∙g−1, Figure 4(a)). For Chlamydomonas sp. the total content of FAMEs was significantly higher (at 99% of confidence level) in the optimized medium than in ASM-1 (162.7 mg∙g−1 and 68.1 mg∙g−1 of biomass produced, respectively, Figure 4(b)).
The reduction of nutrients or the addition of sodium chloride did not trigger variations of FAME profiles in the two tested microalgae, with the exception of C18:5 in Chlamydomonas sp., which increased significantly in comparison to the control condition, explaining the distinct increase of FAMEs overall. Ankistrodesmus sp. showed high concentrations of unsaturated fatty acids with therapeutic properties (Figure 5(a) and Figure 5(b)). The predominant fatty acids were: C18:3n3 (alpha-linolenic acid), C18:1n9c (oleic acid) and C18:2n6c (linoleic acid), represent ingomega 3, 6 and 9, respectively (Figure 5(c)). In accordance to the literature [48] [49] [50] , C16:0 (palmitic) was the most representative saturated fatty acid in both microalgal strains, and its concentration did not significantly change under the various concentrations of nutrients tested. This is not surprising since palmitic acid is a major component of most microalgal classes, while the profiles of unsaturated fatty acids differ among algal groups [51] [52] . Chlamydomonas sp. (Figure 6(a) and Figure 6(b)) produced large amounts of C18:1n9c and C18:3n3 when cultivated in optimized medium. We observed a larger variety of saturated fatty acids in this strain, including C15:0 (pentadecanoic acid), C17:0 (hepatdecanoic acid), C:20 (arachidic acid) and C:22 (behenic acid) (Figure 6(c)). A similar variety is characteristic for tallow or palm oils, which are rich in saturated fatty acids and used to produce biodiesel in Brazil (Figure 5).
4. Conclusion
The simple factorial design associated with response surface methodology was found to be effective in optimizing the concentrations of nitrate and phosphate for an efficient use of chemicals in microalgae cultivation. In order to increase lipid production nitrate and phosphate concentrations of the ASM-1 medium can be reduced by 75%, reducing

Figure 5. GC chromatogram of representative FAMEs from microalgae Ankistrodesmus sp. in optimized medium (a) and ASM-1 (b) after 10 days of cultivation. Numerals above peaks show the retention time of appropriate peak in minutes. (c) FAMEs content in lipid extract in optimized medium (black columns) and ASM-1 (gray columns). *The position of the double bond was not identified, **correspond to 16:3n3Δ4,7,10 and ***16:3n3Δ7,10,13. Data points are means of three independent experiments and error bars show SD.

Figure 6. GC chromatogram of representative FAMEs from microalgae Chlamydomonas sp. in optimized medium (a) and ASM-1 (b) after 10 days of cultivation. Numerals above peaks show the retention time of appropriate peak in minutes. (c) FAMEs content in lipid extract in optimized medium (black columns) and ASM-1 (gray columns). *The position of the double bond was not identified, **correspond to 16:3n3Δ4,7,10 and ***16:3n3Δ7,10,13. Data points are means of three independent experiments and error bars show SD.
biomass losses and improving the control of operation conditions. Our results conclude that Ankistrodesmus sp. (ANRF-1) and Chlamydomonas sp. (CHLRN-1) could be suitable for biodiesel production due to a high proportion of C16-C18 fatty acids, attending the basic requirements for biodiesel production. In addition, the ability to grow in a medium with added sodium chloride also contributes to the significant potential of these strains for biodiesel production, demonstrating that fresh-water is not required for further scaling up to semi-pilot scale. This is particularly relevant in a global scenario where fresh water supplies must be protected to maintain environmental and public health.
Acknowledgements
The authors would like to thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for providing the scholarships. Thanks to Centro de Pesquisas e Desenvolvimento Leopoldo A. Miguez de Mello (CENPES) for graciously providing the microalgae Chlamydomonas sp. (CHLRN-1 strain) and for the financial support. We express our gratitude to Mileane S. Busch for the technical assistance on GC-MS analysis.
