Neuroscience & Medicine
Vol.2 No.2(2011), Article ID:5519,6 pages DOI:10.4236/nm.2011.22013
5-HT4 Receptor Agonists for the Treatment of Alzheimer’s Disease
![]()
Suven Life Sciences Limited, Hyderabad, India.
Email: nvsrk@suven.com
Received February 15th, 2010; revised March 16th, 2011; accepted March 23rd, 2011.
Keywords: 5-HT4 Receptor, Cognition, Neurogenesis, Alzheimer’s Disease
ABSTRACT
Alzheimer’s disease (AD) is a progressive neurological disorder primarily affecting new memory formation as well as retrieval of previously acquired memories. According to World Health Organization, current global population suffering from cognitive impairment is estimated to 37 million. The number is projected to double in next one and half decade. Half of the population afflicted with dementia is represented by AD patients. Current therapies, which provide marginal symptomatic relief to AD patients, are effective only in half of the patient population. In depth understanding of the molecular mechanism of the disease is urgently required to develop more effective therapies. Therapies in clinical development may either offer symptomatic relief to patients or provide pure disease modifications, thus limiting benefit to patients. 5-HT4 receptor agonists offer an attractive option for the treatment of AD patients. Activation of 5-HT4 receptor under preclinical conditions is demonstrated to improve neurotransmission and enhance the release of acetylcholine resulting in the memory formation. In various cell based and animal models, partial 5-HT4 receptor agonists are demonstrated to promote the release of soluble amyloid precursor protein alpha and block the release of amyloid beta peptide offering suitable candidates as disease modification agents. Remarkably, 5-HT4 receptor agonists are also reported to induce neurogenesis in hippocampus as well as enteric system through the activation of cyclic AMP response element binding protein in rodents. Taken together, 5-HT4 agonists address all major facets of Alzheimer’s disease and may provide therapeutic potential for other neurological disorders.
1. Introduction
5-HT4 receptor is a G protein coupled receptor (GPCR) which belongs to serotonin receptor family and is coupled to G protein containing Gas subunit [1]. The receptor, upon activation by an agonist, leads to the generation of intracellular cyclic AMP (cAMP) which in turn activates Protein kinase A. A cascade of signaling events result in the phosphorylation of cAMP response element binding protein (CREB) which binds to its response element leading to the expression of a number of genes involved in cell survival. Many excellent review articles were published recently highlighting the role of 5-HT4 receptor agonists for treatment of gastro-intestinal (GI) spectrum of diseases [2-4]. Contributions of 5-HT4 receptor along with other 5-HT receptors in learning and memory were also reviewed recently in a series of in depth articles [5,6]. An impressive article highlighting the role of 5-HT4 receptor in AD was written with specific emphasis on beta amyloid peptide secretion [7].
In the present article, we focus on the role of 5-HT4 receptor agonists in the treatment of AD with current understanding of their role in cognitive improvement, disease modification and neurogenesis. We further list enormous challenges that need to be overcome to realize the potential of the target and offer possible future directions that may help in overcoming some of the challenges.
2. Receptor Splice Variants
A larger number of 5-HT4 receptor splice variants were identified both in human and rodents with no significant splice specific tissue expression [8]. Figure 1 provides an image of various 5-HT4 receptor splice variants based on sequence comparison. All the identified human splice variants exhibited identical amino acid sequence up to Leucine 358 with differences emerging in the short carboxyl terminal tail [9]. Such an observation led to the conclusion that ligand binding properties of various receptor isoforms may not differ, as entire extracellular and transmembrane domain remained conserved across various receptor subtypes. Indeed 5-HT as well as 5-HT4 receptor antagonist GR-113808 did not demonstrate any significant differences in their binding affinity to human 5-HT4(a) and 5-HT4(b) receptors [10]. In the same set of studies, a number of compounds were evaluated for their ability to induce generation of cAMP and they all showed similar functional affinities for both the receptor variants. Activation of 5-HT4 receptor was reported to induce influx of calcium ions in the host cell by blocking the potassium channels. Surprisingly, a differential effect in calcium mobilization was observed with various compounds acting through 5-HT4(a) receptor but not through 5-HT4(b) receptor [10]. However, 5-HT did not exhibit any differential effect on the calcium mobilization between two receptor variants. Above observations suggested interaction of diverse effector proteins with different receptor subtypes as expected from the diversity of the carboxyl terminal tail. Furthermore, it appears that interaction of an effector protein with the receptor may be modulated by the type of ligands and their mode of interactions.
