American Journal of Plant Sciences
Vol.5 No.16(2014), Article
ID:48406,13
pages
DOI:10.4236/ajps.2014.516268
Iron Deficiency Tolerance at Leaf Level in Medicago ciliaris Plants
Wissal M’sehli1*#, Hayet Houmani2#, Silvia Donnini3, Graziano Zocchi3, Chedly Abdelly2, Mohammed Gharsalli2
1Laboratory of Legumes (LL), Biotechnology Centre of Borj Cedria (CBBC), Hammam-Lif, Tunisia
2Laboratory of Extremophile Plants (LPE), Biotechnology Centre of Borj Cedria (CBBC), Hammam-Lif, Tunisia
3Dipartimento di Produzione Vegetale, University of Milan, Milan, Italy
Email: *wissal_msehli@yahoo.fr
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 24 May 2014; revised 28 June 2014; accepted 21 July 2014
ABSTRACT
Iron deficiency is an important environmental factor restricting plant productivity. Selecting tolerant genotypes is one of the possible ways to solve this problem. Many studies reported the effects of Fe deficiency on photosynthesis and anti-oxidative defense system. Yet, there is little information available on the use of these attributes as selective criteria. In the present study, we aim to determine some physiological and biochemical traits conferring Fe deficiency tolerance at leaf level in two lines of Medicago ciliaris. Our results showed that Fe deprivation had a lowering effect on photosynthesis (chlorophyll, photosynthetic electron transport activity and chlorophyll fluorescence) in both lines studied. However, the sensitive line TN8.7 was more affected. Hydrogen peroxide concentration was negatively correlated with the activities of antioxidant enzymes and with the concentration of some non-enzymatic antioxidant. The tolerant line TN11.11 was characterized by a more efficient antioxidant defense system in comparison with the sensitive line TN8.7. The main conclusion of this study is that photosynthesis and antioxidant defense system could be used as physiological and biochemical indicators of Fe deficiency tolerance in Medicago ciliaris plants.
Keywords:Iron Deficiency, Medicago ciliaris, Photosynthesis, Oxidative Stress, Tolerance Indicators

1. Introduction
Under natural conditions, plants are exposed to many constraints that significantly reduce their growth and productivity. Iron (Fe) deficiency is considered as a major factor threatening plant development in arid and semiarid regions, especially in calcareous soils leading to the appearance of Fe chlorosis symptoms in plants [1] .
Iron deficiency affects plant morphological and physiological responses because of the fundamental role of Fe in several biological processes. In fact, Fe is a constituent of electron transport chains of both mitochondria and chloroplasts. As a result of its absence, a decrease in the activity of electron transport [2] and a diminution of photosynthetic pigments were noted, resulting in a reduction of net photosynthesis per unit area [3] . Moreover, Fe is actively involved in cellular reactions of detoxification because it is present, as an heme moiety, in several anti-oxidant enzymes such as catalases (CAT) and peroxidases (POD), which both ensure the reduction of H2O2, and as metal co-factor in Fe-superoxide dismutase (Fe-SOD) which, together with the other metal isoforms, converts the superoxide anion to H2O2 [4] . Ascorbate peroxidase (APx) and guaiacol peroxidase are distributed throughout the cell and catalyze the reduction of H2O2 to H2O [5] . Ascorbate peroxidase, which contains a nonheme Fe [6] , uses ascorbate as electron donor in the first step of the ascorbate-glutathione cycle and is considered as the most important plant peroxidase in H2O2 detoxification [7] .
In previous works [8] -[10] , we found that Fe deficiency tolerance in Medicago ciliaris line TN 11.11 is mainly associated with the activation of root plasma membrane H+-ATPase and Fe (III) reductase. In the present study, we investigate some possible mechanisms responsible for Fe deficiency tolerance at leaf level in two Medicago ciliaris lines in order to identify leaf parameters useful for rapid screening of genotypes. For that purpose, we firstly, studied the effect of Fe deficiency on the whole photosynthetic electron transport chain and on the chlorophyll a fluorescence parameters. Then, we evaluated the influence of Fe deficiency on lipid peroxidation and the role of several enzymes of the antioxidant machinery in the production and/or degradation of reactive oxygen species (ROS). The content of some anti-oxidant molecules, such as glutathione and ascorbic acid, has been also determined.
2. Materials and Methods
2.1. Plant Material
Two lines of Medicago ciliaris L. created by three generations of spontaneous selfing in the greenhouse were used: TN11.11 and TN8.7. Seeds were obtained from the Laboratory of Legumes, Biotechnology Center of Borj-Cedria (CBBC), Tunisia.
Seeds were germinated and grown for 3 days in Agriperlite moistened with 0.1 mM CaSO4. Three-day-old seedlings were transferred to a half strength aerated hydroponic nutrient solution for 7 days and then similar sized seedlings were selected and cultured as groups of 10 plants in 10 L of full strength aerated hydroponic nutrient solution. The composition of the nutrient solution was: 1.25 mM Ca (NO3)2, 1.25 mM KNO3, 0.5 mM MgSO4, 0.25 mM KH2PO4 and 10 µM H3BO3, 1 µM MnSO4, 0.5 µM ZnSO4, 0.05 µM (NH4)6Mo7O24 and 0.4 µM CuSO4.
Two treatments were applied: −Fe (0 mM) and +Fe (0.03 mM). Both media have a pH around 6.2. Fe was supplied as Fe (III)-EDTA. The solution was renewed every 5 days. Plants were maintained in a growth chamber with a 16/8 h day/night, 24˚C/18˚C regime and a relative humidity of 70%.
