Advances in Microbiology
Vol.3 No.1(2013), Article ID:29119,10 pages DOI:10.4236/aim.2013.31005
Purification and Characterization of Alkaline Xylanase Secreted from Paenibacillus macquariensis
School of Biotechnology, Devi Ahilya University, Indore, India
Email: *ak_sbt@yahoo.com
Received November 12, 2012; revised December 15, 2012; accepted January 16, 2013
Keywords: Xylanase; Paenibacillus macquariensis; Purification; Michaelis Constant; Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis
ABSTRACT
An alkaline xylanase secreted by Paenibacillus macquariensis RC 1819 has been purified using ammonium sulfate fractionation, ion exchange chromatography using DEAE-cellulose and gel filtration chromatography over Sephadex G-200 and Sephadex G-100. The purified enzyme had the specific activity, 25.2 units/mg protein with birchwood xylan as a substrate. The purified enzyme showed a single protein band over sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The molecular weight of the enzyme has been found to be 31,000 ± 2000 as determined by using Sephadex G-200 gel filtration chromatography. The subunit molecular weight has also been found to be ~31,000 as determined using SDS-PAGE indicating monomeric enzyme. The enzyme showed optimum activity at pH 8.6 and temperature, 50˚C. The Michaelis constant (Km) of the enzyme for birchwood xylan was 2.2 mg/ml as determined using velocity saturation plot. The metal ions viz. Co+2 and Mn+2 stimulated xylanase enzyme activity whereas Hg+2 inhibited the enzyme activity.
1. Introduction
Hemicellulose is the second most abundant component in plant cell wall and xylan is the major component of hemicellulose which is found in solid agricultural and agroindustrial residues, as well as in effluents released during wood processing [1,2]. Xylan is a heterogeneous carbohydrate consisting of a backbone of β-1, 4 linked D-xylopyranosyl units and short chain branches consisting of O-acetyl, α-L-arabinofuranosyl and α-D-glucuronyl residues [3]. A 20% - 35% of the total dry weight of plants accounts for xylans which can be used for production of fermentable sugars and fuels [4,5].
Xylanase is produced by many bacteria and fungi [6,7]. It has been exploited for a range of industrial and environmental applications. Xylanase is commercially important enzyme used in the pulp and paper industries to increase the brightness of pulp without the use of bleach [8,9]. Besides, xylanase is also used in number of other industries viz. food, beverage, textile and animal feed industries [7]. A recent application of xylanase is in the production of biofuels. It is estimated that the total energy content of global xylan and cellulose waste is equivalent to almost 640 billion tons of oil [10].
Xylanases are also used to convert the polymeric xylan into fermentable sugars for the production of ethanol and xylitol from plant biomass [11,12]. Xylanases can also be used for tailor designing of drugs and modifying the properties of food. Xylanases have also been used in animal feed to improve the digestibility of animal feed for better feed utilization [13,14].
Earlier, we showed that Paenibacillus macquariensis secretes extra-cellular xylanase and gets induced in the presence of xylan. We also optimized the conditions for its secretion. The Paenibacillus macquariensis is a gram positive bacteria. It showed optimum growth at 37˚C. However, the growth is much poor at 42˚C. The bacteria showed good growth in the pH range 5.2 to 10 indicating that this bacteria has much tolerance of pH change and has growth in acidic to highly alkaline range. This bacteria secreted xylanase enzyme as tested by Congo red dye staining method [15]. There are other reports also indicating secretion of xylanase by Paenibacillus species [16,17]. Therefore, the bacteria is capable of degrading xylan, xylanase being a xylan degrading enzyme. In the present study, we reported its purification and characterization from Paenibacillus macquariensis RC 1819. The main objective of the present work was to purify xylanase secreted by Paenibacillus macquariensis and to compare the characteristics of the present xylanase enzyme with the enzymes earlier reported in order to check its suitability for industrial exploitation.
2. Material and Methods
2.1. Culture and Growth Conditions
A soil bacterium Paenibacillus macquariensis was isolated in pure form from petroleum containing soil collected from the vicinity of a petrol pump at Indore as described earlier [15]. Pure culture of P. macquariensis was used as master culture for subsequent production of xylanase. Pure bacterial culture was inoculated into sterilized Emmerson medium (yeast extract, 5 gm; peptone, 5 gm; K2HPO4, 1 gm; MgSO4·7H2O, 0.2 gm per liter and pH adjusted to 9 with 1N NaOH). The medium was supplemented with 2% birchwood xylan (purchased from Sigma-Aldrich, USA) in order to induce xylanase secretion. The culture was grown at 37˚C for 48 hours.
2.2. Enzyme Assay
Xylanase enzyme was assayed by measuring the release of the reducing sugars from birchwood xylan following the dinitrosalicylic acid (DNS) method [18]. A 0.9 ml sample of 1% birchwood xylan dissolved in 50 mM glycine-NaOH buffer, pH 8.6 was pre-incubated at 50˚C for 5 minutes. To this, 0.1 ml enzyme (supernatant of the broth/purified enzyme) and buffer was added (buffer was added with purified enzyme if amount of the enzyme preparation taken was less than 0.1 ml) and incubated at 50˚C for 15 minutes. The reaction was stopped by adding 1.5 ml of 1% (w/v) DNS solution and the tubes were put in a boiling water bath for 15 minutes. A control was also run simultaneously where enzyme was added after the addition of DNS. A blank was also prepared where no enzyme was added and against the blank, zero was set in the colorimeter. The D-xylose was used as standard during the colorimetric estimation. One unit of xylanase activity was taken as the amount of the enzyme required to release one micromole of the reducing sugar equivalent to one micromole of xylose per minute at 50˚C under conditions of the enzyme assay.
2.3. Protein Estimation
Protein estimation was carried out according to the procedure of Lowry et al. [19]. In the enzyme sample, protein was precipitated out by adding trichloroacetic acid (TCA) to a final concentration of 5% and thereafter, sample was incubated for 4 to 5 hours in the cold condition (0˚C to 4˚C). TCA precipitates proteins as protein trichloroacetate. Thereafter, the precipitate was collected by centrifugation and washed with 5% TCA to remove any adhering impurity(ies). The washed precipitate was dissolved in 0.1N sodium hydroxide. This dissolved sample was used as protein sample for estimation using Folin Ciocalteau reagent as described by Lowry et al. [19]. Bovine serum albumin was used as a standard protein.
