Natural Science
Vol.4 No.9(2012), Article ID:22462,9 pages DOI:10.4236/ns.2012.49097
Skeletal extension rate of the reef building coral Porites species from Aqaba and their environmental variables
![]()
Marine Science Station, The University of Jordan, Aqaba, Jordan; s.rousan@ju.edu.jo
Received 10 July 2012; revised 8 August 2012; accepted 21 August 2012
Keywords: Coral Growth; Porites; Skeletal Extension Rate; Gulf of Aqaba; Red Sea
ABSTRACT
Annual skeletal extension rates of the scleraactinian corals Porites species were investigated in 32 colonies from the northern Gulf of Aqaba fringing reef at various depths (1 - 42 m). All corals reveal clear and regular skeletal density banding patterns. Results showed that the high-density annual growth bands were formed during winter and the low-density annual growth bands during summer. The mean annual extension rates of the studied corals reveal a large inter-colony variability with values ranged between 2.36 to 20.0 mm/year. While a general trend of decreasing coral extension rate with depth was observed and best explained by a simple exponential model, the rates clustered into two groups: 10.86 ± 2.54 mm/year in water depths less than 10 m, and 5.23 ± 1.99 mm/year below 12 m. Light intensity seems to be the primary environmental factor responsible for decreasing coral extension rate with depth since the effect of other environmental parameters could be neglected from the Gulf of Aqaba. Time series record of the mean annual coral extension rate showed a slight increasing linear trend which could be linked to increase seawater temperature over the period of time represented.
1. INTRODUCTION
Massive stony corals from modern and fossil reefs of the tropical and sub-tropical oceans provide an important archive of past climate and ocean variability [1-6]. These corals build skeletons of aragonite (CaCO3) and grow at rates of millimetres to centimetres per year.
Due to the enormous environmental information that can be recognized in coral skeletal materials, the understanding of coral growth records have been increased over the last decades [6-11]. The annual density bands in massive corals that are produced in the skeleton during growth [12,13] have been widely used to achieve this purpose as it provides long-term dated histories (chronology) of coral growth and calcification [14-16].
The coral growth is an important parameter in assessing the impact of natural or anthropogenic climate and environmental changes [15,17]. It can reflect several parameters such as temperature, nutrient and food availability, water transparency and sediment input [17-19]. The environmental and ecological factors that could influence coral growth are almost unlimited in number (reviewed by [14] and [17]). Some of these factors could decrease the growth rates such as increasing latitude [20]; increasing the water motion [21], decrease nutrient and light availability [22], increasing water depth [8], increase turbidity and sedimentation [11,23] and decreasing water temperature [15,16].
In the Gulf of Aqaba, coral reef communities represent the northern limit (29˚32′N) for reef corals in the western Indo-Pacific region [24,25]. They are highly diverse and mostly of the fringing type, because the Red Sea and the Gulf of Aqaba are devoid of a true continental shelf and the offshore profiles are very steep, therefore, the reefs are narrow and they closely follow the shorelines [26].
In this study, I have investigated the annual skeletal extension rate obtained from 181 annual growth bands in Porites corals from various depths in Aqaba reef. The Porities coral spp. was chosen because it is widely distributed and represents a major reef building scleractinia at the reef throughout the Gulf of Aqaba and Red Sea at all depths, it also shows a clear annual growth bands and grows continuously at high rates. The environmental variables that may affect coral extension rates from the study area were also discussed.
2. MATERIALS AND METHODS
2.1. Site Description
The study area is located at the northern end of the Gulf of Aqaba (Figure 1), which is the northward extension of the desert-enclosed Red Sea. The Gulf is a semiisolated basin separated from the Red Sea proper by the Straits of Tiran, which are about 240 m deep. The maximum depth of the Gulf is 1830 m; its 180 km long and 5 - 26 km wide.
The Gulf is located within the very warm portion of the Sahara bio-climatic zone. The climate is arid with high evaporation (~400 cm/year) and negligible precipitation (~2.2 cm/year) and runoff [27]. The mean sea surface temperatures (SSTs) are 23.5˚C and mean salinity values in the upper waters are 40.4‰ - 40.6‰ [28].
Extremely oligotrophic conditions are prevailing in the Gulf due to the arid climate and because it receives its waters from the nutrient-depleted Red Sea surface waters through the Straits of Tiran [27]. The deep light penetration and high transparency due to low amount of resuspended materials and fresh water flux [29] results in extending the depth limit of massive hermatypic corals such as Porites down to 40 - 50 m [30,31].
2.2. Coral Samples
All investigated corals in this study were collected from a depth transect from the reef complex in front of the Marine Science Station in Aqaba (29˚27′N, 34˚58′E), (Figure 1), except the corals AQB-Big, AQB-18H, AQB-ph. The transect was laid out on the reef between the reef flat and the deep fore-reef down to 42 m. Scleractinian corals were distributed along this transect at all depths.
A total of 32 columns of Porites spp. colonies (Table 1) were collected along this transect from different depths between 1 and 42 m and at different time periods (April 1999, June 2000, May 2005 and June 2010). Colonies were harvested by scuba diving by removal of hemispherical-shaped colonies (5 - 10 cm in diameter) underwater using hammer and chisel. After sampling, the corals were cleaned under high-pressure tap water to remove the residual organic matter and then dried under the sun.
The corals were sectioned longitudinally into slabs of 4 mm thickness parallel to the axis of maximum growth. The slabs were cleaned and X-rayed using industrial X-ray machine. Black-and-white positive prints of the x-radiographs were obtained for extension rate measurement.