However, it was also reported that a partial 5-HT4 receptor agonist SL65.0155 as well as 5-HT showed significant differences in their functional affinities to human 5-HT4(b) and 5-HT4(e) receptors [11]. Above observation suggested that carboxyl terminal tail of the 5-HT4 receptor variants may modulate the affinity of certain ligands to the receptor apart from its ability to differentially transmit the intracellular signaling. Such a differential affinity of a ligand for various 5-HT4 receptor variant may be more prominent between specific variants.
3. Cognitive Enhancement
Positive impact of cAMP in acquisition and consolidation of memory is well accepted. Activation of CREB protein by cAMP dependent protein kinase is an important mediator of memory formation [12,13]. A number of signaling pathways leading to cAMP accumulation in neurons are being explored as possible candidates for therapeutic interventions of cognitive deficits associated with various neurological disorders.
5-HT4 receptor is expressed at high level in limbic system of CNS. The receptor is coupled to G protein containing Gas subunit [1]. Thus, activation of the receptor by an agonist leads to cAMP formation which through CREB phosphorylation is proposed to help in new memory formation. CREB mediated memory formation is likely mediated through the expression of brain derived neurotrophic factor (BDNF) and other trophic and procognitive factors. An illustration of 5-HT4 receptor’s role in memory formation is provided in Figure 2. In addition, activation of the 5-HT4 receptor is proposed
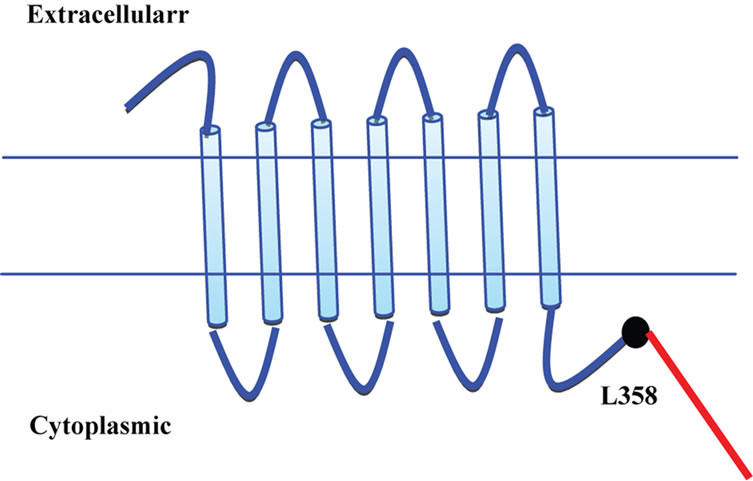
Figure 1. 5-HT4 receptor splice variants. The human 5-HT4 receptor sequence is conserved up to leucine 358 with differences appearing at amino acid 359 onwards depicted in red. Sequence of different variants downstream of Leu358 is shown. The blue line represents the conserved sequence with light blue barrels as the transmembrane domain. 5-HT4(a) RYTVLHRGHHQELEKLPIHNDPESLESCF; 5- HT4(b) RDAVECGGQWESQCHPPATSPLVAAQPSDT; 5- HT4(c)SSGTETDRKKLWNKEEKIDQTIQMPKRKRKKKASLSYEDLILLGRKSCFREGK; 5-HT4(d) RF; 5-HT4(e) SFPLLFCNRPVPV; 5-HT4(f) SPVPV; 5-HT4(g) SGCSPVSSFLLLFCNRPVPV.