2.2. Bivalent Iron Content (Fe2+)
Bivalent Fe content was determined according to the method of [11] modified by [12] . After 10 days of treatment, leaves were weighted and dried at 60˚C for 72 h. Then, samples were finely crushed in a standard agate crusher in order to avoid powder contamination by Fe traces. Then, 2 g of dry samples were digested with 15 ml HCl(1N) and vigorously agitated. Digests were thereafter kept for 4 h in the vessels and then filtered before bringing them to a volume of 25 mL with distilled water. The absorbance of the solution was determined by an atomic absorption spectrophotometer (VARIAN 220FS).
2.3. Chlorophyll Content
Eight plants per treatment were used to determine total chlorophyll content of young leaves according to the method of [13] . One hundred mg of small discs from young leaves were incubated in 5 mL 80% acetone in darkness at 4˚C for three days (until complete chlorophyll extraction). Total chlorophyll content was determined by reading the absorbance at 649 and 665 nm.
2.4. Photosynthetic Electron Transport Activity
Thylakoid membranes were isolated from leaves as described by [14] . Photosynthetic electron transport rates through the two photosystems (PSI and PSII) were measured on isolated thylakoid membranes by means of Clark-type oxygen electrode (Hansatech, UK) fitted with a circulating water jacket at 20˚C, using a light intensity of 1100 µ E m−2∙s−1 at the surface of the water bath cell. The sample volume was 1 mL and the chlorophyll content was 10, 20 and 40 µg. The uncoupled rate of the whole chain electron transport was determined by measuring the rate of reoxidation of photoreduced methyl viologen (MV). The reaction buffer contained 100 mM sorbitol, 5 mM MgCl2, 0.5 mM NaN3, 3 mM NH4Cl, 0.1 mM MV, and 50 mM HEPES-NaOH pH 8.0 [3] .
2.5. Measurement of Leaf Chlorophyll Fluorescence
Iron-sufficient and Fe-deficient M. ciliaris leaves were adapted to dark conditions for 1h at room temperature before experiments. Chla fluorescence was measured in leaf discs using a PAM 2000 fluoremeter (Heinz Walz, Germany). The measurements were performed at room temperature (25˚C). F0 was measured by switching on the modulated light at 0.6 kHz; PPFD was less than 0.1 µ E m−2∙s−1 at the leaf surface. Fm was measured at 20 kHz with a 1s pulse of 6000 µ E m−2∙s−1 of white light. The initial fluorescence (F0) and the maximal fluorescence were determined after dark acclimation. The variable fluorescence (Fv) was taken from the formula: Fv = Fm − F0 and the parameter used were Fv/Fm and 1/F0 − 1/Fm [15] .
2.6. Hydrogen Peroxide Determination
The H2O2 level was calorimetrically measured as described by [16] . H2O2 was extracted by homogenizing 50 mg leaves with 3 mL of phosphate buffer (50 mM, pH 6.8). The homogenate extract was centrifuged at 6,000g for 25 min. To determine H2O2 levels, 3 mL of extracted solution were mixed with 1 mL of 0.1% titanium chloride in 20% (v/v) H2SO4 and the mixture was then centrifuged at 6000g for 15 min. The intensity of the yellow colour of the supernatant was measured at 410 nm. Hydrogen peroxide level was calculated using the extinction coefficient 0.28 µM−1∙cm−1.
2.7. Production of Volatile Aldehydes
Volatile aldehydes were extracted in 5 ml of 2% (v/v) ethanol at 25˚C for 2 hours, under constant stirring using 0.3 g leaf fresh tissues. A 1 mL aliquot was transferred to a test tube containing 1 mL of 0.1% (w/v) 3-methyl-2- benzothiazolinone hydrazone (MBTH) before the addition of 2.5 mL of FeCl36H2O solution (0.23% (w/v)). After 5 min of incubation, 6 mL of acetone were added and the absorbance was read at 635 nm [17] . The amount of volatile aldehydes produced was estimated using a formaldehyde calibration curve.
2.8. Lipid Peroxidation
Lipid peroxidation was determined as described by [18] : 0.2 g of fresh leaves were homogenized in 4 mL of 1% (w/v) trichloroacetic acid (TCA). The homogenate was filtered through four layers of gauze and then centrifuged at 12,000 g for 15 min. One mL aliquots of the supernatants were added to 3 ml of 0.5% (w/v) thiobarbituric acid (TBA) in 20 % (w/v) (TCA) and the tubes were incubated in a shaking water bath at 95˚C for 2 h. The reaction was stopped by placing the tubes in ice. Tubes were subsequently centrifuged at 9000 g for 10 min and the absorbance of the supernatant was read at 532 and 600 nm. The concentration of the malondialdehyde (MDA)-TBA complex produced was calculated using the molar extinction coefficient of 155 mM−1∙cm−1.
2.9. Antioxidant Enzyme Assays
All operations were carried out at 4˚C. Leaves were homogenised with quartz sand, liquid nitrogen and 10% (w/w) polyvinylpyrolidone (PVP) in a grinding medium (1:2.5, w:v) prepared as described below, and centrifuged at 12000 g for 30 min at 4˚C.
The extraction buffer for superoxide dismutase (SOD) and guaiacol-POD (POD) contained 220 mM Tris-HCl (pH 7.4), 250 mM sucrose, 50 mM KCl, 1 mM MgCl2, 1% β mercaptoethanol and 0.01% (w/v) phenylmethylsulfonylfluoride (PMSF) [19] . For ascorbate peroxidase (APx) extraction, 50 mM Na-ascorbate was added to the buffer to prevent enzyme inactivation [19] . After centrifugation, the supernatant was dialyzed for 2 h against 50 mM reduced ascorbate solution (ASA) and tested for enzyme activity.
Superoxide dismutase activity was measured according to the method of [20] based on the enzyme ability to inhibit the reduction of nitrobluetetrazolium (NBT). One enzymatic unit is defined as the amount of enzyme inhibiting 50% of NBT photoreduction.