The annual skeletal extension rates of all corals were directly measured along the major (vertical) growth axes (Figure 2) from the positive prints of the x-radiographs as the distance between the top edges of low-density bands [8]. Each couplet of high-/low-density bands represents an annual growth increment. As a double check for this method, the annual extension rates of corals were calculated from the seasonal cycle of δ18O (or Sr/Ca) as the distance from the maximum δ18O (Sr/Ca) value (which corresponds to the seasonal temperature minimum) in a given year to the maximum value in the fol-
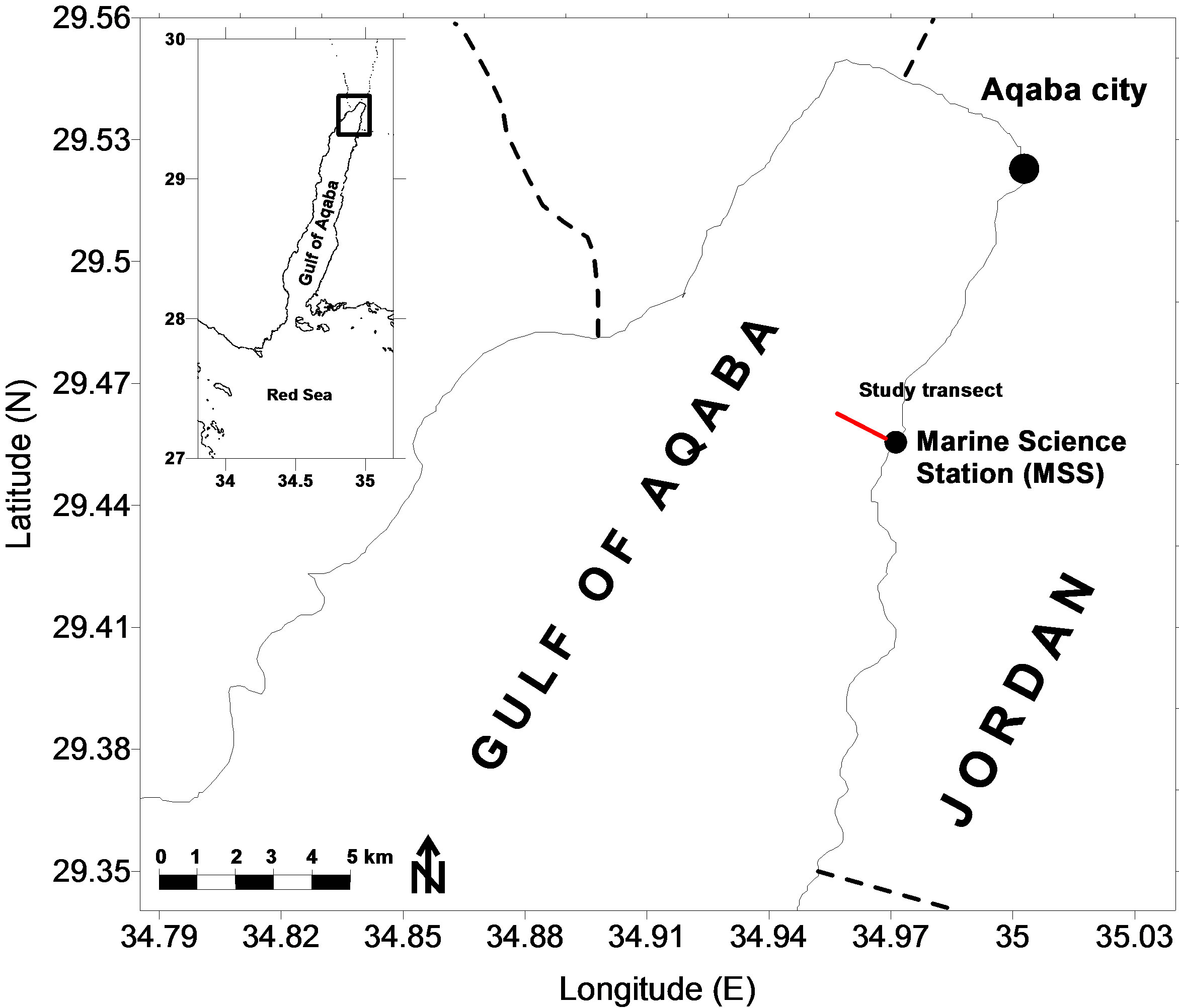
Figure 1. Map of the northern Gulf of Aqaba showing the location of the study area and the depth transect where coral samples were collected.