2.4. Enzyme Purification
2.4.1. Crude Enzyme Preparation
The Paenibacillus macquariensis culture (500 ml) in Emmerson medium with 2% xylan after growth for 48 hours at 37˚C was centrifuged at 10,000 ×g for 10 minutes at 0˚C to 4˚C in a cooling centrifuge. The supernatant was used as crude enzyme preparation. Thereafter, following procedure was carried out at 0˚C - 4˚C.
2.4.2. Ammonium Sulfate Fractionation
To the crude enzyme preparation (500 ml), finely ground solid ammonium sulfate was added slowly with constant stirring maintaining the pH at 9.0 by the addition of 1% (v/v) ammonia to get 0% - 30% saturation. After storage for 3 hours in the cold condition, the suspension was centrifuged at 10,000 ×g for 30 minutes and the supernatant was further subjected to 30% - 60% saturation by adding finely ground ammonium sulfate. After storage for 3 hours in the cold condition, the suspension was centrifuged at 10,000 ×g for 30 minutes. The pellet (30% to 60% pellet) was dissolved in 50 mM potassium phosphate buffer, pH 8.6 (buffer A). It was a turbid suspension, therefore, it was centrifuged at 10,000 ×g for 10 minutes to get clear supernatant which was desalted by passing through a column of Sephadex G-25.
2.4.3. DEAE Cellulose Chromatography
The desalted ammonium sulfate fraction was loaded onto a DEAE-cellulose column (2.5 × 40 cm), previously equilibrated with buffer A. After washing with buffer A, enzyme was eluted by using a linear NaCl gradient (0 to 1 M) and volume of the gradient was 1000 ml (500 ml buffer A in each mixing chamber and reservoir). Fractions of 10 ml were collected at a flow rate of 2 ml/min. The fractions having xylanase enzyme activity constituting a single peak were pooled. To this, solid ammonium sulfate was added to get 90% saturation and the suspension was kept for 5 hours for complete precipitation. The precipitate was collected by centrifugation at 10,000 ×g for 10 minutes and dissolved in buffer A. The turbid suspension was centrifuged at 10,000 ×g for 10 minutes to get clear supernatant
2.4.4. Sephadex G-200 Chromatography
A Sephadex G-200 column (1.5 × 60 cm) was equilibrated with buffer A. The concentrated xylanase fraction after DEAE-cellulose chromatography was loaded onto the column and subsequently chromatographed using buffer A. Fractions of 2 ml were collected at a flow rate of 10 ml/hour and the active fractions pooled. To the pooled fraction, solid ammonium sulfate was added to get 90% saturation and the suspension was kept for 5 hours for complete precipitation. The precipitate was collected by centrifugation at 10,000 ×g for 10 minutes and dissolved in buffer A. The turbid suspension was centrifuged at 10,000 ×g for 10 minutes to get clear supernatant.
2.4.5. Sephadex G-100 Chromatography
A Sephadex G-100 column (1.5 × 50 cm) was equilibrated with buffer A. The concentrated xylanase fraction after Sephadex G-200 chromatography was loaded onto the column and subsequently chromatographed using buffer A. Fractions of 2 ml were collected at a flow rate of 12 ml/hour and the active fractions pooled.
2.4.6. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was carried out according to the procedure of Laemmli [20] using 10% acrylamide in slab gels (1 mm thick). The protein (xylanase during various purification steps and protein markers) was incubated for 5 minutes at 100˚C in the presence of 1% sodium dodecyl sulfate and 0.02 M 2-mercaptoethanol to convert it into monomers. Around 20 µg of enzyme protein was loaded in the well. Bromophenol blue was used as a tracking dye. The buffer systems used were 0.2 M Tris-HCl, pH 8.8 for separating gel and 0.2 M Tris-HCl pH 6.8 for the stacking gel. Electrophoresis was carried out using a current of 3 mA/cm. The current was turned off when the tracking dye reached the bottom of the gel. Coomassie brilliant blue (0.2%) in methanol, acetic acid and water (3:1:6) was used to stain the protein bands. The gel was stained for one hour and then repeatedly destained using the same solvents used for dissolving coomassie brilliant blue till the gel turned transparent and distinct bluish bands were visible.
2.5. Molecular Weight Determination
Native molecular weight of xylanase was determined using gel filtration chromatography over Sephadex G- 200 according to the procedure of Whitaker [21]. The purified enzyme after Sephadex G-100 chromatography was applied on to a Sephadex G-200 column (1.5 × 60 cm), pre-equilibrated with buffer A. The flow rate was 12 ml/hour. Bovine serum albumin (MW 66,000), trypsinogen (MW 23,000) and lysozyme (MW 14,000) were used as standard proteins (all from Sigma-Aldrich, USA). Subunit molecular weight was determined by sodium dodecyl sulfate polyacrylamide gel electrophoresis as described above. Standard molecular markers having subunit molecular weights of 97,000, 66,000, 45,000, 29,000, 21,000, and 14,000 (all from Helini Biomolecules, India) were used as standard proteins. To denature the proteins, solutions were incubated with 1% SDS and 20 mM 2- mercaptoethanol at 100˚C for 5 minutes.
2.6. Effect of pH on Xylanase Activity
The relative xylanase activity at different pHs was determined using 1% birchwood xylan as substrate. The pH range used was from 4 to 11. Three different buffers were used viz. 0.1 M citrate buffer for pH range 4 to 6, 0.1 M sodium phosphate buffer for pH range 6 to 8, and 0.1 M glycine NaOH buffer for pH range 8 to 11. The substrate (1% birchwood xylan) was prepared in buffers of different pHs and pre-incubated at 50˚C for 10 minutes. To this, 0.1 ml enzyme was added and incubated for 15 minutes. The reaction was stopped by adding 1.5 ml of DNS solution and the tubes were put in a boiling water bath for 15 minutes. The further procedure was same as given under the enzyme assay.
2.7. pH-Stability of the Enzyme
For pH stability, xylanase enzyme was incubated with different buffers viz. 0.1 M citrate buffer for pH range 4 to 6, 0.1 M sodium phosphate buffer for pH range 6 to 8, and 0.1 M glycine NaOH buffer for pH range 8 to 11 at room temperature (25˚C) for 30 minutes. Thereafter, enzyme activity was determined at pH 8.6 by using the enzyme assay as described above.