Peroxidase activity was tested using guaiacol as reducing phenolic substrate. The reaction medium contained 20 mM Na-acetate (pH 5.0), 30 mM H2O2, 2 mM guaiacol and an appropriate amount of enzyme extract. The rate of guaiacol oxidation was recorded at 470 nm [19] and the activity was calculated using the extinction coefficient of 26.6 mM−1∙cm−1.
Ascorbate peroxidase activity was determined following the decrease in absorbance at 290 nm due to the oxidation of ascorbic acid in the first 30 s from the beginning of the reaction, using the extinction coefficient of 2.8 mM−1∙cm−1 for ascorbate. The reaction medium contained 0.5 mM reduced ascorbate (ASA), 0.1 mM H2O2, 1 mM EDTA and 0.1 M HEPES-KOH buffer (pH 7.8) [21] . One enzymatic unity is equivalent to 1 µmol min−1∙cm−1 of ascorbic acid oxidized. To discriminate between APx and POD activity, 50 mM p-chloromercuribenzoate (pCMB), responsible for the inactivation of APx, was added to the enzymatic reaction mixture [22] .
Non-enzymatic antioxidant activity, ascorbate and glutathione content.
Total non-enzymatic antioxidant activity was measured using the Ferric Reducing Ability of Plasma (FRAP) assay. Leaf tissue (0.5 g) was homogenized in 5 ml methanol. The homogenate was first filtered and then centrifuged at 12,800 g for 2 min. The FRAP assay was performed with FRAP reagent, i.e. 1 mM 2,4,6-tripyridyl-2- triazine (TPTZ) and 20 mM ferric chloride in 0.25 M sodium acetate (pH 3.6). An aliquot of 100 ml of leaf extract (0.5 g/10 mL in methanol) was added to 2 mL of FRAP reagent and thoroughly mixed. Next, the mixture was left at 20˚C for 5 min and the absorbance was measured at 593 nm. Calibration was obtained by means of a standard curve (25 - 1600 mM ferrous iron) using freshly prepared ammonium ferrous sulfate [23] [24] .
Extraction of ascorbate and glutathione Leaf samples were ground in a cold mortar with liquid nitrogen and homogenized with 25 mM sulphuric acid (800 µl/400 mg fresh weight). The homogenate was centrifuged at 16,700 g for 30 min at 4˚C, and the supernatant was collected for analysis of ascorbate and glutathione.
Quantification of reduced (ASC) and oxidized (DHA) ascorbate The assay is based on the reduction of Fe3+ to Fe2+ by ascorbic acid [25] . Fe2+ forms a red complex with bipyridyl that absorbs at 550 nm. DHA was reduced to ASC by pre-incubating the sample with dithiothreitol (DTT). After DTT excess was removed with N-ethylmaleimide (NEM), total ascorbate (ASC+DHA) content was measured. DHA content was then estimated from the difference between total ascorbate and the ASC. A standard curve covering the range of 2.5 - 60 µmol ASC was used.
Quantification of reduced (GSH) and oxidized (GSSG) glutathione The contents of GSH and GSSG were determined as described by [21] based on the oxidation of GSH by DTNB (5,5'-dithio-bis-nitrobenzoic acid) to give GSSG and TNB (5-thio-2-nitrobenzene). GSSG was reduced to GSH by the action of glutathione reductase and NADPH. GSSG was assayed from the sample after the removal of GSH by 2-vinylpyridine and triethanolamine derivatizations. The rate of DTNB reduction was recorded at 412 nm for 2 min. The contents were calculated using a standard curve. The GSH content was calculated from the difference between the total glutathione and GSSG.
2.10. Statistical Analysis
A two-way analysis of variance (ANOVA), with lines and treatments as factors, was performed for the whole data using the STATI-CF statistical program. Means were compared using the Newman-Keuls test at the p<0.05 level when significant differences were found. Data shown are means of five replicates for each treatment.
3. Results
3.1. Chlorophyll and Bivalent Iron Concentrations
Medicago ciliaris plants grown under Fe deficiency conditions for 10 days developed symptoms of chlorosis. These visual manifestations were observed in both lines (TN8.7 and TN11.11) and were concomitant with a significant reduction in chlorophyll concentration (Table 1), especially for TN8.7 line (−51%). The same trend was observed for leaf Fe concentration; this decrease respect to controls was more pronounced in TN8.7 (−37%) than in TN11.11 plants (−23%) (Table 1).
3.2. Photosynthetic Electron Transport Activity and Chlorophyll Fluorescence
Iron deficiency affected significantly the whole chain photoelectron transport rate of both genotypes TN8.7 and TN11.11 (Figure 1). As for chlorophyll and Fe2+ concentrations, TN8.7 exhibited the strongest reduction (57%) as compared to control plants, while for TN11.11 this decrease was less than 25%.
The measurement of chlorophyll fluorescence as the ratio Fv/Fm revealed that at the beginning of the experiment, this parameter was similar in the control leaves of both genotypes (Table 2). Under Fe deficiency conditions, this ratio decreased significantly in chlorotic leaves of both lines. This reduction was about −23% and −33% in TN11.11 and TN8.7, respectively.
3.3. Parameters Related to Oxidative Stress (Lipid Peroxidation, Volatile Aldehydes and H2O2 Content)
For all samples, lipid peroxidation estimated by the MDA-TBA complex concentration was evaluated (Figure 2). Plants optimally supplied with Fe showed similar values. Under Fe deficiency conditions, an important increase in leaf MDA content was noted in both lines. Yet, this increase was more marked in TN8.7 (+140%) compared to TN11.11 (+95%) (Figure 2).