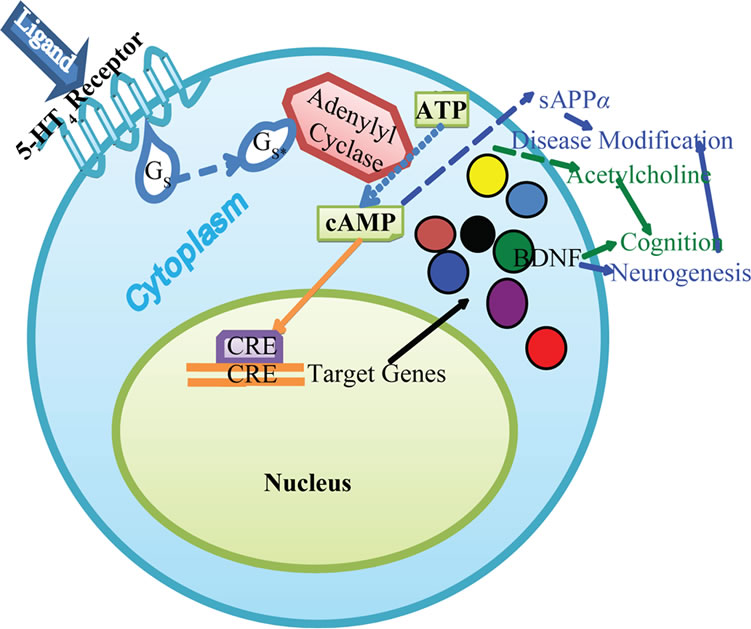
Figure 2. A cartoon representation of 5-HT4 receptor activation leading to various cellular events. Activation of 5- -HT4 receptor leads to acetylcholine release, which coupled with the release of BDNF, may help in memory formation. The activation of the receptor is also reported to enhance the release of sAPPa, which along with BDNF-induced neurogenesis offers disease modifying potential for AD patients.
to facilitate the release of various neurotransmitters by blocking potassium channels and subsequent mobilization of calcium ions. Indeed a number of 5-HT4 receptor agonists were reported to promote learning tasks in various animal models. 5-HT4 receptor partial agonist SL- 65.0155 demonstrated pro-cognitive effect in a Novel Object Recognition Task (NORT) in a dose dependent manner. The compound also reversed scopolamine-induced amnesia in rats when evaluated in a water maze assay. The pro-cognitive effect of the compound was blocked by 5-HT4 receptor antagonist SDZ 205,557 in the NORT assay [11]. SL65.0155 was also reported to reverse amnesia in rodents induced by diverse class of agents such as amyloid beta peptide 1 - 42, carbon monoxide and methylazoxymethanol acetate suggesting that the target offers a great therapeutic potential [14]. In a model of attention, five choice serial reaction time task, SL65.0155 reduced incorrect choices and promoted correct choices [15]. This observation suggested that compounds developed through 5-HT4 receptor may also find therapeutic utility in the treatment of ADHD patients apart from their potential in therapeutic interventions in cognitive deficits associated with AD.
In a passive avoidance paradigm, icv injection of SC 53116, a 5-HT4 receptor agonist reversed scopolamine induced amnesia [16]. A number of studies were reported in various international conferences with a 5-HT4 partial agonist PRX03140 (formerly VRX03011). The compound reached up to Phase IIb of clinical development for the treatment of AD. PRX-03140 demonstrated high functional affinity to various human 5-HT4 receptor splice variants with no apparent binding to other serotonin receptors. The compound demonstrated activity in a four arm cross maze assay in delayed mode response [17]. In a recently published study, various 5-HT4 receptor agonists were demonstrated to reverse scopolamine induced amnesia in a Water Maze paradigm [18]. The dose dependent effect of the ligands was blocked by selective 5-HT4 receptor antagonist GR125487. Taken together, all the above studies and reports provide a strong support for the development of 5-HT4 receptor agonists as therapeutic agents for the treatment of various dementia related disorders.