2.8. Effect of Temperature on Xylanase Activity
The apparent optimum temperature for the purified xylanase was determined by assaying the enzyme activity at different temperatures ranging from 30˚C to 70˚C. At each temperature, 0.9 ml of 1% birchwood xylan dissolved in 50 mM glycine-NaOH buffer, pH 8.6 was preincubated for 10 minutes. To this, 0.1 ml enzyme was added and incubated for 15 minutes. The reaction was stopped by adding 1.5 ml of DNS solution and the tubes were put in a boiling water bath for 15 minutes. The further procedure was same as given under the enzyme assay.
2.9. Thermo-Stability of the Enzyme
For thermo-stability, xylanase enzyme was pre-incubated at different temperatures ranging from 30˚C to 100˚C in a water bath. After every 30 minutes, the sample tubes were removed from the water bath and stored on ice prior to enzyme assay. Thereafter, enzyme activity was determined at 50˚C as described above.
2.10. Substrate Specificity
Xylanase activity was determined using cellulose, birchwood xylan, xylobiose as substrates.
2.11. Xylanase-Metal Ions Binding Studies
Xylanase aliquots (0.5 ml) were incubated at the room temperature (25˚C) with 1 mM metal ions viz. Ca+2, Mg+2, Hg+2, Fe+2, Cu+2, Mn+2, Co+2, Zn+2, As+3, Mo+2 for 2 hours. Thereafter, enzyme activity was determined using 0.1 ml of the incubated enzyme with 0.9 ml of 1% birchwood xylan dissolved in 50 mM glycine-NaOH buffer, pH 8.6 and incubated at 50˚C for 15 min.
2.12. Km Determination
Birchwood xylan hydrolysis rates were determined at all substrate concentrations ranging from 0.5% to 8% in 50 mM glycine-NaOH buffer at pH 8.6.
3. Results
3.1. Enzyme Purification
The summary of purification of xylanase from Paenibacillus macquariensis has been shown in Table 1. The ion exchange chromatography over DEAE-cellulose of the 30% to 60% ammonium sulfate fraction revealed only one peak of xylanase activity that was eluted at 0.22 M NaCl (Figure 1). The concentration of NaCl in the eluent was calculated by using the formula given by Morris and Morris [22]. The gel filtration chromatogramphy over Sephadex G-200 of the enzyme fraction after ion exchange chromatography also revealed a single peak of xylanase activity (Figure 2). Since after Sephadex G- 200 chromatography, enzyme preparation showed a minor band also on sodium dodecyl sulfate polyacrylamide gel electrophoresis, it was further loaded on to a Sephadex G-100 column and chromatographed using buffer A. After Sephadex G-100 chromatography, there was single peak of enzyme activity. The results indicated absence of multiple forms of the enzyme. The Sephadex G-200 chromatography resulted in 11.62 fold purification of xylanase with 25.3% recovery from the crude extract. Thereafter, Sephadex G-100 chromatography resulted in 14.24 fold purification of the enzyme with 22.3% recovery. The specific activity of the enzyme was found to be 25.20 units/mg protein. The purified enzyme showed a single protein band on sodium dodecyl sulfate polyacrylamide gel electrophoresis (Figure 3).
Table 1. Purification profile of xylanase from Paenibacillus macquariensis. The final purified xylanase showed specific activity, 25.20 units/mg protein.
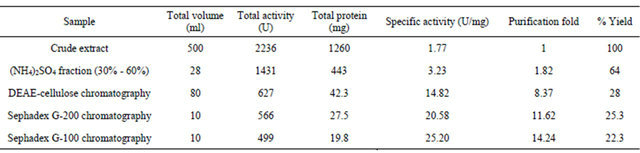
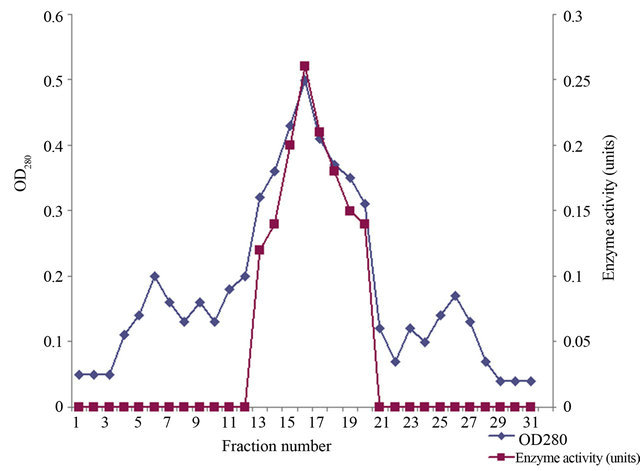
Figure 1. Elution profile of xylanase on DEAE cellulose chromatography. The profile showed single peak of xylanase enzyme activity indicating absence of multiple forms.
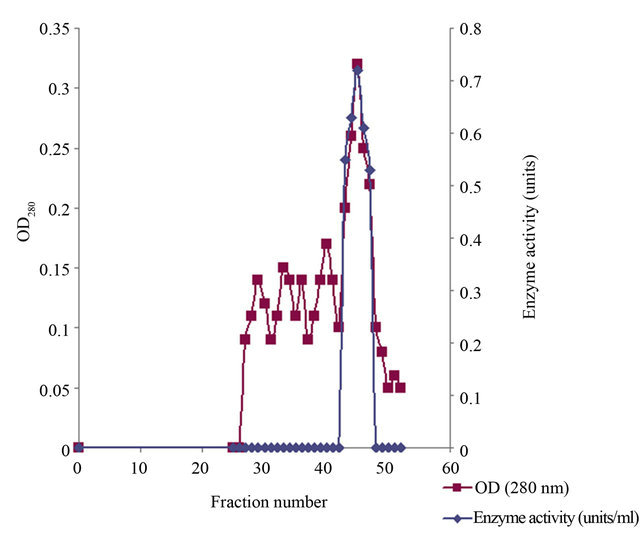
Figure 2. Elution profile of xylanase on Sephadex G-200 chromatography. The profile showed single peak of xylanase enzyme activity indicating absence of multiple forms.