Cite this paper
Miranda, C.T., de Lima, D.V.N., Atella, G.C., de Aguiar, P.F. and Azevedo, S.M.F.O. (2016) Optimization of Nitrogen, Phosphorus and Salt for Lipid Accumulation of Microalgae: Towards the Viability of Microalgae Biodiesel. Natural Science, 8, 557-573. http://dx.doi.org/10.4236/ns.2016.812055
References
- 1. Bold, H.C. (1942) The Cultivation of Algae. Botanical Review, 8, 69-138.
https://doi.org/10.1007/BF02879474 - 2. Pringsheim, E.G. (1946) The Biphasic or Soil-Water Culture Method for Growing Algae and Flagellata. Journal of Ecology, 33, 193-204. http://www.jstor.org/stable/2256465
https://doi.org/10.2307/2256465 - 3. Ma, F. and Hanna, M.A. (1999) Biodiesel Production: A Review. Bioresource Technology, 70, 1-15.
https://doi.org/10.1016/S0960-8524(99)00025-5 - 4. Huang, G., Chen, F., Wei, D., Zhang, X. and Chen, G. (2010) Biodiesel Production by Microalgal Biotechnology. Applied Energy, 87, 38-46.
https://doi.org/10.1016/j.apenergy.2009.06.016 - 5. Brennan, L. and Owende, P. (2010) Biofuels from Microalgae—A Review of Technologies for Production, Processing, and Extractions of Biofuels and Co-Products. Renewable & Sustainable Energy Reviews, 14, 557-577.
https://doi.org/10.1016/j.rser.2009.10.009 - 6. Koller, M., Salerno, A., Tuffner, P., Koinigg, M., Bochzelt, H., Schober, S., et al. (2012) Characteristics and Potential of Micro Algal Cultivation Strategies: A Review. Journal of Cleaner Production, 37, 377-388.
https://doi.org/10.1016/j.jclepro.2012.07.044 - 7. Sforza, E., Bertucco, A., Morosinotto, T. and Giacometti, G.M. (2012) Photobioreactors for Microalgal Growth and Oil Production with Nannochloropsis salina: From Lab-Scale Experiments to Large-Scale Design. Chemical Engineering Research and Design, 90, 1151-1158.
https://doi.org/10.1016/j.cherd.2011.12.002 - 8. Venkata Mohan, S. and Devi, M.P. (2014) Salinity Stress Induced Lipid Synthesis to Harness Biodiesel during Dual Mode Cultivation of Mixotrophic Microalgae. Bioresource Technology, 165, 88-94.
https://doi.org/10.1016/j.biortech.2014.02.103 - 9. Passarge, J., Hol, S., Escher, M. and Huisman, J. (2006) Competition for Nutrients and Light: Stable Coexistence, Alternative Stable States, or Competitive Exclusion? Ecological Monographs, 76, 57-72.
https://doi.org/10.1890/04-1824 - 10. Wijffels, R.H., Kruse, O. and Hellingwerf, K.J. (2013) Potential of Industrial Biotechnology with Cyanobacteria and Eukaryotic Microalgae. Current Opinion in Biotechnology, 24, 405-413.
https://doi.org/10.1016/j.copbio.2013.04.004 - 11. Smith, V.H., Sturm, B.S.M., de Noyelles, F.J. and Billings, S. (2010) The Ecology of Algal Biodiesel Production. Trends in Ecology & Evolution, 25, 301-309.
https://doi.org/10.1016/j.tree.2009.11.007 - 12. Chen, G., Zhao, L. and Qi, Y. (2015) Enhancing the Productivity of Microalgae Cultivated in Wastewater toward Biofuel Production: A Critical Review. Applied Energy, 137, 282-291.
https://doi.org/10.1016/j.apenergy.2014.10.032 - 13. Larsdotter, K. (2006) Wastewater Treatment with Microalgae—A Literature Review. Vatten, 62, 31-38.