4. Neurotransmitter Release
In order to better understand the mode of action of 5-HT4 receptor agonists leading to memory formation, specific and targeted studies were conducted using various neurochemistry and electrophysiology based approaches. Role of 5-HT4 receptor in synaptic plasticity was demonstrated in freely moving rats with implanted microelectrodes. Activation of the receptor by specific agonist RS67333 led to neurotransmission in hippocampus [19]. Similar neurotransmission was demonstrated in CA1 region of hippocampus in anesthetized rats using another 5-HT4 selective agonist SC 53116 [16]. As mentioned earlier, activation of 5-HT4 receptor leads to the release of various neurotransmitters mediated through calcium influx, as a result of blockade of potassium channels. A 5-HT4 receptor agonist, methoxytryptamine was reported to increase the level of acetylcholine in prefrontal cortex of rats. It was further demonstrated that PRX-03140 enhanced the release of acetylcholine upon trial challenge in hippocampus of rats but not during resting period [17]. One of the mechanistic roles of extracellular acetylcholine is to enhance cholinergic neurotransmission which plays central role in memory formation. Thus, 5-HT4 receptors may improve memory formation by enhancing the synaptic release of acetylcholine in the brain which would in turn enhance the cholinergic transmission.
5. APP Processing
A major proposed advantage of using 5-HT4 receptor agonist for the treatment of AD patients is its ability to shift the equilibrium of amyloid precursor protein (APP) processing from amyloidogenic to non-amyloidogenic form [7, Figure 2]. Ab peptide either in soluble oligomer or aggregated multimer format in the brain is a major contributing factor of AD pathology [20]. The peptide is generated by the action of beta and gamma secretases which either produce 42 or 40 amino acid peptides [21]. Both these peptides are amyloidogenic and implicated in the progression of the disease with 42 amino acid peptide having higher potential to aggregate. Another enzyme reported to cleave APP within the amyloidogenic peptide sequence is less known alpha secretase [21,22]. Activation of the alpha secretase pathway is shown to shift the balance of APP cleavage from amyloidogenic to nonamyloidogenic form. The first product of alpha secretase pathway is soluble APP alpha (sAPPa) protein. sAPPa acts as a neurotrophic factor and helps in neuronal survival. The protein gets further cleaved by other proteases and eliminated.
Major efforts to develop disease modifying drugs for AD treatment focused on identifying potent and selective inhibitors of beta or gamma secretases [23,24]. The blockade of these enzymes would not only prevent generation of amyloidogenic form of the peptide but may also shift the equilibrium of APP processing towards alpha secretase pathway. Highly involved effort for the identification of potent and selective beta secretase inhibitors was hampered due to large and complex substrate binding pocket of the enzyme [24]. In addition, compounds which demonstrated acceptable in vitro properties exhibited poor pharmacokinetic and brain penetration profile. Development of gamma-secretase inhibitor was more successful with molecules reaching up to phase III of clinical development [23]. As gamma secretase was reported to cleave other critical signaling proteins, development of compounds which block the APP processing activity without affecting the critically required enzyme action on other substrates had offered initial challenge which was overcome with smart chemistry efforts [23, 25].
Activation of alpha secretase pathway, despite offering distinct advantages as a therapeutic target for AD treatment, was overlooked for too long [22]. Such reluctance to develop alpha-secretase activator was probably due to strong and valid historical reason. It is much easier to identify a compound which can bind to the active site of an enzyme and inhibit its activity. For synthesis of an enzyme activator, an allosteric site needs to be first characterized which can bind to specific compounds. To overcome the challenge of an allosteric site, activation of an upstream signaling pathway may be exploited to enhance the enzyme activity. Agonism of 5-HT4 receptor is reported to activate alpha-secretase pathway offering an attractive mechanism to develop new therapy for AD (Figure 2). The shifted equilibrium of APP processing from amyloidogenic to non-amyloidogenic form generates sAPPa which is reported to provide neuroprotective effect. Activation of alpha-secretase pathway by 5-HT4 agonists was first reported in a recombinant as well as a neuronal cell line [26] and subsequently demonstrated in other cell lines as well as in mouse brain [10,27]. Treatment with 5-HT4 receptor agonist RS67333 is also reported to inhibit the generation of Ab peptide from the cortical neurons of Tg 2576 mice under culture conditions [28].