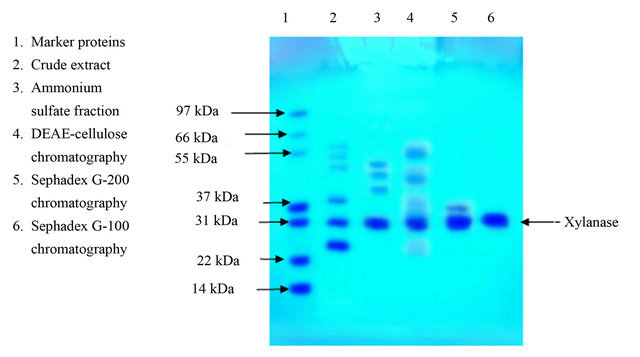
Figure 3. SDS-PAGE analysis during purification steps of xylanases from Paenibacillus macquariensis. The purified xylanase after Sephadex G-100 chromatography showed a single band.
3.2. Molecular Weight
The molecular weight of xylanase was found to be 31,000 ± 2000 as determined by using Sephadex G-200 gel filtration chromatography according to the procedure of Whitaker [21]. Bovine serum albumin, trypsinogen and lysozyme were used as standard markers (Figure 4). On sodium dodecyl sulfate polyacrylamide gel electrophoresis, the subunit molecular weight of single band obtained was ~31,000 indicating monomeric nature of the enzyme.
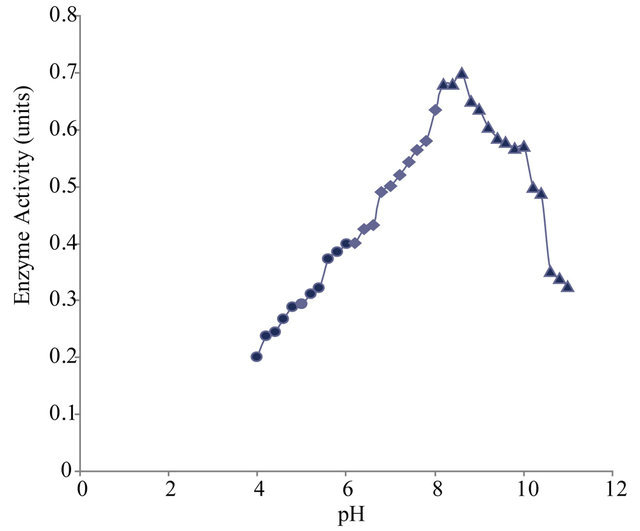
3.3. Effect of pH on Xylanase Activity
Enzyme assay was carried out in the pH range varying from pH 4 to 11 to find out the optimum pH at which the enzyme shows maximum activity. The purified xylanase showed maximum activity (optimum pH) at pH 8.6 and half maximum activity at pH 5.8 and pH 10.6 (Figure 5(a)).
3.4. pH Stability of the Enzyme
The pH stability profile showed that xylanase is more stable in the pH range 8 to 9 (Figure 5(b)).
3.5. Effect of Temperature on Xylanase Activity
Enzyme assay was carried out in the temperature range of 30˚C to 70˚C to find out the apparent optimum temperature at which the enzyme shows maximum activity. The purified xylanase showed maximum activity (optimum temperature) at 50˚C and half maximum activity at 27˚C and 67˚C (Figure 5(c)).
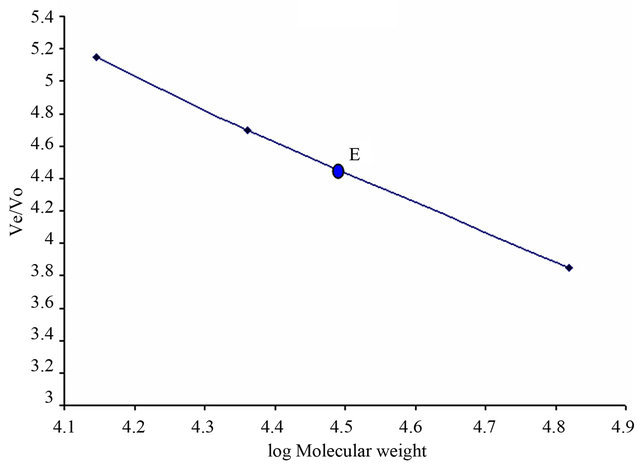
Figure 4. Molecular weight determination of xylanase using Sephadex G-200 gel filtration chromatography (Whitaker method [21]). 1) Bovine serum albumin (MW 66,000); 2) trypsinogen (MW 23,000); 3) lysozyme (MW 14,000). The molecular weight of xylanase has been estimated to be 31,000 ± 2000.