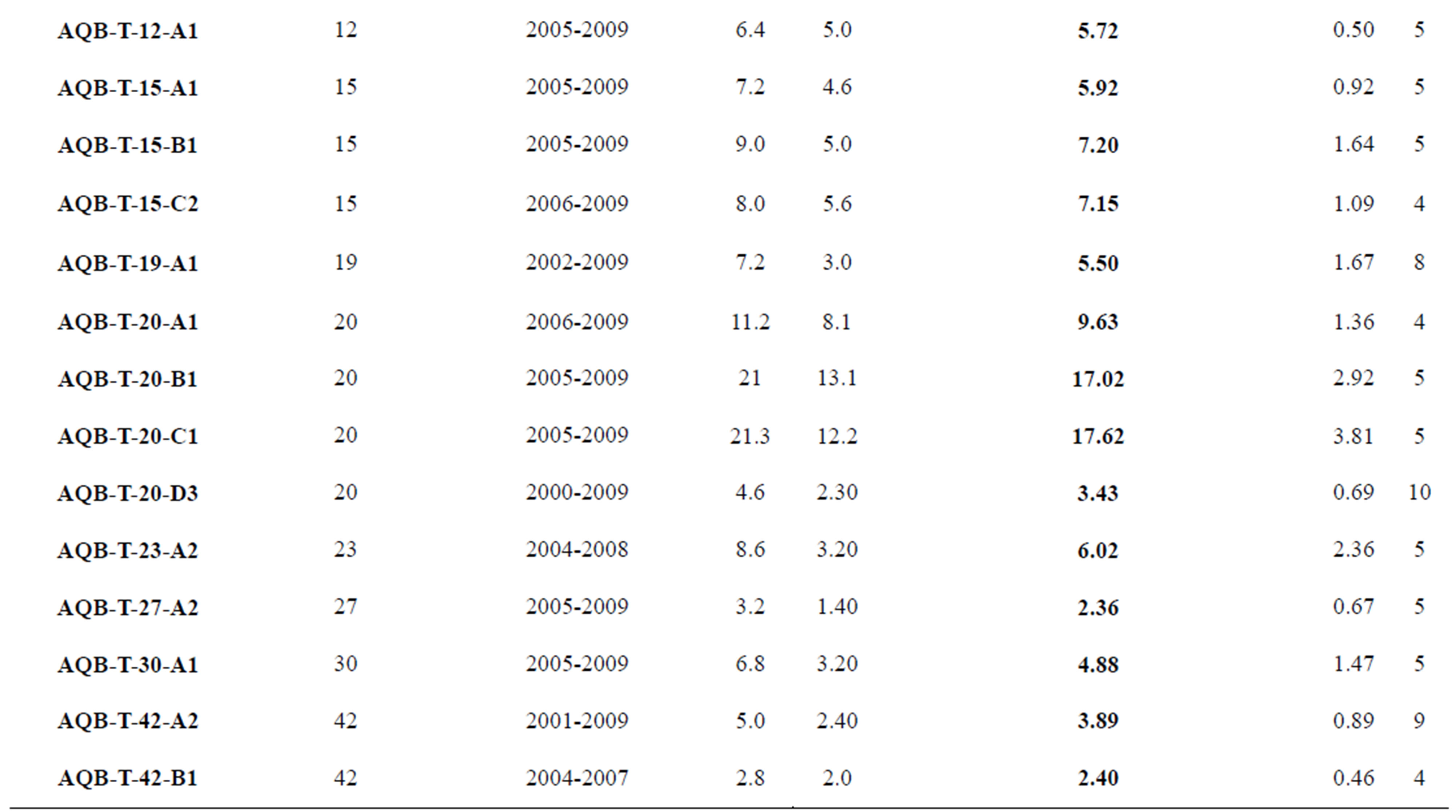

Table 1. Summary statistics for annual mean extension rates (mm/year), standard deviation (SD), minimum value (Min), maximum value (Max), and number of growth bands measured (N) for Porites spp. coral collected from Aqaba reef.
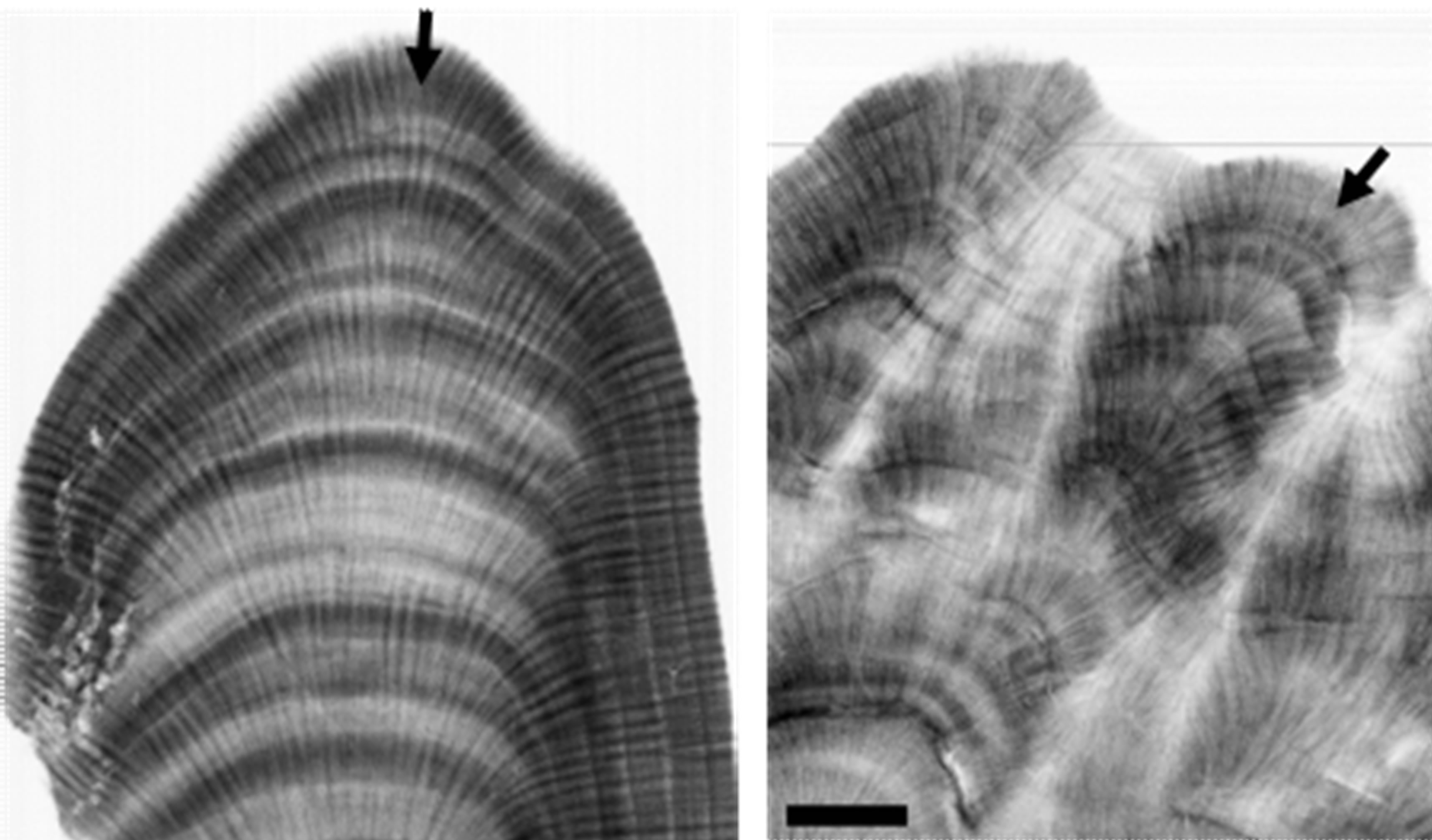
Figure 2. X-radiographs positive prints of shallow (AQB-T-10-D2, left) and deep (AQB-T-30-A1, right) Porites spp. corals from Aqaba reef. Skeletal density banding pattern of alternating bands of high and low density can be clearly seen. The major growth axes are indicated by black arrows. The black bar is 1 cm.
lowing year [31-33]. The mean extension rate of a given colony was calculated as the average of its annual values.
3. RESULTS AND DISCUSSION
3.1. Annual Density Growth Bands
Corals investigated in the present study reveal clear and regular skeletal density banding patterns of alternating bands of high and low-density (Figure 2). All coral samples were retrieved between April and June and they showed their high-density annual growth bands in the outermost parts suggesting that they were formed during winter season.
The timing of annual density bands formation from this study is similar to that obtained by Rosenfeld et al. [34] whom found that low-density skeleton is produced during summer and high-density skeleton is produced during winter in the same Porites corals grown in both shallow and deep growth phases (corals transplanted from 6 to 40 m). However, Klein et al. [30] documented the deposition of high-density bands during winter only in shallow water (3 m) Porites colony and opposite pattern in deep water (51 m) coral.
It was suggested that seawater temperature and light intensity are the major factors influencing the density patterns in Porites corals from the Red Sea [35]. The seawater temperature record in the study area shows strong seasonal pattern with highest temperature occurring in August (26.32˚C) and lowest in March (21.18˚C), [31].
3.2. Variation of Coral Extension Rates with Depth