Most of the investigators utilized 5-HT4(e) receptor variant for the study of APP processing under artificial conditions [17,26,29]. However, 5-HT4(d) receptor activation was also demonstrated to enhance sAPPa level with concomitant decrease in Ab peptide level [7,18]. It is not yet clear whether other variants of 5-HT4 receptor would also exhibit similar kind of differential APP processing. 5-HT4(e) receptor is not restricted to CNS but is expressed in other peripheral tissues [8]. In addition as mentioned earlier, all differences in various 5-HT4 receptor variants are restricted at the carboxy terminal cytoplasmic domain while their extracellular and transmembrane domains are absolutely conserved [9]. Majority of the compounds are not expected to show significant differences in their binding affinity to various receptor variants based on absolute conservation of the sequence in the ligand binding domain. However, due to differences in the carboxyl terminal domain of the receptor, these variants may interact with distinct or overlapping signaling machinery leading to differential intracellular responses. Blockade of specific 5-HT4 activated pathway is reported to reduce the sAPPa level in the medium under cell culture conditions [28]. Despite identification of potent and selective 5-HT4 receptor agonists long ago [30], their development for CNS indications moved at a slow pace. Potential impedance for the development of such compounds for neurological disorders could likely be their poor pharmacokinetic and brain penetration profile coupled with a concern for cardiovascular safety. The safety concern was further compounded due to withdrawal of marketed drugs acting through this target for their cardiovascular liability. However, most of the compounds which were withdrawn from the market demonstrated poor selectivity and their cardiovascular liability could have been independent of the target. Indeed, few 5-HT4 agonists are still in market for the treatment of GI conditions. In addition, Prucalopride, a selective 5-HT4 receptor agonist with excellent cardiovascular safety profile from Movetis is already approved for treatment of chronic constipation in women in Europe offering a proof of concept for the target.
6. Neurogenesis
One of the most remarkable features of 5-HT4 receptor agonists is their ability to induce neurogenesis in hippocampus as well as enteric system in rodents [31,32]. This feature of 5-HT4 receptor agonists truly offers disease modifying potential, as the compounds may be able to replace the degenerated cells by inducing production of new neurons (Figure 2). Partial 5-HT4 receptor agonist RS67333 demonstrated capacity to induce neurogenesis in hippocampus of treated rats resulting in faster onset of the antidepressant activity [32]. Due to abundant expression of 5-HT4 receptor in enteric system, neurogenesis mediated by the receptor was investigated extensively in enteric system. A number of 5-HT4 receptor agonists are reported to induce the survival of enteric neurons and induce the neurite outgrowth under culture conditions which can be blocked by 5-HT4 receptor specific antagonist [31]. In the same study, incorporation of bromo-deoxyuridine was reported in vivo in enteric cells expressing neuronal markers upon treatment with 5-HT4 receptor agonists. Mouse model with targeted disruption of 5-HT4 gene was explored to investigate any role of the receptor in neurogenesis. While the number of neurons in the enteric system were comparable between wild type and gene knock out mice at birth, the number of neurons declined over a period of time in the mutant mice [31]. Above observations strongly support the role of 5-HT4 receptor and its agonists in neurogenesis in brain and peripheral tissues.
7. Major Challenges
5-HT4 receptor agonists are used in clinic for the treatment of various GI indications. Some of the drugs targeting 5-HT4 receptor such as Cisapride were withdrawn from the market due to cardiovascular liability. As the receptor is also expressed in heart tissue, a major and justifiable concern of on-target cardiovascular liability of 5-HT4 receptor agonists is valid. However, more selective compounds targeting 5-HT4 receptor are still in market. Prucalopride, a selective 5-HT4 receptor agonist with excellent cardiovascular safety profile is approved in Europe for treatment of chronic constipation in women. Above molecules offer a clinical proof of concept for exploration of 5-HT4 receptor agonists as drugs. A partial 5-HT4 agonist PRX-03140 advanced to phase II of clinical development for the treatment of AD. However, overall development of 5-HT4 receptor agonists for CNS disorders was likely hampered due to poor pharmacokinetic profile or inadequate brain penetration of the molecules. While 5-HT4 partial agonists RS67333 and SL65.0155 were identified more than a decade ago and demonstrated excellent efficacy in various animal models, they could not be developed as drugs possibly due to selectivity or cardiovascular liabilities. RS67333 exhibited strong binding to sigma receptors [30]. SL65.0155 reached to phase II of clinical development but was abandoned for undisclosed reasons. Another major challenge was to develop molecules which offer strong neurological effect with minimum activity at GI level. Identification of CNS specific molecules is highly challenging as most of the receptor variants show common expression pattern in various tissues. For example, there is no CNS specific 5-HT4 receptor isoform.