Table 1. Bivalent iron content (µmol∙g−1 DW) and chlorophyll concentration (mg∙g−1 FW) of two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
Table 2. Evolution of fluorescence parameters measured in leaves of two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
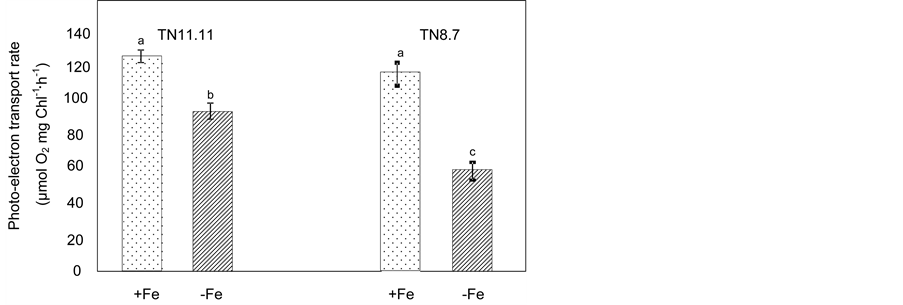
Figure 1. Photo-electron transport rate of two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
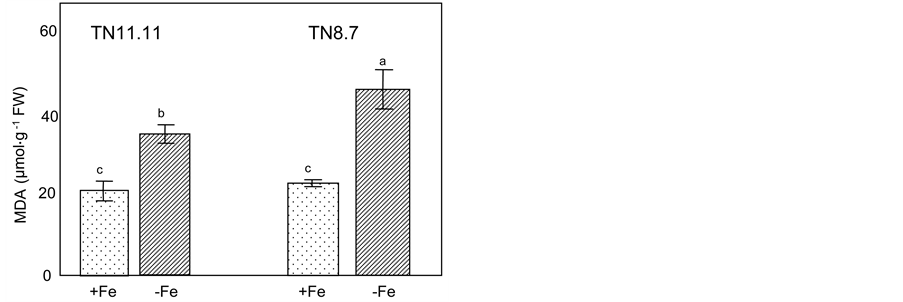
Figure 2. Leaf malondialdehyde content (MDA) of two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
The concentration of volatile aldehydes as an indicator of oxidative stress degree (mg∙g−1 FW) was determined (Figure 3). Our results showed that for TN11.11 no difference was found between plants supplied or not with Fe. Yet, a significant increase (+127%) was detected inTN8.7 Fe-deficient plants respect to the control.
The leaf tissue content of H2O2, another signal of oxidative stress, was measured. Hydrogen peroxide content significantly increased in both genotypes grown under Fe deprivation (Figure 4), suggesting the induction of oxidative stress in leaves of Fe starved plants. The highest value was detected in TN8.7 line (+68 % as compared to control plants), while for TN11.11, this increase was about 10%.
3.4. Antioxidant Enzymes
Total SOD activity was significantly stimulated under Fe deficiency in TN8.7 (+32% respect to the control), while in TN11.11 it was increased only by 13% (Table 3). To investigate whether, and to what extent, this increment was ascribable to different enzymatic isoforms, KCN and H2O2 were added to the enzymatic assay mixture, as different inhibitors of the three different SOD isoforms: CuZn-SOD, Mn-SOD and Fe-SOD. Results revealed that Fe deficiency induced an enhancement of CuZn-SOD (KCN-sensitive) and Mn-SOD (KCN and H2O2- insensitive) isoform activities, especially in TN8.7, in which the activity of both SOD isoforms was increased around two folds as compared to control plants. By contrast, Fe-SOD activity (H2O2-sensitive and

Figure 3. Leaf volatile aldehyde content of two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
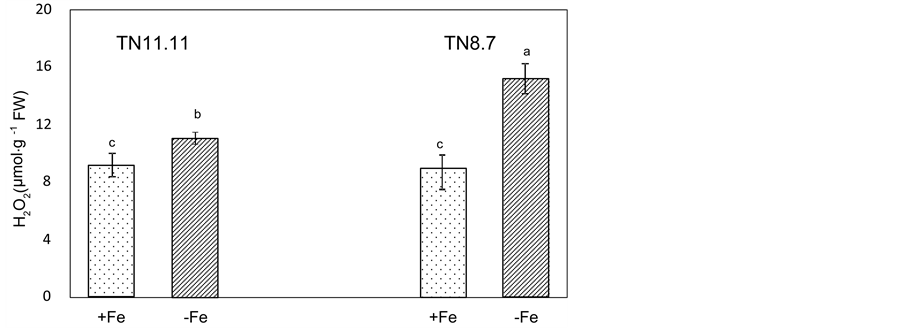
Figure 4. H2O2 content in leaves of two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
Table 3. Enzymatic activity of total superoxide dismutase (SOD) and its different isoforms in two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). One enzymatic unit is defined as the amount of enzyme inhibiting 50% of NBT-photoreduction. Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
KCN-insensitive) was detected only in plants of both genotypes grown in the presence of Fe in the nutrient solution (Table 3).
Data related to POD activity demonstrated a significant decrease under Fe deficiency conditions respect to the control in both lines (−23% and −14% for TN8.7 and TN11.11 respectively). Ascorbic peroxidase exhibited the strongest decrease under Fe deficiency respect to the control in both genotypes, being −45% and −32% in TN8.7 and TN11.11 respectively (Table 4).
3.5. Non-Enzymatic Antioxidant Activities, Ascorbate and Glutathione
Leaf antioxidant activity (FRAP) of the two lines was presented in Table5 Iron deprivation led to a significant FRAP reduction in TN8.7, (−17%) while no variation was noted for TN11.11. The results related to ascorbate (total, ASC and DHA) and glutathione (total, GSH and GSSG) concentrations in both lines under Fe-sufficient and Fe-deficient conditions are summarized in Table5 The results revealed that TN11.11 plants subjected to Fe deprivation exhibited a significant increase in total ascorbate concentration, reaching +85% with respect to the control. The same tendency was observed for reduced ascorbate (ASC) but not for the oxidized form (DHA). Consequently, the ratio ASC/DHA showed a significant increase. However, Fe deficiency had no significant effect neither on the total glutathione concentration nor on its two forms (GSSG and GSH), resulting in a similar GSH/GSSG ratio in both control and treated plants (Table 5).