 (a)
(a)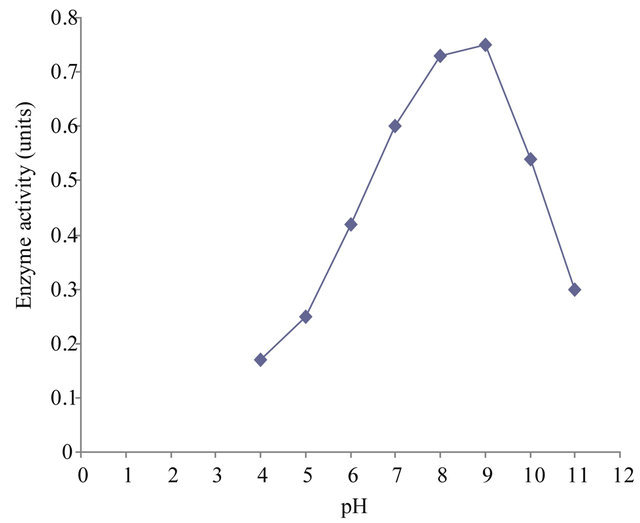 (b)
(b)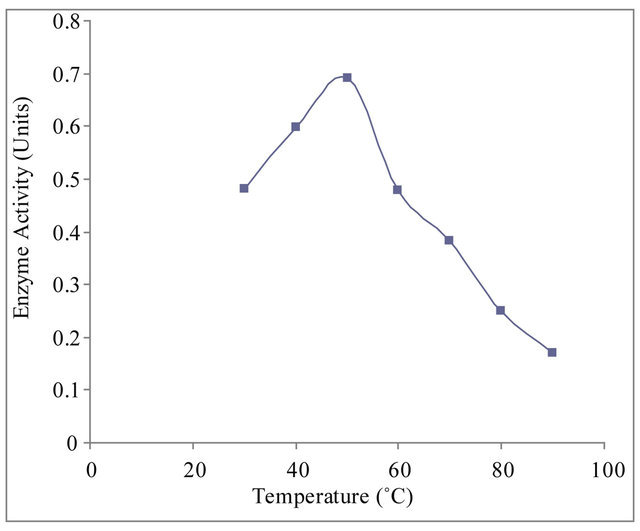 (c)
(c)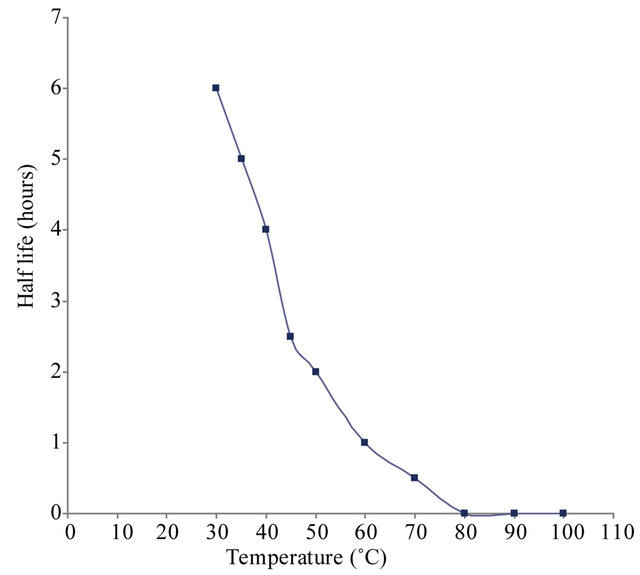 (d)
(d)
Figure 5. (a) Effect of pH on xylanase activity of Paenibacillus macquariensis. The purified xylanase showed maximum activity (optimum pH) at pH 8.6 and half maximum activity at pH 5.8 and pH 10.6. (b) pH-stability profile of xylanase. It showed more stability in the pH range 8 to 9. (c) Effect of temperature on xylanase activity. The enzyme showed maximum activity (optimum temperature) at 50˚C and half maximum activity at 27˚C and 67˚C. (d) Temperature stability profile of xylanase. At 50˚C, the enzyme showed a half life of two hours whereas at 60˚C, it showed a half life of one hour only. In cases of (a), (b) and (c), 15 μl of purified xylanase was used in the enzyme assay.
3.6. Thermo-Stability of the Enzyme
The thermo-stability studies showed that xylanase is stable up to 60˚C. At 50˚C, enzyme had a half life of two hours whereas at 60˚C, it showed a half life of one hour only (Figure 5(d)).
3.7. Substrate Specificity
The enzyme hydrolyzed β-1,4-xylan (birchwood xylan). It also hydrolyzed cellulose (CM-cellulose) but activity was nearly 10% as compared to xylan. The enzyme showed no activity with xylobiose.
3.8. Effect of Metal Ion Binding on Xylanase Activity
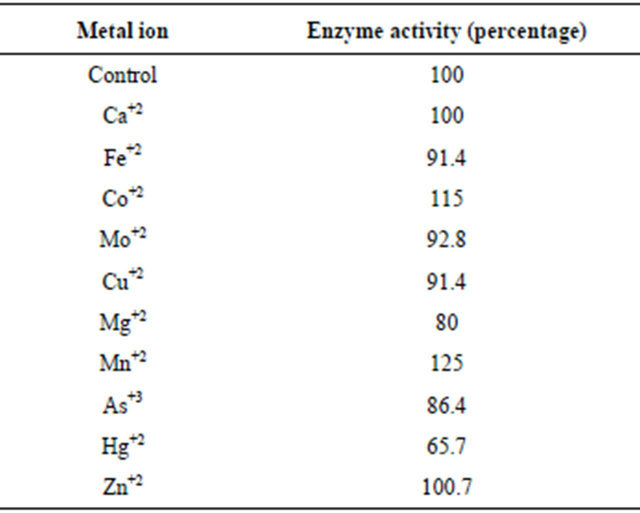
Purified xylanase aliquots were incubated at the room temperature (25˚C) with 1 mM metal ions viz. Ca+2, Mg+2, Hg+2, Fe+2, Cu+2, Mn+2, Co+2, Zn+2, As+3, Mo+2 for 2 hours. Enzyme activity got stimulated upon incubation with Co+2 and Mn+2. However, metal ions viz. Ca+2, Fe+2, Mo+2, Cu+2, Mg+2, Zn+2, As+3 showed no considerable effect on xylanase activity. In contrast, 1 mM Hg+2 showed 35% inhibition of xylanase activity. The purified xylanase was also incubated without any metal ion which was taken as a control (Table 2). Inhibition of the enzyme by Hg+2 looks due to heavy metal effect since if it is due to –SH group(s), inhibition would be nearly 100% at 1 mM concentration of Hg+2. In a separate experiment, enzyme was incubated with EDTA in place of a metal ion. EDTA showed no effect on the enzyme activity.
3.9. Km (Michaelis Constant) Determination
Xylan saturation curve was found to be rectangular hyperbolic indicating that the enzyme obeyed classical
Table 2. Effect of various metal ions on xylanase activity. Enzyme activity got stimulated upon incubation with Co+2 and Mn+2. However, Hg+2 inhibited the enzyme activity.
Michaelis kinetics. The Km value of xylanase for birchwood xylan was calculated to be 2.2 ± 0.2 mg/ml.
4. Discussion
Microorganisms of extreme environments have attracted much attention owing to their habitat-related adaptive properties. For example, alkaline xylanases have been isolated from microbes and plants of alkaline or neutral environments.
To obtain an alkaline xylanase we selected the soil containing petroleum as the source material for microorganism isolation. The enzyme production was maximum on birchwood xylan. Alkaline xylanases are of much industrial importance especially in paper and pulp Industries.