The annual extension rates measured using the x-radiography and the δ18O (or Sr/Ca) curves reveal identical results. Mean annual extension rates of the 32 Porites spp. corals as obtained from 181 annual growth bands reveal a large inter-colony variability with values ranged between 2.36 to 20.0 mm/year (average 9.48 ± 4.98 mm/year). The maximum extension rate value of 20.0 mm/year was recorded in corals growing at a depth of 19 m and the minimum of 2.36, 2.40 mm/year in corals from 27 and 42 m depth, respectively (Table 1, Figure 3). These values fall within the global average of 12 mm/year in massive corals under normal conditions [14,15] and within the range of values presented by other authors from the Gulf of Aqaba [30,33,36-40]. For example, Heiss et al. [36] found that the growth rates of Porites corals in shallow water from Aqaba (1 - 3 m) vary between 8.64 - 12.26 mm/year, which is high for reefs at this latitude.
Most interesting, is the highest extension rates (15.2 - 20 mm/year) measured in corals growing at depths around 20 m (Aq19-A, Aq19a, Aq19b, AQB-T20-B1 and AQB-T-20-C1). This could be attributed to the position of these colonies, since they stand in the upper fore reef and the top of these colonies is about 1.5 m above the sea bottom. This will elevate the colony to a position of either more light, less abrasion by moving sand or both [23]. The same reason could be also responsible for the

Figure 3. Scatter plot and regression of the relationship between the mean annual extension rates of Porites spp. corals (mm/year) and depth (m) from Aqaba reef (Mean ± Standard Deviation). High values obtained from corals growing at 20 m depth were excluded.
high extension rate value (16.3 mm/year) obtained from coral AQB-Big which is a huge colony and lies only 1 m below seawater surface. Similarly, Heiss [41] found that growth rates higher than 8 mm/year could occur in colonies derived from a free-standing Porites pinnacle.
In Caribbean and Indo-Pacific regions, Porites corals exhibit annual growth increments of about 10 mm/year for shallow water region and a decrease to 4 mm/year in more deeper waters. However, it is noticed that even in the deeper water environment (e.g., 10 - 25 m), the annual growth rates is still high with up to 7 mm/year [10]. The decline in coral extension rates in deeper waters could reflect the position of a light threshold below which calcification is reduced as suggested earlier by Chalker et al. [42]. For Caribbean reefs, Adey [43] cited 15 m as a controlling depth for reef initiation after Holocene sea level rise slowed; it is also the cut off between shallow and deep reef species on many Caribbean reefs.