Table 4. Enzymatic activity of guaiacol peroxidase (POD) and ascorbate peroxidase (APx) in two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). Enzyme activity of G-POD and APx is expressed as µmol guaiacol or ascorbate consumed min−1∙mg∙prot−1, respectively. Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
Table 5. FRAP assay, changes in ascorbate (reduced (ASC) and oxidized (DHA) ascorbate)and glutathione(reduced (GSH) and oxidized (GSSG) glutathione) concentration in leaves of two lines of Medicago ciliaris plants grown on a control nutrient solution (+Fe) or in the absence of iron (−Fe) during the treatment period (10 days). Values followed by different letters are significantly different at P < 0.05 according to Newman-Keuls.
In TN8.7 line, both the total ascorbate concentration and the reduced form significantly decreased while the oxidized form did not change under Fe starvation. Nevertheless, no modification of the ASC /DHA ratio was noted under Fe deficiency stress. Similarly, the total glutathione pool and its reduced form decreased in Fedeficient leaves (respectively −19% and −25% respect to the control), whereas the oxidized form (GSSG) was similar to the control. Also in this case, the GSH/GSSH ratio remained constant (Table 5).
4. Discussion
In the present study, we investigate possible mechanisms responsible for Fe deficiency tolerance at leaf level in two Medicago ciliaris lines differing in their tolerance to Fe deficiency. The genotype TN11.11 is native of a calcareous soil, characterized by its high pH causing Fe deficiency, and is known for its ability to deal with such severe conditions. The genotype TN8.7, colonizing a soil poor in lime content where the pH is lower, is considered to be more sensitive and less efficient in responding to Fe deficiency [10] .
Based on the leaf responses to Fe deficiency, significant differences were observed between the two lines studied. In fact, the majority of these responses, including leaf chlorophyll and Fe concentrations, fluorescence parameters and antioxidative defence were genotype-dependant. Our results showed that the photosynthetic parameters, leaf Fe content and antioxidant system of the sensitive line TN8.7 were more affected than those of the tolerant TN11.11 line by Fe starvation. These recent findings reinforced our previous work focused in root responses of the same Medicago ciliaris lines [10] .
Chlorosis is a phenomenon which appears in young leaves and is often known as the first visual sign of Fe deficiency in plants. It is associated not only with a loss of chlorophyll content, since several steps of its biosynthesis are Fe-depend, but also with changes in the expression and assembly of other components of the photosynthetic apparatus [26] . In our case, both lines of Medicago ciliaris exhibited the classic visual chlorosis symptoms, with a more severe impact in TN8.7 line compared to TN11.11. Such behaviour could be explained by a considerable decrease in chlorophyll and bivalent Fe concentrations, which is particularly strong in the sensitive TN8.7 (Table 1). Photosynthesis is considered as an interesting physiological indicator of the plant tolerance to stress. Under unfavourable conditions, the first response of the plant to an environment challenge is the decrease of the photosynthetic rate and the inhibition of numerous molecular mechanisms. In this study, the chlorophyll fluorescence parameter, which is related to the photosynthetic activity, well reflected the different tolerance to Fe deficiency in the two genotypes, suggesting that it could be usefully employed for rapid screening of iron deficiency tolerance in Medicago ciliaris. According to many authors, this parameter provides useful information about plant photosynthetic performance under abiotic stresses [27] [28] . Our data showed that the maximal potential photochemical efficiency of PSII under dark-adapted conditions (Fv/Fm) and the excitationenergy trapping capacity at the reaction centre of PSII (1/F0 - 1/Fm) were significantly affected by Fe deficiency treatment especially in the sensitive line TN8.7. Our findings agree with those obtained by [3] [29] [30] obtained on other species. According to [3] , the decline in Fv/Fm ratio under Fe deficiency conditions could be explained by the changes in the electron transport chain. In order to verify if the Fv/Fm decrease is associated to electron transport chain through the PSI and PSII, we analyzed the electron transport in isolated thylakoid obtained from chlorotic leaves. We found that the transport of electron through the photosynthetic chain was inhibited by the absence of iron, especially in the sensitive genotype TN8.7. Taken together, these data indicated that Fe deficiency affects the functionality of the photosynthetic machinery, more in the sensitive line TN8.7 than in the tolerant TN11.11.
The impact of Fe deficiency stress on photosynthetic apparatus mainly by impairing the electron transport chain functionality both at the mitochondrion and at the chloroplast level [31] results in the formation of reactive oxygen species (ROS ) leading to the induction of oxidative stress. In the current study, three indicators of oxidative stress MDA, H2O2 and volatile aldehyde content, were examined in the two Medicago ciliaris lines characterized by different tolerance to Fe deficiency.
Aldehydes are intermediates in several fundamental metabolism pathways for carbohydrates, vitamins, steroids, amino acids, and lipids [32] . Such compounds are indispensable for organism life but, excessive amounts threat seed survive because of their chemical reactive nature [33] . They are also produced in response to environmental stresses that disturb plant metabolism, such as salinity, dehydration, desiccation, cold and heat shock [34] . Our results showed that the volatile aldehyde concentration was similar in control plants of the two lines. In the absence of Fe, this parameter increased by 1.13 and 2.17-fold in the Fe-tolerant (TN11.11) and Fe-sensitive (TN8.7) lines respectively. The lack of Fe in the medium induced also a considerable increase of lipid peroxidation, especially in the sensitive line (TN8.7). Taken together, the increase in volatile aldehyde and MDA contents demonstrated that Fe deficiency in the growth medium can cause a higher oxidative stress in the Fe-sensitive line compared to the Fe-tolerant one.