Earlier, from our laboratory, Mahatman et al. [23] purified xylanase from Bacillus halodurans strain KR-1 and showed it to be nearly 70% pure with a specific activity of 14.61 units/mg protein. Vahino and Nakane [24] purified xylanase from a Bacillus sp. and showed specific activity of the purified enzyme to be 5.33 units/mg protein. Blanco et al. [25] purified xylanase from an another Bacillus sp. and reported its specific activity to be 40.2 units/mg protein. Nakamura et al. [26] reported specific activity of purified xylanase from a Bacillus sp. to be 310 units/mg protein. However, Gessesse [27] reported presence of two multiple forms of xylanase in a Bacillus species having specific activity of 524.2 and 367.9 units/mg protein, respectively. From the data, it is evident that there is wide variation in the specific activity of xylanase from different bacterial species. In the present case, specific activity is at par or more compared to many other reports. On the other hand, if compared with the xylanase purified from Bacillus sp. studied by Nakamura et al. [26] and Gessesse [27], specific activity in the present case is comparatively low. Recently, Dheeran et al. [16] reported a highly active xylanase from Paenibacillus macerans IIPSP3 having enzyme activity 4170 units/mg protein. However, it is more active at acidic pH 4.5.
Xylanase from Paenibacillus macquariensis showed maximum activity at pH 8.6. However, it exhibited enzyme activity over a broad pH range of pH 4 - 11, Earlier, from our laboratory, Mahatman et al. [23] reported optimum pH 9.0 for xylanase from Bacillus halodurans strain KR-1. Nakamura et al. [26] reported optimum pH 9 for xylanase from a Bacillus sp. strain 41M-1. However, Blanco et al. [25] reported optimum pH 5.5 of xylanase from a Bacillus species strain BP-23. Vahino and Nakane [24] showed optimum pH 4 for the enzyme from a Bacillus sp. Kubata et al. [28] reported optimum pH 6.8 for xylanase from Aeromonas caviae ME-1. An optimum pH 7.5 has been reported for xylanase from Alcaligenes sp. XY-234 [29]. Lin et al. [30] reported optimum pH 6.5 for xylanase from Thermomyces lanuginosus-SSBP.
The present enzyme may be exploited as chicken feed additive since physiological pH of chicken digestive tract has been reported to be ranging from pH 4.5 to 7.5 [31]. The results are comparable with an antarctic bacterium, Pseudoalteromonas haloplanktis, whose xylanase has been reported to exhibit enzyme activity in a wide pH range between pH 5.3 and 8 [32]. Xylanase from Paenibacillus curdlanolyticus B-6 derived from an anaerobic fermenter has also been shown to retain more than 80% activity in a wider pH range of pH 5.0 to 9.0 [33].
Xylanase from Paenibacillus macquariensis showed maximum (optimum) activity at 50˚C. The thermo-stability of the enzyme was also tested after pre-incubation of the enzyme at 30˚C to 100˚C for 30 minutes. The enzyme had half life of two hours at 50˚C whereas it had a half life of one hour at 60˚C. At high temperatures, enzyme gets partly unfolded. Indeed, this point of maximum activity is the result of the intersection of an exponential curve corresponding to the increase of the activity as a function of temperature and of an unfolding curve due to deleterious effect of heat. Dheeran et al. [16] reported a xylanase from Paenibacillus macerans exhibiting activity over a broad range of temperature, 40˚C to 90˚C with optimum activity at 60˚C. However, Heck et al. [34] showed that xylanase isolated from Bacillus coagulans BL69 and grown on soybean residue exhibited enzyme activity over a wide range of temperature ranging from 45˚C to 75˚C, However, thermal stability of the enzyme was much poor. Wamalwa et al. [35] also reported optimum temperature 40˚C for xylanase from an another strain of Bacillus halodurans. However, Nakamura et al. [36] reported optimum temperature 50˚C for xylanase from Bacillus sp. strain 41M-1. Blanco et al. [25] also reported optimum temperature 50˚C for the enzyme from a Bacillus sp. strain BP-23. However, Vahino and Nakane [24] showed optimum temperature 80˚C for xylanase from a Bacillus sp. Kubata et al. [28] showed optimum temperature of xylanases from Aeromonas caviae ME-1 to be 30˚C and 37˚C. Araki et al. [29] reported optimum temperature, 40˚C for xylanase from Alcaligenes sp. XY-234. On the other hand, an optimum temperature of 70˚C - 75˚C has been reported for xylanase from Thermomyces lanuginosus-SSBP [30]. Therefore, it is clear that there is wide variation in the optimum temperature of xylanase from different microbes.
In the present case, xylanase activity was stimulated in the presence of Co+2 and Mn+2 ions. However, xylanase activity got inhibited in the presence of Hg+2 ions. There are reports of similar results [37-39]. Inkyung and Jaiesoon [40] reported stimulation of Paenibacillus sp. strain K1J1 xylanase in the presence of Ca2+, Co2+, Zn2+, Cu 2+, and Mn2+. Lee et al. [41] reported stimulation of Bacillus licheniformis xylanase in the presence of Ca2+, Co2+, Zn2+, Cu2+, and Mn2+. However, Collins et al. [1] reported inhibition of xylanase from the antarctic bacterium, Pseudoalteromonas haloplanktis in the presence of Cu2+ and Zn2+.
In the present case, Km value of the enzyme for birchwood xylan has been calculated to be 2.2 mg/ml. The Km value of the enzyme from Arthrobacter for wheat bran has been reported to be 0.9 mg/ml. Similarly, Km value of the enzyme from Bacillus subtilis and Bacillus sp. strain TAR-1 has been reported in the range of 0.9 mg/ml [42,36]. Therefore, in the present case, Km value is more compared to other reported values indicating lower affinity for the substrate. However, here we used birchwood xylan and in the other reports, it was wheat bran.
The molecular weight of the enzyme in the present case has been found ~31,000. Ratanakhanokchai and Tanticharoen [43] reported a molecular weight of 23,000 for xylanase from Bacillus sp. strain K-1, whereas, Muniswaran et al. [44] reported a molecular weight 36,000 for xylanase from Bacillus sp. 41M. Contrarily, Sa-Pereira et al. [42] reported a molecular weight of 340,000 for xylanase from Bacillus subtilis. Dheeran et al. [16] reported a molecular weight, 205,000 for xylanase from Paenibacillus macerans.