- 14. Quiroz Arita, C.E., Peebles, C. and Bradley, T.H. (2015) Scalability of Combining Microalgae-Based Biofuels with Wastewater Facilities: A Review. Algal Research, 9, 160-169.
https://doi.org/10.1016/j.algal.2015.03.001 - 15. Thompson, G. (1996) Lipids and Membrane Function in Green Algae. Biochimica et Biophysica Acta, 1302, 17-45.
https://doi.org/10.1016/0005-2760(96)00045-8 - 16. Li, Z., Yuan, H., Yang, J. and Li, B. (2011) Optimization of the Biomass Production of Oil Algae Chlorella minutissima UTEX2341. Bioresource Technology, 102, 9128-9134.
https://doi.org/10.1016/j.biortech.2011.07.004 - 17. Work, V.H., D’Adamo, S., Radakovits, R., Jinkerson, R.E. and Posewitz, M.C. (2012) Improving Photosynthesis and Metabolic Networks for the Competitive Production of Phototroph-Derived Biofuels. Current Opinion in Biotechnology, 23, 290-297.
https://doi.org/10.1016/j.copbio.2011.11.022 - 18. Picardo, M.C., De Medeiros, J.L., Monteiro, J.G.M., Chaloub, R.M., Giordano, M. and De Queiroz Fernandes Araújo, O. (2013) A Methodology for Screening of Microalgae as a Decision Making Tool for Energy and Green Chemical Process Applications. Clean Technologies and Environmental Policy, 15, 275-291.
https://doi.org/10.1007/s10098-012-0508-z - 19. Reynolds, C.S. (2006) The Ecology of Phytoplankton. Cambridge University Press, Cambridge.
https://doi.org/10.1017/CBO9780511542145 - 20. Sommer, U. (1999) A Comment on the Proper Use of Nutrient Ratios in Microalgal Ecology. Archiv fur Hydrobiologie, 146, 55-64.
https://doi.org/10.1127/archiv-hydrobiol/146/1999/55 - 21. Miranda, C.T., Pinto, R.F., De Lima, D.V.N., Viegas, C.V., Costa, S.M. and Azevedo, S.M.F.O. (2015) Microalgae Lipid and Biodiesel Production: A Brazilian Challenge. American Journal of Plant Sciences, 6, 2522-2533.
https://doi.org/10.4236/ajps.2015.615254 - 22. Gurr, M., Harwood, J. and Frayn, K. (2002) Lipid Biochemistry: An Introduction. Blackwell Science, Hoboken.
https://doi.org/10.1002/9780470774366 - 23. Mata, T.M., Martins, A. and Caetano, N.S. (2010) Microalgae for Biodiesel Production and Other Applications: A Review. Renewable & Sustainable Energy Reviews, 14, 217-232.
https://doi.org/10.1016/j.rser.2009.07.020 - 24. Spolaore, P., Joannis-Cassan, C., Duran, E. and Isambert, A. (2006) Commercial Applications of Microalgae. Journal of Bioscience and Bioengineering, 101, 87-96.
https://doi.org/10.1263/jbb.101.87 - 25. Meng, X., Yang, J., Xu, X., Zhang, L., Nie, Q. and Xian, M. (2009) Biodiesel Production from Oleaginous Microorganisms. Renewable Energy, 34, 1-5.
https://doi.org/10.1016/j.renene.2008.04.014 - 26. Rezanka, T., Lukavsky, J., Nedbalov, L. and Sigler, K. (2011) Effect of Nitrogen and Phosphorus Starvation on the Polyunsaturated Triacylglycerol Composition, Including Positional Isomer Distribution, in the alga Trachydiscus minutus. Phytochemistry, 72, 2342- 2351. https://doi.org/10.1016/j.phytochem.2011.08.017
- 27. Gao, Y., Yang, M. and Wang, C. (2013) Nutrient Deprivation Enhances Lipid Content in Marine Microalgae. Bioresource Technology, 147, 484-491.