According to [35] , membrane damage might be caused by the production of high H2O2 levels, which could accelerate the Haber-Weiss reaction, resulting in hydroxyl radical (OH-) formation and to lipid peroxidation. Accordingly, the evaluation of H2O2 concentrations showed that this reactive species was strongly enhanced under Fe deficiency conditions, with a highest value registered in TN8.7. The results obtained suggest that Fe starvation enhance some systems responsible for H2O2 production. To check this hypothesis, we analysed the main enzymes involved in the ROS metabolism. Among these enzymes, SODs, which catalyse the conversion of the superoxide radical into molecular oxygen and H2O2, were assayed. This enzymatic activity plays an important role in protecting plants against oxidative damage. There are three types of SODs based on their metal cofactor: Mn-SOD, Fe-SOD and CuZn-SOD. Measurements of these SOD isoforms are useful for assessing the micronutrient status of plants, and have been used to study deficiencies in Zn, Fe, Mn and Cu. In the present work, total SOD activity was induced in Fe-deficient leaves of both lines, but to a lower extent for TN11.11, suggesting a low need for superoxide radical scavenging in this line. Moreover, in both genotypes grown under Fe-limiting conditions leaf Fe-SOD activity was undetectable, whereas total SOD activity recorded an increase. In concomitance with disappearance of Fe-SOD, Fe deficient leaves increased CuZn-SOD and Mn-SOD activities respect to the control. It has been previously suggested that a limitation in the concentration of a micronutrient can depress its corresponding isoform enzyme activity but apparently there is a compensatory mechanism involving the induction of other isoforms, in order to keep an adequate level of SOD for the cell protection against deleterious effects of superoxide radicals. So far, this compensatory mechanism between Mn-SOD and CuZn-SOD has been well described in pea leaves [36] and in tobacco plants grown under limiting Mn conditions [37] [38] , between Mn-SOD and Fe-SOD in Fe deficient pea leaves [39] and between CuZn-SOD and FeSOD in pea leaves conducted under Cu-limiting conditions [40] . Peroxidase enzymes are importantly involved in reducing H2O2 to H2O. In the present study, our results revealed a reduction in POD and even more in APx activities in both lines. This could be explained by the fact that POD contains a haem Fe and APx contains, other than a haem Fe, also a non-haem Fe atom. The lower level of H2O2 accumulated in the tolerant line suggests that this is better protected against oxidative damage caused by Fe starvation than the sensitive one. Non-enzymatic antioxidant activity is represented by a series of antioxidant molecules used by plants against ROS formation [35] . FRAP was used to provide information on total non-enzymatic antioxidant activity. As expected, in TN11.11 line, Fe deficiency increased non-enzymatic antioxidant activity.
Ascorbate (Asc) and glutathione (GSH) are important metabolites in plants. Their most prominent and best established functions are their crucial antioxidant roles in the Asc-GSH cycle [41] . Our results showed that in the tolerant line TN11.11 the total ascorbate content was increased by Fe deprivation. This could be explained by a stimulation of its synthesis or by a diminution of its catabolism and suggests an important role for this antioxidant against oxidative stress provoked by Fe deprivation in TN11.11. The change in the ASC/DHA ratio, an important indicator of the redox status of the cell, is one of the first signs of oxidative stress [42] . The increase in ASC to DHA ratio observed in TN11.11 plants subjected to Fe deprivation could be due to an enhanced sythesis of reduced ascorbate. This is observed just in the tolerant line, suggesting that the metabolism is shifted towards reductive pathways. Concerning glutathione, our results showed no effect of Fe deficiency treatment on the total glutathione content, GSH, GSSG, and GSH/GSSG ratio indicating that glutathione metabolism was unaffected. Regarding the susceptible line TN8.7, ASC significantly decreased in iron-deficient leaves, probably because of the H2O2- mediated oxidation of ASC, while DHA remained unchanged. As suggested by [43] and [44] , the ASC pool could be lowered by oxidative stress when the biosynthesis capacity is exceeded. Also the content of GSH was significantly decreased in TN8.7 under Fe deficiency, which was probably due to the decrease in the activities of some enzymes of this cycle such as GR activity.
5. Conclusion
According to the literature, many studies are focused on Fe deficiency tolerance in plant. Nevertheless, there are no well-defined plant indicators for tolerance to this abiotic stress at leaf level. This is the first paper studying the biochemical (MDA, enzymatic and non-enzymatic antioxidant system) and physiological (photosynthetic electron transport of the whole chain and the chlorophyll a fluorescence parameters) traits in Fe-stressed Medicago ciliaris leaves. Our results showed significant changes in these parameters in both lines considered: TN11.11, which is tolerant to Fe deficiency and TN8.7 which is susceptible. Only the tolerant line TN11.11 showed an efficient enzymatic and non-enzymatic antioxidant defence system in order to limit ROS production under Fe deficiency stress and consequently to prevent over reduction of the photosynthetic electron transport chain. The main output of this study is that the parameters here considered can be used as precious criteria for predicting plant performance under Fe deficiency stress and for screening tolerant genotypes at leaf level.
Acknowledgements
This work was supported by grants from the Tunisian Ministry of Higher Education, Scientific Research (LR10- CBBC02).