5. Acknowledgements
The authors acknowledge the Department of Biotechnology, Ministry of Science and Technology, Government of India, New Delhi, India for its facilities under M.Sc. Biotechnology program and the Bioinformatics Sub Centre.
REFERENCES
- T. Collins, C. Gerday and G. Feller, “Xylanases, Xylanase Families and Extremophilic Xylanases,” FEMS Microbiology Reviews, Vol. 29, No. 1, 2005, pp. 3-23. doi:10.1016/j.femsre.2004.06.005
- L. Anthony, A. Marcel and R. Eric, “Overproduction and Characterization of Xylanase B from Aspergillus niger,” Canadian Journal of Microbiology, Vol. 51, No. 2, 2005, pp. 177-183. doi:10.1139/w04-116
- S. G. Nair, R. Sindhu and S. Shashidhar, “Fungal Xylanase Production under Solid State and Submerged Fermentation Conditions,” African Journal of Microbiology Research, Vol. 2, No. 4, 2008, pp. 82-86.
- D. Haltrich, B. Nidetzky, K. D. Kulbe, W. Steiner and S. Zupancic, “Production of Fungal Xylanases,” Bioresource Technology, Vol. 58, No. 2, 1996, pp 137-161. doi:10.1016/S0960-8524(96)00094-6
- E. X. F. Filho, “Hemicellulase and Biotechnology,” In: S. G. Pandalai, Ed., Recent Research Development in Microbiology, Research Signpost, Trivandrum, 1998, pp. 165- 176.
- C. T. D. Marta, C. A. P. Ana, P. P. Edilberto, N. P. Alexandre and T. F. Telma, “Characterization of Alkaline Xylanases from Bacillus pumilus,” Brazilian Journal of Microbiology, Vol. 31, No. 2, 2000, pp. 90-94.
- M. L. T. M. Polizeli, A. C. S. Rizzatti, R. Monti, H. F. Terenzi, J. A. Jorge and D. S. Amorim, “Xylanases from Fungi: Properties and Industrial Applications,” Applied Microbiology and Biotechnology, Vol. 67, No. 5, 2005, pp. 577-591. doi:10.1007/s00253-005-1904-7
- L. Viikari, M. Ranua, A. Kantelinen, J. Sundquist and M. Linko, “Bleaching with Enzymes,” Proceedings of the 3rd International Conference on Biotechnology, Pulp Paper Industry, Stockholm, 1986, pp. 67-69.
- N. Garg, K. K. Mahatman and A. Kumar, “Xylanase: Applications and Biotechnological Aspects,” LAP Lambert Academic Publishing AG & Co. KG, 2010.
- P. S. Pereira, H. Paveia, M. C. Ferreira and M. R. AiresBarros, “A New Look at Xylanases: An Overview of Purification Strategies,” Molecular Biotechnology, Vol. 24, No. 3, 2003, pp. 257-281. doi:10.1385/MB:24:3:257
- M. Galbe and G. A. Zacchi, “A Review of the Production of Ethanol from Softwood,” Applied Microbiology and Biotechnology, Vol. 59, No. 6, 2005, pp. 618-628.
- Q. A. Beg, M. Kapoor, G. Mahajan and S. Hoondal, “Microbial Xylanases and Their Industrial Applications: A Review,” Applied Microbiology Biotechnology, Vol. 6, No. 3-4, 2001, pp. 326-338. doi:10.1007/s002530100704
- T. N. Nortey, J. F. Patience and R. T. Zijlstra, “Xylanase Supplementation Improves Energy Digestibility of Wheat By-Products in Grower Pigs,” Livestock Science, Vol. 109, No. 1, 2007, pp. 96-99. doi:10.1016/j.livsci.2007.01.092
- H. Uhlig, “Industrial Enzymes and Their Applications,” John Wiley & Sons Ltd., New York, 1998.
- M. Sharma and A. Kumar, “Optimization of Xylanase Secretion from Paenibacillus Macquariensis,” Current Trends in Biotechnology and Pharmacy, Vol. 6, No. 2, 2012, pp. 190-195.
- P. Dheeran, N. Nandhagopal, S. Kumar, Y. K. Jaiswal and D. K. Adhikari, “A Novel Thermostable Xylanase of Paenibacillus Macerans IIPSP3 Isolated from the Termite Gut,” Journal of Industrial Microbiology & Biotechnology, Vol. 39, No. 6, 2012, pp. 851-860. doi:10.1007/s10295-012-1093-1
- P. Shi, J. Tian, T. Yuan, X. Liu, H. Huang, Y. Bai, P. Yang, X. Chen, N. Wu and B. Yao, “Paenibacillus sp. Strain E18 Bifunctional Xylanase-Glucanase with a Single Catalytic Domain,” Applied and Environmental Microbiology, Vol. 76, No. 11, 2010, pp. 2620-2624. doi:10.1128/AEM.00345-10
- G. L. Miller, “Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar,” Analytical Chemistry, Vol. 31, No. 3, 1959, pp. 426-428. doi:10.1021/ac60147a030
- O. H. Lowry, N. J. Rosebrough, A. L. Farr and R. J. Randall, “Protein Measurement with the Folin Phenol Reagent,” Journal of Biological Chemistry, Vol. 193, No. 1, 1951, pp. 265-275.
- U. K. Laemmli, “Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4,” Nature, Vol. 227, No. 5259, 1970, pp. 680-685.
- J. R. Whitaker, “Determination of Molecular Weights of Proteins by Gel Filtration on Sephadex,” Analytical Chemistry, Vol. 35, No. 12, 1963, pp. 1950-1953.
- C. J. O. R. Morris and P. Morris, In: 2nd Edition, Separation Methods in Biochemistry, Pitman Publishing, London, 1976, pp. 93-148.
- K. K. Mahatman, N. Garg, R. Chauhan and A. Kumar, “Production, Purification and Characterization of Xylanase Using Alkalo-Thermophilic Bacillus halodurans KR-1,” Iranica Journal of Energy and Environment, Vol. 1, No. 4, 2010, pp. 265-274.
- F. Vahino and T. A. Nakane, “Thermostable Xylanase from a Thermophilic Acidophilic Bacillus sp.,” Agricultural and Biological Chemistry, Vol. 45, No. 5, 1980, pp. 1121-1127.