8. Future Direction
Majority of the 5-HT4 receptor ligands developed for GI indications (Cisapride, Tegaserod, Prucalopride etc.) were full agonist whereas compounds with partial agonism to the receptor (PRX-03140, RS67333 and SL 65.0155) exhibited remarkable CNS effect in various animal models. RaQualia (www.raqualia.com) is developing a 5-HT4 receptor partial agonist RQ00000009 for treatment of AD. The compound has completed human Phase I clinical trial and ready to move to Phase II. Identification of a partial agonist with acceptable oral absorption, good pharmacokinetic profile and high brain penetration would be a significant start in this direction. Such molecules are expected to have minimal GI effect as the equilibrium of drugs would shift from systemic circulation to the CNS tissue.
5-HT4 receptor agonists appear to recapitulate all desired features of an ideal treatment for AD patients. Based on their procognitive properties coupled with neurogenesis, they may offer potential treatment for other neurodegenerative diseases. Partial 5-HT4 receptor agonists also offer excellent differentiation from current SSRI based treatment for depression by having quick onset of action [32].
9. Acknowledgements
We would like to thank Suven management for their support and our colleagues for their scientific input. Constructive comments from anonymous referees in improving the article are greatly appreciated.
REFERENCES
- C. Gerald, N. Adham, H.-T. Kao, M. A. Olsen, T. M. Laz, L. E. Schechter, J. A. Bard, P. J.-J. Vaysse, P. R. Hartig, T. A. Branchek and R. L. Weinshank, “The 5-HT4 Receptor: Molecular Cloning and Pharmacological Characterization of Two Splice Variants,” The EMBO Journal, Vol. 14, No. 12, June 1995, pp. 2806-2815.
- H. S. Kim, “5-Hydroxytryptamine4 Receptor Agonists and Colonic Motility,” Journal of Smooth Muscle Research, Vol. 45, No. 1, 2009, pp. 25-29. doi:10.1540/jsmr.45.25
- G. J. Sanger, “Translating 5-HT Receptor Pharmacology,” Neurogastroenterology Motility, Vol. 21, No. 12, December 2009, pp. 1235-1238.
- J. Tack, “Prucalopride: A New Drug for the Treatment of Chronic Constipation,” Expert Review in Gastroenterology and Hepatology, Vol. 3, No. 4, August 2009, pp. 337- 343. doi:10.1586/egh.09.38
- M. V. King, C. A. Marsden and K. C. Fone, “A Role for the 5-HT(1A), 5-HT4 and 5-HT6 Receptors in Learning and Memory,” Trends in Pharmacological Sciences, Vol. 29, No. 9, September 2008, pp. 482-492. doi:10.1016/j.tips.2008.07.001
- G. Perez-Garcia and A. Menesses, “A Memory Formation, Amnesia, Improved Memory and Reversed Amnesia: 5-HT Role,” Behavioural Brain Research, Vol. 195, No. 1, December 2008, pp. 17-29.