https://doi.org/10.1016/j.biortech.2013.08.066 - 28. Siaut, M., Cuiné, S., Cagnon, C., Fessler, B., Nguyen, M., Carrier, P., et al. (2011) Oil Accumulation in the Model Green Alga Chlamydomonas reinhardtii: Characterization, Variability between Common Laboratory Strains and Relationship with Starch Reserves. BMC Biotechnology, 11, 7.
https://doi.org/10.1186/1472-6750-11-7 - 29. Takagi Karseno, M. and Yoshida, T. (2006) Effect of Salt Concentration on Intracellular Accu-mulation of Lipids and Triacylglyceride in Marine Microalgae Dunaliella Cells. Journal of Bioscience and Bioengineering, 101, 223-226.
https://doi.org/10.1263/jbb.101.223 - 30. Ho, S.-H., Ye, X., Hasunuma, T., Chang, J.-S. and Kondo, A. (2014) Perspectives on Engineering Strategies for Improving Biofuel Production from Microalgae—A Critical Review. Biotechnology Advances, 32, 1448-1459.
https://doi.org/10.1016/j.biotechadv.2014.09.002 - 31. Rawat, I., Ranjith Kumar, R., Mutanda, T. and Bux, F. (2013) Biodiesel from Microalgae: A Critical Evaluation from Laboratory to Large Scale Production. Applied Energy, 103, 444-467.
https://doi.org/10.1016/j.apenergy.2012.10.004 - 32. Nicolaisen, O., Cuny, M. and Bolla, S. (2014) Factorial Experimental Designs as Tools to Optimize Rearing Conditions of Fish Larvae. Aquaculture, 422-423, 253-260.
https://doi.org/10.1016/j.aquaculture.2013.12.018 - 33. Chen, J.-J., Li, Y.-R. and Lai, W.-L. (2014) Application of Experimental Design Methodology for Optimization of Biofuel Production from Microalgae. Biomass and Bioenergy, 64, 11-19.
https://doi.org/10.1016/j.biombioe.2014.03.056 - 34. Kangatharalingam, N. (1993) Isolation and Verification of Anatoxin-A Producing Clones of Anabaena flosaquae (Lyngb.) de Breb. from a Eutrophic Lake. FEMS Microbiology Ecology, 12, 127-130.
https://doi.org/10.1111/j.1574-6941.1993.tb00024.x - 35. Bligh, E.G. and Dyer, W.J. (1959) A Rapid Method of Total Lipid Extraction and Purification. Canadian Journal of Biochemistry and Physiology, 37, 911-917.
https://doi.org/10.1139/o59-099 - 36. Montgomery, D. (2009) Introduction to Statistical Quality Control. John Wiley & Sons, Hoboken.
- 37. Rambali, B., Baert, L. and Massart, D.L. (2001) Using Experimental Design to Optimize the Process Parameters in Fluidized Bed Granulation on a Semi-Full Scale. International Journal of Pharmaceutics, 220, 149-160.
https://doi.org/10.1016/S0378-5173(01)00658-5 - 38. Harwood, J. (1989) Gas Chromatography and Lipids: A Practical Guide. Phytochemistry, 28, 3251-3252.
https://doi.org/10.1016/0031-9422(89)80324-3 - 39. Xiong, W., Li, X., Xiang, J. and Wu, Q. (2008) High-Density Fermentation of Microalga Chlorella protothecoides in Bioreactor for Microbio-Diesel Production. Applied Microbiology and Biotechnology, 78, 29-36.
https://doi.org/10.1007/s00253-007-1285-1 - 40. Su, C.H., Chien, L.J., Gomes, J., Lin, Y.S., Yu, Y.K. and Liou, J.S., et al. (2011) Factors Affecting Lipid Accumulation by Nannochloropsis oculata in a Two-Stage Cultivation Process. Journal of Applied Phycology, 23, 903-908.