- A. Blanco, T. Vidal, F. J. Colom and J. I. F. Pastor, “Purification and Properties of Xylanase A from Alkali-Tolerent Bacillus sp. Strain BP-23,” Applied and Environmental Microbiology, Vol. 61, No. 12, 1995, pp. 4468- 4470.
- S. Nakamura, K. Wakabayashi, R. Nakai, R. Aono and K. Horikoshi, “Purification and Some Properties of an Alkaline Xylanase from an Alkaliphilic Bacillus sp. Strain 41M-1,” Applied and Environmental Microbiology, Vol. 59, No. 7, 1993, pp. 2311-2316.
- A. Gessesse, “Purification and Properties of Two Thermostable Alkaline Xylanase from an Alkaliphilic Bacillus sp.,” Applied and Environmental Microbiology, Vol. 64, No. 9, 1998, pp. 3533-3535.
- B. K. Kubata, T. Suzuki, H. Horitsu, K. Kawai and K. Takamizawa, “Purification and Characterization of Aeromonas carviae ME-1 Xylanase,Which Produces Exclusively Xylobiose from Xylan,” Applied and Envionmental Microbiology, Vol. 60, No. 2, 1994, pp. 531-535.
- T. Araki, N. Enoue and T. Morisita, “Purification and Characterization of β-1,3 Xylanase from a Marine Bacterium, Alcaligenes sp. XY-234,” The Journal of General and Applied Microbiology, Vol. 44, No. 4, 1998, pp. 269- 274.
- J. Lin, L. M. Nudlovu, S. Singh and B. Pillay, “Purification and Biochemical Charactrastics of β-D-Xylanase from a Thermophilic Fungus, Thermomyces lanuginosusSSBP,” Biotechnology and Applied Biochemistry, Vol. 30, No. 1, 1999, pp. 73-79.
- J. S. Cho, K. Choi, T. Darden, P. R. Reynolds, J. N. Petitte and S. B. Shears, “Avian Multiple Inositol Polyphosphate Phosphatase in an Active Phytase That Can Be Engineered to Help Ameliorate the Planet’S ‘Phosphate Crisis’,” Journal of Biotechnology, Vol. 126, No. 2, 2006, pp. 248-259.
- T. Collins, M. A. Meuwis, I. Stals, M. Claeyssens, G. Feller and C. Gerday, “A Novel Family 8 Xylanase: Functional and Physiochemical Characterization,” The Journal of Biological Chemistry, Vol. 277, No. 38, 2002, pp. 35133-35139.
- W. Rattiya, P. Pason, K. L. Kyu, K. Sakka, A. Kosugi, Y. Mori and K. Ratanakhanokchai, “Cloning, Sequencing, and Expression of the Gene Encoding a Multidomain endo-1, 4-Xylanase from paenibacillus curdlanolyticus b-6, and Characterization of the Recombinant Enzyme,” Journal of Microbiology and Biotechnology, Vol. 19, No. 3, 2009, pp. 277-285.
- J. Heck, S. Flores, P. Hertzm and M. Ayub, “Optimization of Cellulase-Free Xylanase Activity Produced by Bacillus coagulans BL69 in Solid-State Cultivation,” Process Biochemistry, Vol. 40, No. 1, 2005, pp. 107-112.
- M. B. Wamalwa, G. Zhao, M. Sakka, M. P. Shiundu, T. Kimura and K. Sakka, “High Level Heterologous Expression of Bacillus halodurans Putative Xylanase xyn11A (BHO899) in Kluyveromyces lactis,” Bioscience, Biotechnology and Biochemistry, Vol. 71, No. 3, 2007, pp. 688-693.
- S. Nakamura, Y. Ishiguro, R. Nakai, K. Wakabayashi, R. Aono and K. Horikoshi, “Purification and Characterization of a Thermophilic Alkaline Xylanase from Thermoalkalophilic Bacillus sp. strain TAR-1,” Journal of Molecular Catalysis B: Enzymatics, Vol. 1, No. 1, 1995, pp. 7-15.
- K. B. Bastawade, “Xylan Structure, Microbial Xylanases, and Their Mode of Action,” World Journal of Microbiology and Biotechnology, Vol. 8, No. 4, 1992, pp. 353- 368.
- T. Cesar and V. Mrsia, “Purification and Properties of the Xylanase Produced by Thermomyces lanuginosus,” Enzyme and Microbial Technology, Vol. 19, No. 4, 1996, pp. 289-296.
- E. T. Chivero, A. N. Mutukumira and R. Zvauya, “Partial Purification and Characterization of a Xylanase Enzyme Produced by a Micro-Organism Isolated from Selected from Indigenous Fruits of Zimbabwe,” Food Chemistry, Vol. 72, No. 2, 2001, pp. 179-185.
- P. Inkyung and C. Jaiesoon, “Partial Characterization of Extracellular Xylanolytic Activity Derived from Paenibacillus sp. K1J1,” African Journal of Microbiology Research, Vol. 4, No. 12, 2010, pp. 1257-1264.
- C. C. Lee, R. E. Kibblewhite-Accinelli, M. R. Smith, K. Wagschal, W. J. Orts and D. W. S. Wong, “Cloning of Bacillus licheniformis Xylanase Gene and Characterization of Recombinant Enzyme,” Current Microbiology, Vol. 57, No. 4, 2008, pp. 301-305.
- P. Sa-Pereira, M. Costa-Ferreira and M. R. Atres-Barros, “Enzymatic Properties of a Neutral Endo-1,3(4)-b-xylanase Xyl II from Bacillus subtili,” Journal of Biotechnology, Vol. 94, No. 3, 2002, pp. 256-275.
- K. Ratanakhanokchai, K. L. Kyu and M. Tanticharoen, “Purification and Properties of a Xylan-Binding Endoxylanase from Alkaliphilic Bacillus sp. Strain K-1,” Applied and Environmental Microbiology, Vol. 65, No. 2, 1999, pp. 694-697.
- P. K. A. Muniswaran, P. Selvakumar and N. C. L. Charyulu, “Production of Cellulases from Coconut Coir Pith in Solid State Fermentation,” Journal of Chemical Technology and Biotechnology, Vol. 60, No. 2, 1994, pp. 147- 151.
NOTES
*Corresponding author.