In general, coral extension rates from this study decreased with increasing water depth (excluding the high values from 19 - 20 m depth). The inverse relationship between mean annual extension rates and depth for Porites spp. corals is non-linear and can be explained by a simple exponential model as follows (Figure 3):
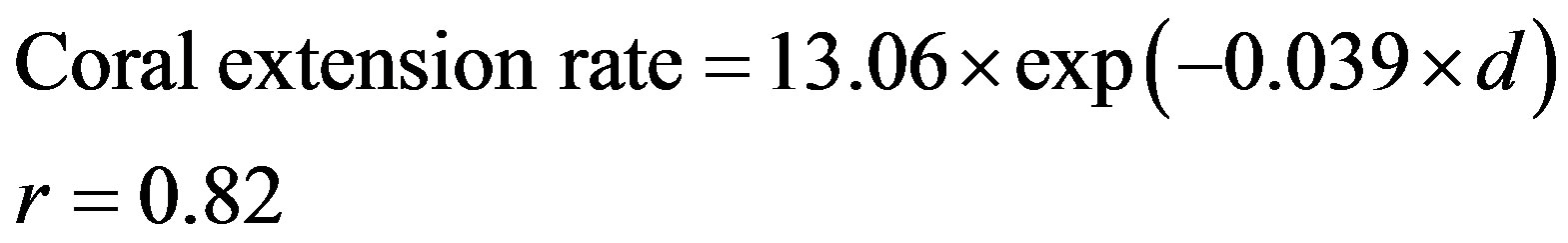
where coral extension rate in mm/year and d is the water depth in meter.
The values tended to cluster around two groups, one at 10.86 mm/year in water depths less than 10 m (range between 7.8 - 16.3 mm/year), and the other around 5.23 mm/year below 12 m depth (range between 2.36 - 9.63 mm/year). The analysis of variance (one way ANOVA) showed that there is statistically significant difference in the mean extension rate of corals between the two groups (p < 0.0001).
The environmental factors that may control extension rates in massive coral include water depth, sedimentation and turbidity, nutrient concentration, seawater temperature and salinity [17,44,45]. In the Gulf of Aqaba, environmental factors such as precipitation, cloud cover and freshwater discharge (terrigenous sediment input) and its effect on salinity, sedimentation and turbidity can be eliminated since the area is a desert-enclosed sea, and annual precipitation is less than 2.2 cm/year [27]. Therefore, the Gulf of Aqaba is characterized by high water clarity and deep light penetration [29].
Depth is considered as a mixed environmental variable including the effect of light, water movement and resuspension of sediment and organic matter, and in some places temperature and/or salinity [10,14]. Seawater temperature effects are most probably not the reason for the decrease in coral extension rate with depth in this study since the Gulf of Aqaba is isothermal over the studied depth range. In the study area and from the same reef transect, the annual water temperature decreases with depth between the depths 7 and 42 m by less than 0.25˚C, and the salinity variations was less than 0.3‰ [31]. Light intensity (mE·m−2·s−1) was measured at monthly basis at a steep reef slope (3 - 40 m depth) from the study area by Kampmann [46]. The record shows that light levels decreases exponentially with depth and coral extension rates from this study follow the same trend (Figure 4).

Figure 4. Scatter plot for the average annual underwater light profile intensities (mE·m−2·s−1) at a steep reef slope between 3 - 40 m depth off the Marine Science Station in the Gulf of Aqaba (after [46]), and the mean annual extension rates of all Porites spp. corals (mm/year) investigated in this study.
At 40 m depths, light intensity drops to about 50 mE·m−2·s−1 in which extension rate of corals is minimum (2.40 mm/year). Therefore, light seems to be the primary environmental factor responsible for decreasing coral extension rate with depth and other parameters are fairly constant throughout the water column from the Gulf of Aqaba. Light enhanced calcification is responsible for most of the skeletal growth and carbonate production of reef building corals [47,48].
3.3. Temporal Variability in Coral Extension Rates
In inter-annual time scale, coral extension rate varies within the same colony over the studied time span. The time series record showed a fluctuated pattern with slightly increasing trend of the extension rate in both shallow and deep corals for the period of time represented despite the significant difference in extension rate between them (Figure 5).
Local sea surface temperature record (SST) from the study site shows similar increasing trend between 1988- 2009 (Figure 5). However, the correlation between the SST record and extension rates from both corals is not strong (r = 0.38, 0.24 for AQB-Big and AQB-T-42-A2, respectively). Several studies have documented the sensitivity of extension in Porites corals to temperature [16,17] over a wide SST ranges (22˚C - 29˚C) and they reported a significant positive correlation between both variables [17]. In our study, the range of inter-annual SST variability during the overall period (1988-2009) is relatively too small (fluctuated by less than 1.2˚C) to be a factor of primary importance affecting coral annual extension rates. Therefore, longer time series records of both SST and annual coral extension rate are needed to validate this relation from the northern Gulf of Aqaba.
4. CONCLUSION
The reef building Porites spp. corals from Aqaba show clear and regular skeletal annual density banding patterns, the high-density growth bands seems to be laid down during winter and the low-density bands during summer. The average extension rates for Porites corals are around 10.86 ± 2.54 mm/year in water depths less than 10 m, and decreases to 5.23 ± 1.99 mm/year between 12 - 42 m. The relationship between mean coral extension rate and depth is best explained by exponential model as a response to decreasing light availability and photosynthesis with depth. However, the effect of other environmental parameters such as sedimentation and turbidity could be neglected throughout the water column and over the studied depths. On inter-annual time scale, seawater temperature variability seems to have an effect on the mean annual coral extension rates suggesting the sensitivity of Porites spp. corals from the Gulf of Aqaba to global warming effects.