https://doi.org/10.1007/s10811-010-9609-4 - 41. Widjaja, A., Chien, C.C. and Ju, Y.H. (2009) Study of Increasing Lipid Production from Fresh Water Microalgae Chlorella vulgaris. Journal of the Taiwan Institute of Chemical Engineers, 40, 13-20.
https://doi.org/10.1016/j.jtice.2008.07.007 - 42. Lv, J.M., Cheng, L.H., Xu, X.H., Zhang, L. and Chen, H.L. (2010) Enhanced Lipid Production of Chlorella vulgaris by Adjustment of Cultivation Conditions. Bioresource Technology, 101, 6797-6804.
https://doi.org/10.1016/j.biortech.2010.03.120 - 43. Sharma, K.K., Schuhmann, H. and Schenk, P.M. (2012) High Lipid Induction in Microal-gae for Biodiesel Production. Energies, 5, 1532-1553.
https://doi.org/10.3390/en5051532 - 44. Morowvat, M.H., Rasoul-Amini, S. and Ghasemi, Y. (2010) Chlamydomonas as a “New” Organism for Biodiesel Production. Bioresource Technology, 101, 2059-2062.
https://doi.org/10.1016/j.biortech.2009.11.032 - 45. Merchant, S.S., Kropat, J., Liu, B., Shaw, J. and Warakanont, J. (2012) TAG, You’re It! Chlamydomonas as a Reference Organism for Understanding Algal Triacylglycerol Accumulation. Current Opinion in Biotechnolo-gy, 23, 352-363.
https://doi.org/10.1016/j.copbio.2011.12.001 - 46. Fan, J., Andre, C. and Xu, C. (2011) A Chloroplast Pathway for the de Novo Biosynthesis of Triacylglycerol in Chlamydomonas reinhardtii. FEBS Letters, 585, 1985-1991.
https://doi.org/10.1016/j.febslet.2011.05.018 - 47. Talebi, A.F., Mohtashami, S.K., Tabatabaei, M., Tohidfar, M., Bagheri, A., Zeinalabedini, M., et al. (2013) Fatty Acids Profiling: A Selective Criterion for Screening Microalgae Strains for Biodiesel Production. Algal Research, 2, 258-267.
https://doi.org/10.1016/j.algal.2013.04.003 - 48. Díaz, G.C., Cruz, Y.R., Fortes, M.M. and Aranda, D.A.G. (2014) Primary Separation of Antioxidants (Unsaponifiables) the Wet Biomass Microalgae Chlamydomonas sp. and Production of the Bio-diesel. Natural Science, 6, 1210-1218.
https://doi.org/10.4236/ns.2014.615108 - 49. Piorreck, M. and Pohl, P. (1984) Formation of Biomass, Total Protein, Chlorophylls, Lipids and Fatty Acids in Green and Blue-Green Algae during One Growth Phase. Phytochemistry, 23, 217-223.
https://doi.org/10.1016/S0031-9422(00)80305-2 - 50. Converti, A., Casazza, A., Ortiz, E.Y., Perego, P. and Del Borghi, M. (2009) Effect of Temperature and Nitrogen Concentration on the Growth and Lipid Content of Nannochloropsis oculata and Chlorella vulgaris for Biodiesel Production. Chemical Engineering and Processing: Process Intensification, 48, 1146-1151.
https://doi.org/10.1016/j.cep.2009.03.006 - 51. Reuss, N. and Poulsen, L.K. (2002) Evaluation of Fatty Acids as Biomarkers for a Natural Plankton Community. A Field Study of a Spring Bloom and a Post-Bloom Period off West Greenland. Marine Biology, 141, 423-434.
https://doi.org/10.1007/s00227-002-0841-6 - 52. Lang, I., Hodac, L., Friedl, T. and Feussner, I. (2011) Fatty Acid Profiles and Their Distribution Patterns in Microalgae: A Comprehensive Analysis of More than 2000 Strains from the SAG Culture Collection. BMC Plant Biology, 11, 124.
https://doi.org/10.1186/1471-2229-11-124