References
- Mengel, K. (1994) Iron Availability in Plant Tissues-Iron Chlorosis in Calcareous Soils. Plant Soil, 165, 275-283. http://dx.doi.org/10.1007/BF00008070
- Nedunchezhain, N., Morales, F., Abadia, A. and Abadia, J. (1997) Decline in Photosynthetic Electron Transport Activity and Changes in Thylakoid Protein Pattern in Field Grown Iron Deficient Peach (Prunus persica L.). Plant Science, 129, 29-38. http://dx.doi.org/10.1016/S0168-9452(97)00170-2
- Donnini, S., Castagna, A., Guidi, L., Zocchi, G. and Ranieri, A. (2003) Leaf Responses to Reduced Iron Availability in Two Tomato Genotypes: T3238FER (Iron Efficient) and T3238fer (Iron Inefficient). Journal of Plant Nutrition, 26, 2137-2148. http://dx.doi.org/10.1081/PLN-120024270
- Asada, K. (1999) The Water-Cycle in Chloroplasts: Scavenging of Active Oxygens and Dissipation of Excess Photons. Annual Review of Plant Physiology and Plant Molecular Biology, 50, 601-639. http://dx.doi.org/10.1146/annurev.arplant.50.1.601
- Azevedo Neto, A.D., Prisco, J.T., Enéas-Filho, J., E. Braga de Abreu, C. and Gomes-Filho, E. (2006) Effect of Salt Stress on Antioxidative Enzymes and Lipid Peroxidation in Leaves and Roots of Salt-Tolerant and Salt-Sensitive MAIZE genotypes. Environmental and Experimental Botany, 56, 87-94. http://dx.doi.org/10.1016/j.envexpbot.2005.01.008
- Kubo, A., Saji, H., Tanaka, K. and Kondo, N. (1992) Cloning and Sequencing of a cDNA Encoding Ascorbate Peroxidase from Arabodopsis thaliana. Plant Molecular Biology, 18, 691-701. http://dx.doi.org/10.1007/BF00020011
- Noctor, G. and Foyer, C. (1998) Ascorbate and Glutathione: Keeping Active Oxygen under Control. Annual Review of Plant Physiology and Plant Molecular Biology, 49, 249-279.
- M’sehli, W., Dell’Orto, M., De Nisi, P., Donnini, S., Abdelly, C., Zocchi, G and Gharsalli, M. (2009) Responses of Two Ecotypes of Medicago Ciliaris to Direct and Bicarbonate-Induced Iron Deficiency Conditions. Acta Physiologiae Plantarum, 31, 667-673. http://dx.doi.org/10.1007/s11738-009-0288-1
- M’sehli, W., Dell’Orto, M., Donnini, S., De Nisi, P., Zocchi, G., Abdelly, C. and Gharsalli, M. (2009) Variability of Metabolic Responses and Antioxidant Defence in Two Lines of Medicago ciliaris to Fe Deficiency. Plant and Soil, 32, 219-230. http://dx.doi.org/10.1007/s11104-008-9887-7
- M’sehli, W., Youssfi, S., Donnini, S., Dell’Orto, M., De Nisi, P., Abdelly, C. and Gharsalli, M. (2008) Root Exudation and Rhizosphere Acidification by Two Lines of Medicago ciliaris in Response to Lime-Induced Iron Deficiency. Plant and Soil, 312, 151-162. http://dx.doi.org/10.1007/s11104-008-9638-9
- Oserkowsky, J. (1933) Quantitative Relation between Chlorophyll and Iron in Green and Chlorotic Leaves. Plant Physiology, 8, 449-468. http://dx.doi.org/10.1104/pp.8.3.449
- Llorente, S., Leon, A., Torrecillas, A. and Alcaraz, C. (1976) Leaf Iron Fractions and Their Relation with Iron in Citrus. Agrochimica, 20, 205-212.
- Torrecillas, A., Léon, A., Del Amor, F. and Martinez-Mompean, M.C. (1984) Rapid Determination of Chlorophyll. Fruits, 39, 617-622.
- Ferraro, F., Castagna, A., Soldatini, G. and Ranieri, A. (2003) Influence of Different Iron Concentrations on Thylakoid Pigment and Protein Composition. Plant Science, 164, 783-792. http://dx.doi.org/10.1016/S0168-9452(03)00065-7
- Hammami, S., Chaffai, R. and El Ferjani, E. (2004) Effect of Cadmium on Sunflower Growth, Leaf Pigment and Photosynthetic Enzymes. Pakistan Journal of Biological Sciences, 7, 1419-1426. http://dx.doi.org/10.3923/pjbs.2004.1419.1426
- Jana, S. and Choudhuri, M.A. (1981) Glycolate Metabolism of Three Submerged Aquatic Angiosperms during Aging. Aquatic Botany, 12, 345-354. http://dx.doi.org/10.1016/0304-3770(82)90026-2
- Peixoto, P.H.P., Cambraia, J., Sant’Anna, R., Mosquim, P.R. and Moreira, M.A. (1999) Aluminuim Effects on Lipid Peroxidation and on the Activities of Enzymes of Oxidative Metabolism in Sorghum. Revista Brasileira de Fisiologia Vegetal, 11, 137-143.
- Cakmak, I. and Horst, J.H. (1991) Effects of Aluminium on Lipid Peroxidation, Superoxide Dismutase, Catalase, and Peroxidase Activities in Root Tips of Soybean (Glycine Max). Physiologia Plantarum, 83, 463-468.http://dx.doi.org/10.1111/j.1399-3054.1991.tb00121.x
- Ranieri, A., D’Urso, G., Nali, G., Lorenzini, G. and Soldatini, G.F. (1996) Ozone Stimulates Apoplastic Systems in Pumkin Leaves. Plant Physiology, 97, 381-387.
- Donahue, J.L., Okpodu, C.M., Cramer, C.L., Grabau, E.A. and Alscher, R.G. (1997) Responses of Antioxidants to Paraquat in Pea Leaves. Plant Physiology, 113, 249-257.
- Griffith, O.W. (1980) Determination of Glutathione Disulphide Using Glutathione Reductase in Leaves of Pea (Pisum sativum L.). Planta, 180, 278-284.
- Miyake, C. and Asada, K. (1992) Thylakoid-Bound Ascorbate Peroxidase in Spinach Chloroplasts and Photoreduction of Its Primary Oxidation Product Monodehydroascorbate Radicals in Thylakoids. Plant and Cell Physiology, 33, 541-553.