5. ACKNOWLEDGEMENTS
I wish to thank Abdullah Al-Momany and the Marine Science Station diver staff in Aqaba for their assistance in coral samples collection. Dr. Riyad Manasrah kindly provided seawater temperature data from Aqaba. Special thanks are also due to Dr. Thomas Felis for facilitating
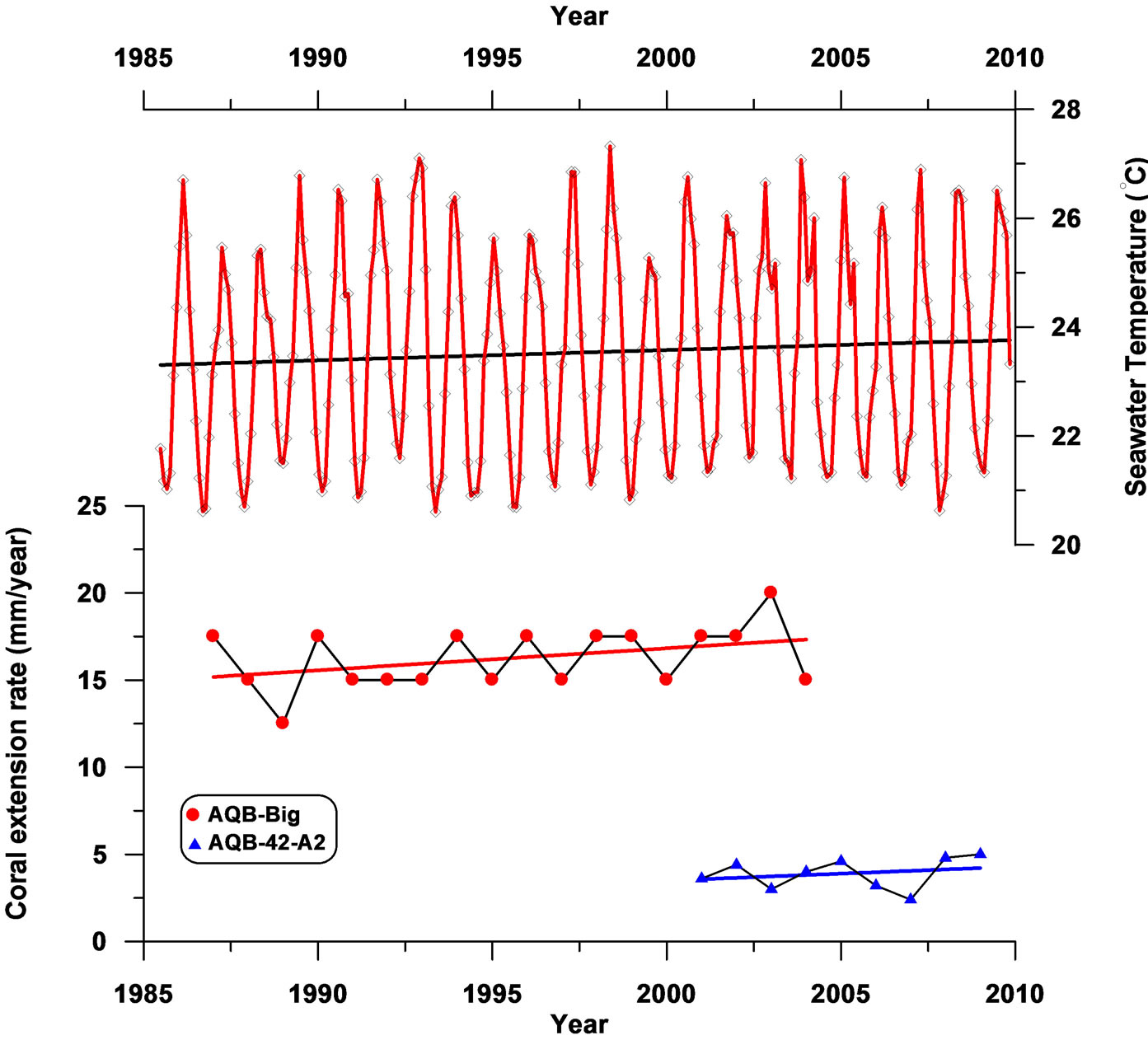
Figure 5. Time series records of the annual extension rates (mm/year) for shallow (AQB-Big) and deep (AQB-T-42-A2) Porites spp. corals and the monthly recorded sea surface temperatures (˚C) in Aqaba between 1988 and 2009 ([28], Manasrah personal communication 2012).
coral slicing and X-radiography work at the University of Bremen, Germany.