- S. J. Robert and F. Lezoualc’h, “Distinct Functional Effects of Human 5-HT4 Receptor Isoforms on Beta-Amyloid Secretion,” Neurodegenerative Diseases, Vol. 5, No. 3-4, March 2008, pp. 163-165. doi:10.1159/000113691
- I. M. Coupar, P. V. Desmond and H. R. Irving, “Human 5-HT4 and 5-HT7 Receptor Splice Variants: Are They Important,” Current Neuropharmacology, Vol. 5, No. 4, December 2007, pp. 224-231. doi:10.2174/157015907782793621
- M. Langlois and R. Fischmeister, “5-HT4 Receptor Ligands: Applications and New Prospects,” Journal of Medicinal Chemistry, Vol. 46, No. 3, January 2003, pp. 319- 344. doi:10.1021/jm020099f
- A. Pindon, G. V. Hecke, P. V. Gompel, A. S. Lesage, J. E. Leysen and M. Jurzak, “Differences in Signal Transduction of Two 5-HT4 Receptor Splice Variants: Compound Specificity and Dual Coupling with Gas and Gai/o-Proteins,” Molecular Pharmacology, Vol. 61, No. 1, January 2002, pp. 85-96. doi:10.1124/mol.61.1.85
- P. C. Moser, O. E. Bergis, S. Jegham, A. Lochead, E. Duconseille, J.-P. Terranova, D. Caille, I. Berque-Bestel, F. Lezoualc’h, R. Fischmeister, A. Dumuis, J. Bockaert, P. George, P. Soubrie and B. Scatton, “SL65.0155, a Novel 5-Hydroxytryptamine 4 Receptor Partial Agonist with Potent Cognition-Enhancing Properties,” Journal of Pharmacology and Experimental Therapeutics, Vol. 302, No. 2, August 2002, pp. 731-741. doi:10.1124/jpet.102.034249
- J.-H. Han, S. A. Kushner, A. P. Yiu, C. J. Cole, A. Matynia, R. A. Brown, R. L. Neve, J. F. Guzowski, A. J. Silva and S. A. Josselyn, “Neuronal Competition and Selection during Memory Formation,” Science, Vol. 316, No. 5823, April 2007, pp. 457-460. doi:10.1126/science.1139438
- M. Xia, R. Huang, V. Guo, N. Southall, M.-H. Cho, J. Inglese, C. P. Austin and M. Nirenberg, “Identification of Compounds that Potentiate CREB Signaling as Possible Enhancers of Long-Term Potentiation,” Proceedings of National Academy of Science USA, Vol. 106, No. 7, February 2009, pp. 2412-2417. doi:10.1073/pnas.0813020106
- V. Micale, G. M. Leggio, C. Mazzola and F. Drago, “Cognitive Effects of SL65.0155, a Serotonin 5-HT4 Receptor Partial Agonist, in Animal Models of Amnesia,” Brain Research, Vol. 1121, No. 1, November 2006, pp. 207-215. doi:10.1016/j.brainres.2006.08.108
- C. Hille, S. Bate, J. Davis and M. I. Gonzalez, “5-HT4 Receptor Agonism in the Five-Choice Serial Reaction Time Task,” Behavioural Brain Research, Vol. 195, No. 1, December 2008, pp. 180-186. doi:10.1016/j.bbr.2008.08.007
- M. Matsumoto, H. Togashi, K. Mori, K.-I. Ueno, S. Ohashi, T. Kojima and M. Yoshioka, “Evidence for Involvement of Central 5-HT4 Receptors in Cholinergic Function Associated with Cognitive Processes: Behavioral, Electrophysiological, and Neurochemical Studies,” Journal of Pharmacology and Experimental Therapeutics, Vol. 296, No. 3, March 2001, pp. 676-682.
- E. G. Mohler, S. Shacham, S. Noiman, F. Lezoualc’h, S. Robert, M. Gastineau, J. Rutkowski, Y. Marantz, A. Dumuis, J. Bockaert, P. E. Gold and M. E. Ragozzino, “VRX-03011, a Novel 5-HT4 Agonist, Enhances Memory and Hippocampal Acetylcholine Efflux,” Neuropharmacology, Vol. 53, No. 4, September 2007, pp. 563-573. doi:10.1016/j.neuropharm.2007.06.016
- F. Shen, J. A. M. Smith, R. Chang, D. L. Bourdet, P. R. Tsuruda, G. P. Obedencio and D. T. Beattie, “5-HT4 Receptor Agonist Mediated Enhancement of Cognitive Function in Vivo and Amyloid Precursor Protein Processing in vitro: A Pharmacodynamic and Pharmacokinetic Assessment,” Neuropharmacology.