- Benzie, I.E.F. and Strain, J.J. (1996) The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Analytical Biochemistry, 239, 70-76. http://dx.doi.org/10.1006/abio.1996.0292
- Rosales, M.A., Ruiz, J.M., Hernández, J., Soriano, T., Castilla, N. and Romero, L. (2006) Antioxidant Content and Ascorbate Metabolism in Cherry Tomato Exocarp in Relation to Temperature and Solar Radiation. Journal of the Science of Food and Agriculture, 86, 1545-1551. http://dx.doi.org/10.1002/jsfa.2546
- Kampfenkel, K., Vanmontagu, M. and Inze, D. (1995) Extraction and Determination of Ascorbate and Dehydroascorbate from Plant Tissue. Analytical Biochemistry, 255, 165-167. http://dx.doi.org/10.1006/abio.1995.1127
- Donnini, S., Castagna, A., Ranieri, A. and Zocchi, G. (2009) Differential Responses in Pear and Quince Genotypes Induced by Fe Deficiency and Bicarbonate. Journal of Plant Physiology, 166, 1181-1193.http://dx.doi.org/10.1016/j.jplph.2009.01.007
- Longenberger, P.S., Smith, C.W., Duke, S.E. and McMichael, B.L. (2009) Evaluation of Chlorophyll Fluorescence as a Tool for the Identification of Drought Tolerance in Upland Cotton. Euphytica, 166, 25-33.http://dx.doi.org/10.1007/s10681-008-9820-4
- Brestic, M. and Zivcak, M. (2013) PSII Fluorescence Techniques for Measurement of Drought and High Temperature Stress Signal in Crop Plants: Protocols and Applications. In: Rout, G.R. and Das, A.B., Eds., Molecular Stress Physiology of Plants, Springer India, New Delhi, 87-131.
- Belkhodja, R., Morales, F., Quilez, R., Lopez-Millan, A.F., Abadia, A. and Abadia, J. (1998) Iron Deficiency Causes Changes in Chlorophyll Fluorescence Due to the Reduction in the Dark of the Photosystem II Acceptor Side. Photosynthesis Research, 25, 173-185.
- Jelali, N., Salah, I.B., M’sehli, W., Donnini, S., Graziano, Z. and Gharsalli, M. (2011) Comparison of Tree Pea Cultivars (Pisum sativum) Regarding Their Responses to Direct and Bicarbonate-Induced Iron Deficiency. Scientia Horticulturae, 129, 548-553.
- Pascal, N. and Douce, R. (1993) Effect of Iron Deficiency on the Respiration of Sycamore (Acer pseudoplatanus L.) Cells. Plant Physiology, 103, 1329-1338.
- Yoshida, A., Rzhetsky, A., Hsu, L.C. and Chang, C. (1998) Human Aldehyde Dehydrogenase Gene Family. European Journal of Biochemistry, 251, 549-557.
- Lindahl, R. (1992) Aldehyde Dehydrogenases and Their Role in Carcinogenesis. Critical Reviews in Biochemistry and Molecular Biology, 27, 283-335. http://dx.doi.org/10.3109/10409239209082565
- Bartels, D. (2001) Targeting Detoxification Pathways: An Efficient Approach to Obtain Plants with Multiple Stress Tolerance? Trends in Plant Science, 6, 284-286. http://dx.doi.org/10.1016/S1360-1385(01)01983-5
- Mittler, R. (2002) Oxidative Stress, Antioxidants and Stress Tolerance. Trends in Plant Science, 7, 405-410.http://dx.doi.org/10.1016/S1360-1385(02)02312-9
- Del Río, L.A., Sevilla, F., Gómez, M., Yañez, J. and López-Gorge, J. (1978) Superoxide Dismutase: An Enzyme System for the Study of Micronutrient, Interactions in Plants. Planta, 140, 221-225. http://dx.doi.org/10.1007/BF00390251
- Yu, Q. and Rengel, Z. (1999) Micronutrient Deficiency Influences Plant Growth and Activities of Superoxide Dismutases in Narrow-Leafed Lupins. Annals of Botany, 83, 175-182. http://dx.doi.org/10.1006/anbo.1998.0811
- Randall, P. and Bouma, D. (1973) Zinc Deficiency, Carbonic Anhydrase, and Photosynthesis in Leaves of Spinach. Plant Physiology, 52, 229-232. http://dx.doi.org/10.1104/pp.52.3.229
- Iturbe-Ormaetxe, I., Moran, F., Arrese-Igor, C., Gogorcena, Y., Klucas, R. and Becana, M. (1995) Activated Oxygen and Antioxidant Defences in Iron-Deficient Pea Plants. Plant, Cell & Environment, 18, 421-429.http://dx.doi.org/10.1111/j.1365-3040.1995.tb00376.x
- Del Río, L.A., Sevilla, F., Sandalio, L. and Palma, J. (1991) Nutritional Effect and Expression of SODs: Induction and Gene Expression; Diagnostics; Prospective Protection against Oxygen Toxicity. Free Radical Research, 13, 819-827.http://dx.doi.org/10.3109/10715769109145863
- Asada, K. and Takahashi, M. (1987) Photoinhibition Production and Scavenging of Activated Oxygen. In: Kyle, D.J, Osmond, C.B. and Arntzen, C.J., Eds., Photoinhibition, Elsevier Science Publishers, Amsterdam, 227-287.
- Meneguzzo, S., Navari-Izzo, F. and Izzo, R. (1999) Antioxidative Responses of Shoots and Roots of Wheat to Increasing NaCl Concentrations. Journal of Plant Physiology, 155, 274-280. http://dx.doi.org/10.1016/S0176-1617(99)80019-4
- Foyer, C.H., Lelandais, M. and Kunert, K.J. (1994) Photooxidative Stress in Plants. Physiologia Plantarum, 92, 696-717. http://dx.doi.org/10.1111/j.1399-3054.1994.tb03042.x
- Smirnoff, N. and Pallanca, E. (1996) Ascorbate Metabolism in Relation to Oxidative Stress. Biochemical Society Transactions, 24, 472-478.
NOTES

*Corresponding author.
#These authors contributed equally to this work.