REFERENCES
- Pätzold, J. (1984) Growth rhythms recorded in stable isotope and density bands in reef coral Porites lobata (Cebu, Phillippines). Coral Reefs, 3, 87-90. doi:10.1007/BF00263758
- Linsley, B.K., Dunbar, R.B., Wellington, G.M. and Mucciarone, D.A. (1994) A coral based reconstruction of intertropical convergence zone variability over central America since 1707. Journal of Geophysical Research, 99, 9977-9994. doi:10.1029/94JC00360
- Felis, T. and Pätzold, J. (2003) Climate records from corals. In: Wefer, G., Lamy, F. and Mantoura, F., Eds., Marine Science Frontiers for Europe, Springer, Berlin, 11-27. doi:10.1007/978-3-642-55862-7_2
- Corrège, T. (2006) Sea surface temperature and salinity reconstruction from coral geochemical tracers. Palaeogeography Palaeoclimatology Palaeoecology, 232, 408- 428. doi:10.1016/j.palaeo.2005.10.014
- Eakin, M. and Grottoli, A.G. (2006) Paleo-climate changes and corals. In: Phinney, J., Skirving, W., Kleypas, J., Hoegh-Guldberg, O. and Strong, A.E., Eds., Coral Reefs and Climate Change: Science and Management. Coastal and Estuarine Studies, 61, 33-54.
- Gischler, E., Hudson, J. H. and Storz, D. (2009) Growth of pleistocene massive corals in south Florida: low skeletal extension-rates and possible ENSO, decadal, and multi-decadal cyclicities. Coral Reefs, 28, 823-830. doi:10.1007/s00338-009-0537-1
- Chalker, B.E. and Barnes, D.J. (1990) Gamma densitometry for the measurement of coral skeletal density. Coral Reefs, 9, 11-23. doi:10.1007/BF00686717
- Logan, A. and Tomascik, T. (1991) Extension growth rates in two coral species from high-latitude reefs of Bermuda. Coral Reefs, 10, 155-160. doi:10.1007/BF00572174
- Guzman, H.M. and Cortes, J. (1989) Growth rates of eight species of scleractinian corals in the eastern Pacific (Costa Rica). Bulletin Marine Science, 44, 1186-1194.
- Dullo, W.-Chr. (2005) Coral growth and reef growth: a brief review. Facies, 51, 33-48. doi:10.1007/s10347-005-0060-y
- Guzman, H.M., Cipriani, R. and Jackson J.B.C. (2008) Historical decline in coral reef growth after the Panama Canal. Ambio, 37, 342-346. doi:10.1579/07-A-372.1
- Knutson, D.W., Buddemeier, R.W. and Smith, S.V. (1972) Coral chronometers: Seasonal growth bands in reef corals. Science, 177, 270-272. doi:10.1126/science.177.4045.270
- Scoffin, T.P., Tudhope, A.W., Brown, B.E., Chansang, H. and Cheeney, R.F. (1992) Patterns and possible environmental controls of skeletogenesis of Porites lutea, South Thailand. Coral Reefs, 11, 1-11. doi:10.1007/BF00291929
- Buddemeier, R.W. and Kinzie, R.A. (1976) Coral growth. Oceanography and Marine Biology: Annual Review, 14, 183-225.
- Lough, J.M. and Barnes, D.J. (1997) Coral records of past climates and environments. Bulletin of Australian Meteorological and Oceanographic Society, 10, 84-90.
- Lough, J.M. and Barnes, D.J. (2000) Environmental controls on growth of the massive coral Porites. Journal of Experimental Marine Biology and Ecology, 245, 225-243. doi:10.1016/S0022-0981(99)00168-9
- Lough, J.M. and Cooper, T.F. (2011) New insights from coral growth band studies in an era of rapid environmental change. Earth Science Reviews, 108, 170-184. doi:10.1016/j.earscirev.2011.07.001
- Pätzold, J. and Wefer, G. (1992) Bermuda coral reef record of the last 1000 years. 4th International Conference on Paleoceanography. Geol.-Paläont. Inst Univ Kiel, Kiel, 224-225.
- Pätzold, J., Bickert, T., Flemming, B., Grobe, H. and Wefer, G. (1999) Holozänes Klima des Nordatlantiks rekonstruiert aus massiven Korallen von Bermuda. Natur und Museum, 129, 165-177.
- Crossland, C.J. (1988) Latitudinal comparisons of coral reef structure and function. Proceeding of the 6th International Coral Reef Symposium, 1, 221-226.
- Isdale, P.J. (1983) Geographical patterns in coral growth rates on the Great Barrier Reef. In: Baker, J.T., Carter, R.M., Sammarco, P.W. and Stark, K.P., Eds., Inaugural Great Barrier Reef Conference, James Cook University Press, Townsville, 28 August-2 September 1983, 327- 330.
- Risk, M.J. and Sammarco, P.W. (1991). Cross-shelf trends in skeletal density of the massive coral Porites lobata from the Great Barrier Reef. Marine Ecology Progress Series, 69, 195-200. doi:10.3354/meps069195
- Hubbard, D.K. and Scaturo, D. (1985) Growth rates for seven scleractinean corals from Cane Bay and Salt River, St. Croix, USVI. Bulletin of Marine Sciences, 36, 325- 338.
- Schuhmacher, H., Kienne, W. and Dullo, W-Chr. (1995) Factors controlling Holocene Reef growth: An interdisciplinary approach. Facies, 32, 145-188. doi:10.1007/BF02536867
- Felis, T., and Rimbu, N. (2010) Mediterranean climate variability documented in oxygen isotope records from northern Red Sea corals—A review, Global and Planetary Change, 71, 232-241. doi:10.1016/j.gloplacha.2009.10.006
- Gvirtzman, G., and Buchbinder, B. (1978) The Late Tertiary of the Coastal Plain and Continental Shelf of Israel and its bearing on the history of the Eastern Mediterranean. Deep Sea Drilling Project, 42, 1195-1222.