- A. Kulla and D. Manahan-Vaughan, “Modulation by Serotonin 5-HT4 Receptors of Long-Term Potentiation and Depotentiation in the Dentate Gyrus of Freely Moving Rats,” Cerebral Cortex, Vol. 12, No. 2, February 2002, pp. 150-162. doi:10.1093/cercor/12.2.150
- S. A. Small and K. Duff, “Linking Ab and Tau in Late- -Onset Alzheimer’s Disease: A Dual Pathway Hypothesis,” Neuron, Vol. 60, No. 4, 26 November 2008, pp. 534- 542. doi:10.1016/j.neuron.2008.11.007
- E. R. L. C. Vardy , A. J. Catto and N. M. Hooper, “Proteolytic Mechanisms in Amyloid-b Metabolism: Therapeutic Implications for Alzheimer’s Disease,” Trends in Molecular Medicine, Vol. 11, No. 10, October 2005, pp. 464-472. doi:10.1016/j.molmed.2005.08.004
- J. Nunan and D. H. Small, “Regulation of APP Cleavage by a-, band g-Secretases,” FEBS Letters, Vol. 483, No. 1, October 2000, pp. 6-10. doi:10.1016/S0014-5793(00)02076-7
- W.-L. Wu and L. Zhang, “g-Secretase Inhibitors for the Treatment of Alzheimer’s Disease,” Drug Development Research, Vol. 70, No. 2, March 2009, pp. 94-100. doi:10.1002/ddr.20288
- S. J. Stachel, “Progress toward the Development of a Viable BACE-1 Inhibitor,” Drug Development Research, Vol. 70, No. 2, March 2009, pp. 101-110. doi:10.1002/ddr.20289
- M. L. Hemming, J. E. Elias, S. P. Gygi and D. J. Selkoe, “Proteomic Profiling of g-Secretase Substrates and Mapping of Substrate Requirements,” PLoS Biology, Vol. 6, No. 10, October 2008, pp. 2314-2328. doi:10.1371/journal.pbio.0060257
- S. J. Robert, J. L. Zugaza, R. Fischmeister, A. M. Gardiers and F. Lezoualc’h, “The Human Serotonin 5-HT4 Receptor Regulates Secretion of Non-Amyloidogenic Precursor Protein,” Journal of Biological Chemistry, Vol. 276, No. 48, November 2001, pp. 44881-44888. doi:10.1074/jbc.M109008200
- M. Cachard-Chastel, F. Lezoualc’h, I. Dewachter, C. Delomenie, S. Croes, S. Devijver, M. Langlois, F. V. Leuven, S. Sicsic and A. M. Gardier, “5-HT4 Receptor Agonists Increase sAPPa Levels in the Cortex and Hippocampus of Male C57BL/6j Mice,” British Journal of Pharmacology, Vol. 150, No. 7, April 2007, pp. 883-892. doi:10.1038/sj.bjp.0707178
- S. Cho and Y. Hu, “Activation of 5-HT4 Receptors Inhibits Secretion of b-Amyloid Peptides and Increases Neuronal Survival,” Experimental Neurology, Vol. 203, No. 1, January 2007, pp. 274-278.
- S. Robert, M. Maillet, E. Morel, J.-M. Launay, R. Fischmeister, L. Mercken and F. Lezoualc’h, “Regulation of the Amyloid Precursor Protein Ectodomain Shedding by the 5-HT4 Receptor and Epac,” FEBS Letters, Vol. 579, No. 5, 14 February 2005, pp. 1136-1142. doi:10.1016/j.febslet.2005.01.010
- R. M. Eglen, D. W. Bonhaus, L. G. Johnson, E. Leung and R. D. Clark, “Pharmacological Characterization of Two Novel and Potent 5-HT4 Receptor Agonists, RS 67333 and RS 67506, in Vitro and in Vivo,” British Journal of Pharmacology, Vol. 115, 1995, pp. 1387-1392.
- M.-T. Liu, Y.-H. Kuan, J. Wang, R. Hen and M. D. Gershon, “5-HT4 Receptor Mediated Neuroprotection and Neurogenesis in the Enteric Nervous System of Adult Mice,” Journal of Neuroscience, Vol. 29, No. 31, August 2009, pp. 9683-9699. doi:10.1523/JNEUROSCI.1145-09.2009
- G. Lucas, V. V. Rymar, J. Du, O. Mnie-Filali, C. Bisgaard, S. Manta, L. Lambas-Senas, O. Wiborg, N. Haddjeri, G. Pineyro, A. F. Sadikot and G. Debonnel, “Serotonin4 (5-HT4) Receptor Agonists are Putative Antidepressants with a Rapid Onset of Action,” Neuron, Vol. 55, No. 5, September 2007, pp. 712-725.