- Reiss, Z. and Hottinger, L. (1984) The gulf of Aqaba: Ecological micropaleontology. Springer-Verlag, Berlin, 345. doi:10.1007/978-3-642-69787-6
- Manasrah, R. (2002) The general circulation and water masses characteristics in the gulf of Aqaba and northern Red Sea. Ph.D. Thesis, Universität Rostock, Rostock.
- Levanon-Spanier, I., Padan, E. and Reiss, Z. (1979) Primary production in a desert enclosed sea-the gulf of Elat (Aqaba), Red Sea. Deep Sea Research, 26, 673-685. doi:10.1016/0198-0149(79)90040-2
- Klein, R., Pätzold, J., Wefer, G. and Loya, Y. (1993) Depth-related timing of density band formation in Porites spp. corals from the Red Sea inferred from X-ray chronology and stable isotope composition. Marine Ecology Progress Series, 97, 99-104. doi:10.3354/meps097099
- Al-Rousan, S., Al-Moghrabi, S., Pätzold, J. and Wefer, G. (2003) Stable oxygen isotopes in Porites corals monitor weekly temperature variations in the northern gulf of Aqaba, Red Sea. Coral Reefs, 22, 346-356. doi:10.1007/s00338-003-0321-6
- Al-Rousan, S., Al-Moghrabi, S., Pätzold, J. and Wefer, G. (2002) Environmental and biological effects on the stable oxygen isotope records of corals in the northern gulf of Aqaba, Red Sea. Marine Ecology Progress Series, 239, 301-310. doi:10.3354/meps239301
- Felis, T., Pätzold, J. and Loya, Y. (2003) Mean oxygenisotope signatures in Porites spp. corals: Inter-colony variability and correction for extension-rate effects. Coral Reefs, 22, 328-336. doi:10.1007/s00338-003-0324-3
- Rosenfeld, M., Yam, R., Shemesh, A. and Loya, Y. (2003) Implication of water depth on stable isotope composition and skeletal density banding patterns in a Porites lutea colony: results from a long-term translocation experiment. Coral Reefs, 22, 337-345. doi:10.1007/s00338-003-0333-2
- Klein, R. and Loya, Y. (1991) Skeletal growth and density patterns of two Porites corals from the gulf of Eilat, Red Sea. Marine Ecology Progress Series, 77, 253-259. doi:10.3354/meps077253
- Heiss, G.A., Dullo, W.-Chr. and Reijmer, J.J.G. (1993) Shortand long-term growth history of massive Porites sp. from Aqaba (Red Sea). Senckenbergiana Maritima, 23, 135-141.
- Heiss, G.A. (1996) Variation in annual band width of Porites sp. from Aqaba, Gulf of Aqaba, Red Sea. Bulletin of Marine Science, 59, 393-403.
- Heiss, G. A., Dullo, W.-Chr., Joachimski, M.M, Reijmer, J.J.G. and Schuhmacher, H. (1999) Increased seasonality in the gulf of Aqaba, Red Sea, recorded in the oxygen isotope record of a Porites lutea coral. Senckenbergiana Maritima, 30, 17-26. doi:10.1007/BF03042826
- Felis, T., Pätzold, J., Loya, Y., and Wefer, G. (1998) Vertical water mass mixing and plankton blooms recorded in skeletal stable carbon isotopes of a Red Sea coral. Journal of Geophysical Research, 103, 30731-30739. doi:10.1029/98JC02711
- Felis, T., Lohmann, G., Kuhnert, H., Lorenz, S.J., Scholz, D., Pätzold, J., Al-Rousan, S.A. and Al-Moghrabi, S.M. (2004) Increased seasonality in Middle East temperatures during the last interglacial period. Nature, 429, 164-168. doi:10.1038/nature02546
- Heiss, G.A. (1994) Coral reefs in the Red Sea: Growth, production and stable isotopes. GEOMAR Report 34, 1-143.
- Chalker, B.E. and Taylor, D.L. (1975) Light-enhanced calcification, and the role of oxidative phosphorylation in calcification of the coral Acropora cervicornis. Proceeding Royal Society London Bulletin, 190, 323-331. doi:10.1098/rspb.1975.0096
- Adey, W. (1978) Coral reef morphogenesis: a multi-dimensional model. Science, 202, 831-837. doi:10.1126/science.202.4370.831
- Baker, P.A. and Weber J.N. (1975) Coral growth rate: Variation with depth. Earth Planetary Science Letters, 27, 57-61. doi:10.1016/0012-821X(75)90160-0
- Hudson, H.J. (1981) Growth rates in Montastrea annularis: a record of environmental change in the Florida Keys. Proceeding of the 4th International Coral Reef Symposium, 233-240.
- Kampmann, H. (2001) Photobiologische, energtische und genetische aspekt des mutualistischen zusammen lebenes von Zooxanthellae (Symbiodininm sp.) und Steinkorrallen im golf von Aqaba. Jordanien. Dissertation, Mathematisch-Naturwissenschaftliche Facultaet, Universitaet zu Koeln.
- Bosscher, H. (1992) Growth potential of coral reefs and carbonate platforms. Ph.D. thesis, Vrije Universiteit Amsterdam, Amsterdam.
- Mutti, M. and Hallock, P. (2003) Carbonate systems along nutrient and temperature gradients: some sedimentological and geochemical constraints. International Journal of Earth Sciences, 92, 465-474. doi:10.1007/s00531-003-0350-y